Методические особенности оценки формирования эктомикориз: изменчивость в связи с глубиной

Автор: Веселкин Д.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 3, 2013 года.
Бесплатный доступ
Рассмотрены особенности формирования боковых недетерминированных корней, поглощающих корней и эктомикориз в зависимости от глубины почвенных слоев у сеянцев Pinus sylvestris. Наибольшая активность формирования всех органов наблюдается в верхней части субстрата - в слое 40-120 мм от поверхности почвы. Предложено представление о функциональных зонах корневых систем, в которых по-разному реализуются функции роста и поглощения. Погрешности определения разных характеристик строения корней и микоризообразо-вания, которые могут быть связаны с неправильным отбором проб по глубине, могут быть 1.5-5-кратными и более.
Корневые системы, эктомикоризы, морфогенез, плотность корней, активность микори-зообразования
Короткий адрес: https://sciup.org/147204653
IDR: 147204653 | УДК: 581.557.24
Methodical features of evaluation of the ectomycorrhiza formation: variability in connection with depth
The article considers the formation peculiarities of lateral nondeterminate roots, absorbing roots and ectomycorrhizas depending on the depth of soil layers in Pinus sylvestris seedlings. The highest activity of all organs formation is observed at the top of a substrate in layer 40-120 mm from the soil surface. A representation of the functional areas of root systems in which different realized growth functions and acquisitions functions. The standard deviations in determining different characteristics of the root and mycorrhizas structure, which may be the result of incorrect selection sampling depth, may comprise 1.5-5 times or more.
Текст научной статьи Методические особенности оценки формирования эктомикориз: изменчивость в связи с глубиной
Важнейшее свойство почвы как среды существо вания организмов – анизотропность или неоднород ность условий как в горизонтальном, так и особенно в вертикальном направлениях [Карпачевский, 1981]. Вертикальная анизотропность почв выражается в за кономерном градиентном изменении их физических и химических характеристик по мере увеличения глу бины, т.е. по мере удаления от поверхности.
Специфичность почвенных условий предопре деляет соответствующую специфику распределе ния и состояния биоты. В частности, в отношении корней деревьев многократно показана неравно мерность их распределения по почвенному профи