Методические особенности гистоморфометрии и стереологического анализа периферических нервов в экспериментальных исследованиях

Автор: Варсегова Т.Н.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Конкурсные работы молодых ученых
Статья в выпуске: 3, 2006 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/142120935
IDR: 142120935
The technique-related peculiarities of histomorphometry and stereologic analysis of peripheral nerves in experimental studies
Текст статьи Методические особенности гистоморфометрии и стереологического анализа периферических нервов в экспериментальных исследованиях
Федеральное государственное учреждение науки
«Российский научный центр "Восстановительная травматология и ортопедия" им. академика Г. А. Илизарова Росздрава», г. Курган (генеральный директор — заслуженный деятель науки РФ, член-корреспондент РАМН, д.м.н., профессор В.И. Шевцов)
Морфологическое описание берцовых нервов в условиях дистракционного остеосинтеза являлось предметом специальных исследований [1-4]. По мере разработки новых методик остеосинтеза возрастает интерес к изучению количественных характеристик периферических нервов, объективно отражающих полноту их мор- фологического восстановления после повреждений [9, 14, 16], а также направленность и временные параметры изменений их морфофункционального состояния в различных условиях эксперимента. Соответственно целесообразна и разработка адекватной технологии количественного исследования периферических нервов.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проведены на 23 взрослых беспородных собаках, которым после закрытой флексионной остеоклазии берцовых костей удлиняли голени аппаратом Илизарова с автоматическим приводом в течение 28 дней с суточным темпом 1 мм за 60 приемов (I серия) и в течение 10 дней с суточным темпом 3 мм за 180 приемов (II серия). Все эксперименты (содержание собак, хирургические вмешательства, эвтаназия) проведены в соответствии с нормативными документами (Приказ МЗ СССР № 755, 1977 г.). Животных выводили из опыта в конце дистракции (I серия n=7, II серия n=3), через 30 дней фиксации (I серия n=3, II серия n=3) и через месяц после снятия аппарата (I серия n=4, II серия n=3). Учитывая различную скорость деструкции тканей, забор и фиксацию большеберцовых нервов (ББН) осуществляли в первую очередь, так как аутолитические изменения в них наступают сразу после гибели животного. После эвтаназии ББН оперированных и контралатеральных конечностей на уровне средней трети голени закрепляли на твердой основе с целью избежания деформации и резецировали для морфологического исследования, отступая на 2 см дистальнее и проксимальнее от основы.1
Нервные стволы обладают ярко выраженной анизоморфностью, так как нервные пучки и волокна в них располагаются вдоль длинной оси. Таким образом, для получения репрезентативных выборок изображений с максимально возможным количеством анализируемых структур для
Научный руководитель работы д.м.н. М.М. Щудло.
морфометрического исследования использовали продольные либо поперечные патогистологиче-ские препараты (срезы). С целью визуализации интересующих структур и химических веществ в изучаемых объектах подобраны адекватные гистологические и гистохимические методики.
Для комплексного исследования периферических нервов на разных морфологических уровнях (органном, тканевом и клеточном) использовали срезы, толщина которых составляет примерно 1/20 диаметра изучаемых структур. При этом возникающий в результате затенения светлоокрашенных структур темноокрашенными эффект Холмса, вносящий систематическую ошибку при морфометрии, сводится к минимуму [11].
Проксимальные и дистальные фрагменты ББН отсекали, делили на столбики, размещали вертикально на заранее промаркированных фильтровальных бумажках и помещали в жидкий азот. На микротоме-криостате МК-25 ТУ 64-1-856-78 при t = -18 °С готовили тотальные поперечные срезы толщиной 7-10 мкм, монтировали на предметные стекла. С целью визуализации периневрия, кальций-кобальтовым методом [7] выявляли активность миозиновой АТФ-азы (рис. 1), являющейся маркером клеток с подвижной цитолеммой (в данном случае периневральных). Такие патогистологиче-ские препараты использовали для количественной оценки реакции нервных стволов на дозированное растяжение. С этой целью, используя большой исследовательский фотомикроскоп “Opton” (Германия) c аппаратно-программным комплексом “Di-aMorph” (Москва), оцифровывали изображения тотальных срезов, в них, используя программу “MEDIAS” (“DiaMorph”, Россия), измеряли общую площадь поперечного сечения нерва (An), суммарную площадь пучков нервных волокон вместе с окружающим их периневрием (Af-pn).
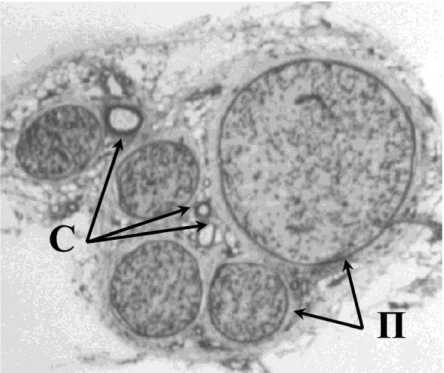
Рис. 1. Большеберцовый нерв собаки. Поперечный тотальный криостатный срез. Реакция на миозиновую АТФ-азу. Лупа 10 . П – периневрий; С – сосуды эпиневрия
Материал, закрепленный на твердой основе фиксировали в смеси 2 % растворов глутарового и параформальдегидов на фосфатном буфере (рН 7,4) и делили на столбики. Часть из них отмывали в проточной воде, на микротоме-криостате готовили продольные и поперечные срезы толщиной 25 мкм, монтировали на предметные стекла и им-прегнировали азотнокислым серебром по пунктам 6-10 методики А.К. Коломийцева с соавт 5 . Продольные срезы использовали для оценки изменения извитости нервных волокон, длин интерно-дальных сегментов, состояния насечек Шмидта-Лантермана и перехватов Ранвье.
Часть столбиков по специально разработанной автором методике заливали в аралдит. Возможности использования полутонких эпоксидных срезов (ПТС) ограничивала их малая стандартная площадь – до 1 мм2, затруднявшая получение репрезентативных выборок изображений для стереологических исследований. Чтобы преодолеть указанное противоречие, был разработан, апробирован и внедрен в повседневную практику ряд технических приемов, позволяющих увеличить площадь одномикронных полу-тонких срезов до 4-8 мм2: 1) применение специального режима проводки материала (увеличение сроков экспозиции и дополнительная пропитка в смоле с катализатором при комнатной температуре); 2) площадь основания пирамидки при ее высоте до 1,0 мм должна быть как минимум в 4 раза больше площади среза, что позволяет избежать вибрации блока и образования механических дефектов на срезах при ультрато-мировании; 3) тщательный контроль качества и отбор стеклянных ножей, либо использование алмазного ножа с достаточной длиной режущей кромки; 4) фиксация срезов к предметному стеклу, предварительно покрытому тонкой целлоидиновой пленкой, для предотвращения появления при окраске неровностей и складок из-за различных биомеханических свойств тканевых составляющих исследуемых объектов 12 .
Для количественного исследования внутри-ствольных составляющих использовали ПТС, окрашенные метиленовым синим и основным фуксином по Уикли 8 , позволяющие работать на больших увеличениях (объективы 40, 100 ). Оцифрованные изображения полутонких срезов (б) и электронограмм (а) периферических нервов сопоставимы (рис. 2), что дало возможность идентифицировать и исследовать на светооптическом уровне эндоневрий, безмиелиновые и миелиновые нервные волокна (МНВ) – диаметры 1-20 мкм.
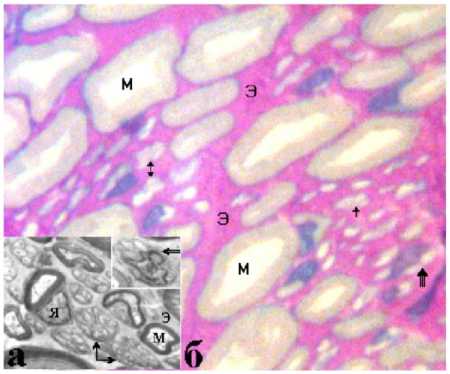
Рис. 2. Большеберцовый нерв собаки. Аралдит, поперечный ультратонкий (а, электронограмма, 1000 ) и полутонкий (б, окраска по Уикли, об. – 100; ок. – 12,5 ) срезы. Э – эндоневрий, М – мякотные, стрелки – без-мякотные нервные волокна, Я – ядро шванновской клетки в составе мякотного нервного волокна; двойная и тройная стрелки – безмякотные нервные волокна, срезанные на уровне ядра нейролеммоцита
С помощью тестовой решетки равноудаленных точек [13] в программе Adobe Photoshop 3,0 рассчитывали стереологические параметры: относительный объем нервных волокон и эндонев-рия, численные плотности МНВ (NA mnf ), их реактивно-деструктивно измененных форм и долю (%) последних в общем объёме выборки. В программе РhоtоFinish измеряли средние диаметры МНВ (Dmnf), строили прецизионные гистограммы их распределения с шагом 1 мкм. С целью исследования ББН как канала передачи информации определяли энтропию по Шеннону, относительную энтропию, избыточность и организацию [6].
Контролем послужили ББН 3 интактных беспородных собак. Достоверность различий определяли в программе «AtteStat» (версия 1,0; И.П. Гайдышев, 2003) по критериям рандомизации и Вилкоксона для независимых выборок.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты исследования показали, что в обеих сериях ББН на стороне удлинения макроскопически не обнаруживали признаков травма-тизации. Ни на одном из сроков эксперимента общие площади поперечных сечений ББН на стороне удлинения на уровне дистракционного регенерата достоверно не увеличивались относительно аналогичных контралатеральных нервов, а суммарная площадь пучков волокон, включая периневрий, достоверно повышалась только во II серии (табл. 1) через 10 дней дистракции в ускоренном тепе (3 мм в сутки).
По окончании дистракции в проводниковой части нервов удлиненных конечностей обнаружили единичные, преимущественно крупные проводники с признаками реактивно-деструктивных изменений, составившие в среднем 2,4 % в I и 2,6 % во II сериях (табл. 2).
Такие волокна встречаются и в интактных нервах, а незначительное повышение их количества, по мнению авторов, может быть связано с элиминацией нефункциональных в новых морфофизиологических условиях удлиненной конечности, преимущественно наиболее чувствительных миелиновых нервных волокон большого калибра, что согласуется с литературными данными [15].
Большинство же мякотных волокон (рис. 3, А) имело нормальную структуру. На последующих этапах эксперимента измененные волока также единичны (рис. 3, Б) в обеих сериях, и доля их сопоставима с контрольными значениями.
Таблица 1
Общая площадь поперечного среза нервного ствола (An) и суммарная площадь пучков нервных волокон с периневрием (Af-pn) на этапах эксперимента (М m)
|
Сроки (серия) эксперимента |
An (103мкм2) |
Af-pn (103мкм2) |
||
|
Опытная |
Контралатер. |
Опытная |
Контралатер. |
|
|
Д28(I) |
2311,6 279,5 |
1958,0 288,4 |
705,5 46,3 |
784,8 26,6 |
|
Ф30(I) |
3563,7 766,4 |
2650,3 344,8 |
868,2 135,7 |
903,3 90,3 |
|
БА30(I) |
2248,1 311,6 |
2029,1 221,0 |
820,8 64,9 |
778,7 58,2 |
|
Д10(II) |
2322,2 203,1 |
2234,7 385,1 |
917,0 64,7* |
718,5 23,9 |
|
Ф30(II) |
2194,4 396,6 |
2348,1 193,4 |
654,1 35,5 |
665,1 33,0 |
|
БА30(II) |
1765,3 588,7 |
1738,0 530,9 |
698,8 1,23 |
665,9 4,1 |
- различия между опытной и контралатеральной конечностями достоверны по критерию рандомизации при p < 0,05.
Таблица 2
Доли измененных мякотных нервных волокон в контроле и на этапах экспериментов
|
Сроки эксперимента |
Конец дистракции |
Фиксация 30 дней (Ф-30) |
Без аппарата 30 дней (БА-30) |
|
I серия |
(Д28) 2,37±0,74 % |
0,54±0,18 % |
0,85±0,13 % |
|
II серия |
(Д-10)2,61±0,11 % |
1,43±0,12 % |
1,61±0,12 % |
|
Контроль |
до 1,52 % |
||
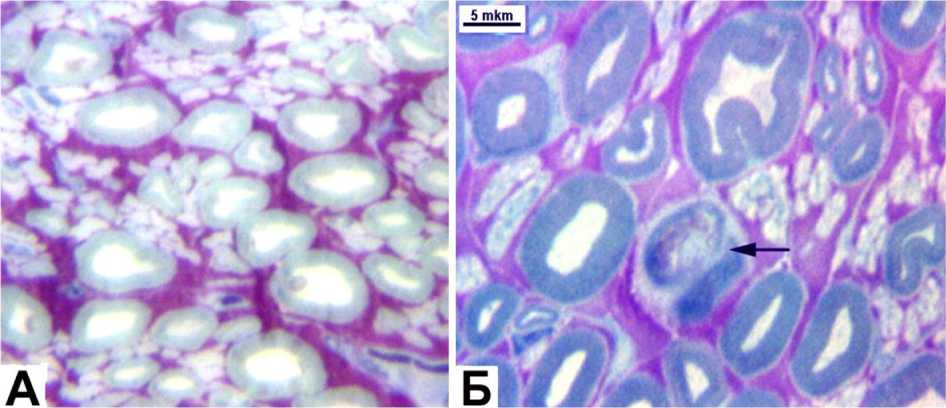
Рис. 3. Поперечные полутонкие срезы большеберцовых нервов, II серия, окраска по Уикли. А –– дистракция 10 дней. Об.-40, ок.- 12,5 ; Б – фиксация 30 дней. Об.- 100 МИ; ок.- 12,5 ; на стрелке – деструктивно измененное миелиновое волокно
Относительный объем волокон в I серии через 28 дней дистракции понижается в 1,4 раза по сравнению с контрольными значениями (рис. 4). Через 30 дней фиксации конечности в аппарате доля нервных волокон возрастает на 25,5 % по сравнению с предыдущим сроком, но остается достоверно пониженной (в 1,1 раза) относительно нормы. В конце эксперимента достоверные различия по этому параметру отсутствуют.
Во II серии в конце дистракции относительный объем эндоневрия оперированных конечностей повышается в 1,1 раза по сравнению с интактными значениями (рис. 4). Через 30 дней фиксации в аппарате этот параметр продолжает увеличиваться – соответственно объемная плотность нейральных элементов понижается в среднем на 10,1 % по сравнению с контролем. В конце эксперимента относительный объем нервных волокон повышается на 5,7 % относительно предыдущего срока наблюдается тенденция восстановления объемных отношений внутрипучко-вых элементов большеберцового нерва.
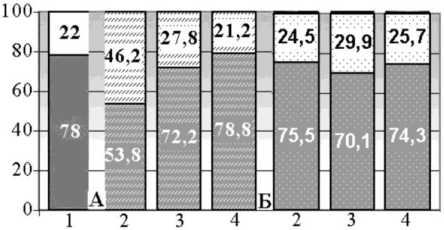
Рис. 4. Объемные плотности нервных волокон и эн-доневрия ББН в контроле ( 1 ) и на этапах удлинения в сериях I (А) и II (Б): 2 – конец дистракции, 3 – конец фиксации, 4 – без аппарата 30 дней
В I серии по окончании дистракции наряду с сохранившимися нервными проводниками в большеберцовом нерве удлиненной конечности обнаруживаются миелинизирующиеся аксоны с очень тонкой, чётко окрашенной миелиновой оболочкой. При этом численная плотность мякотных нервных волокон на 12 % превышает контрольные значения (рис. 5). На этапе фиксации NAmnf остается повышенной, но через 30 дней после снятия аппарата достоверные различия по этому параметру отсутствуют.
Во II серии к концу дистракции численная плотность миелиновых нервных волокон большеберцового нерва удлиненной конечности достоверно снижена на 20 % относительно контроля (рис. 5). Однако это нельзя рассматривать как свидетельство потери численности нервных проводников, поскольку в это время возрастают суммарная площадь пучков нервных волокон и объёмная доля эндоневрия в результате нарушения циркуляции эндоневральной жидкости. На этапе фиксации этот параметр достоверно превышает интактные значения в 1,1 раза, что, по мнению автора, связано с процессами регенерации и спраутинга: именно в это время в полутонких срезах обнаруживаются миелинизирующиеся аксоны малого диаметра. После снятия аппарата NAmnf нормализуется.
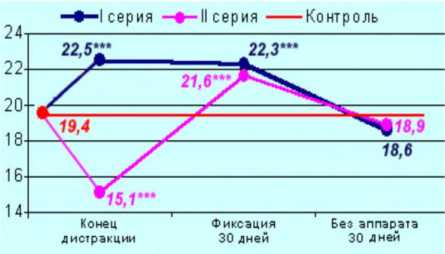
Рис. 5. Изменение численной плотности миелиновых нервных волокон ББН в контроле и на этапах удлинения голени собак автодистракторами в различных режимах: *** различия между опытом и контролем достоверны по критерию Вилкоксона при p<0,001
Такие колебания численной плотности МНВ отражают процессы новообразования волокон (в I серии преимущественно на этапе дистракции, во II серии – фиксации) и элиминации избыточных [10] и функционально «изношенных» нервных проводников на последующих этапах эксперимента.
Рассмотрим, как меняется распределение мякотных волокон по диаметру. В интактном большеберцовом нерве взрослых беспородных собак мякотные волокна имели диаметры от 2 до 17 мкм с бимодальным характером гистограммы распределения.
В конце дистракции в обеих экспериментах бимодальность сохранялась, моды находились в тех же диапазонах, что и в интактном материале (рис. 6). Однако в I серии, по сравнению с контролем, гистограмма смещена влево на 1 разряд: исчезли волокна с D>16,0 мкм и появилась фракция волокон диаметром менее 2,0 мкм. Средний диаметр волокон уменьшился и варьировал от 6,13 до 6,40 мкм (контроль 6,47-7,10). Во II серии в конце дистракции (рис. 6) гистограмма была укорочена справа на 3 разряда (волокна диаметром более 14,1 мкм отсутствовали), однако Dmnf варьировал от 7,07 до 8,39 мкм.
Через 30 дней фиксации в аппарате в I серии левая граница гистограммы восстанавливалась, D mnf возрастал до 6,95 мкм, но отсутствовали волокна диаметром более 15,-17,0 мкм (рис. 6). Через 30 дней после снятия аппарата популяция МНВ проявляет тенденцию к восстановлению: появляется фракция волокон диаметром 15,116,0 мкм. Dmnf превышал контрольные значения и варьировал от 7,21 до 7,30 мкм.
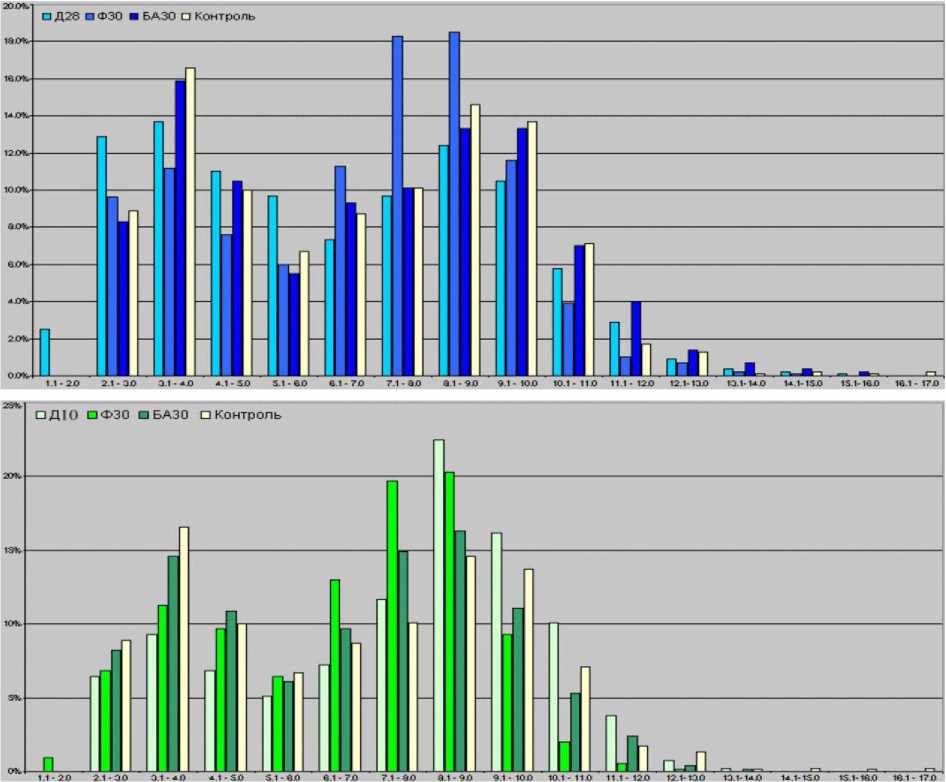
Рис. 6. Распределение по диаметрам миелиновых нервных волокон ББН интактных и опытных животных на этапах эксперимента: I серия (вверху) Д-28, Ф-30 и БА-30; II серия (внизу) Д-10, Ф-30 и БА-30
Во II серии через 30 дней фиксации гистограмма была укорочена справа уже на 4 разряда относительно контроля (рис. 6), так как отсутствовали волокна диаметром более 13,1 мкм. В то же время гистограмма смещалась влево: появлялась фракция волокон диаметром менее 2,0 мкм. При этом Dmnf уменьшался и варьировал от 6,23 до 6,91 мкм. Через 30 дней после снятия аппарата левая граница гистограммы восстанавливалась, правая проявляла тенденцию к восстановлению: вновь появлялась отсутствовавшая на предыдущем сроке фракция волокон диаметром 13,1-14,0 мкм. D mnf варьировал от 5,91 до 7,26 мкм.
Уменьшение D mnf было явлением временным: на этапах фиксации и после снятия аппарата происходила их структурная адаптация. Такое временное истончение нервных волокон, по мнению автора, связано с их упругой деформацией растяжения при удлинении голени. Последующее восстановление размерных параметров укладывается в известную [2, 4] концепцию об интеркалярном росте упруго напряженных тканевых структур за счет активизации биосинтетических процессов в клетках.
По данным информационного анализа, на всех сроках эксперимента в обеих сериях энтропия по Шеннону и относительная энтропия не отличались от контрольных значений, что свидетельствовало о сохранении упорядоченности большеберцового нерва как органа в условиях данных экспериментов. При этом в I серии в конце дистракции возрастали по сравнению с контролем избыточность и организация, снижаясь на этапе фиксации и возвращаясь к норме после снятия аппарата. Во II серии значения избыточности и организации были понижены на всех сроках эксперимента. Уменьшение избыточности свидетельствует о появлении помех и снижении надежности передачи информации по нерву, а ее возрастание – о том, что нерв как система становится более лабильным и менее экономичным [6].
Таким образом, результаты исследования большеберцовых нервов на разных этапах эксперимента показали, что при разных режимах удлинения реакция нервов на растяжение была однотипна. В I первой серии отклонение морфометрических и стереологических параметров от интактных значений наиболее выражено по окончании дистракции и к концу эксперимента достоверные различия отсутствуют: происходит структурно-функциональная адаптация нейронов к увеличению длины конечности. Во II серии удлинение производилось в течение короткого промежутка времени (10 дней), поэтому пластические перестройки большеберцового нерва наиболее выражены на этапе фиксации и не завершаются к концу эксперимента. Существенно то, что при разных режимах удлинения голени доля миелиновых нервных волокон с признаками реактивно-деструктивных изменений превышала интактные значения только в конце дистракции.
ЗАКЛЮЧЕНИЕ
Разработанная технология количественного исследования объективно отражает полноту восстановления периферических нервов, от ко- торой зависит и морфофункциональное состояние удлиняемой конечности в различных условиях дистракционного остеосинтеза.
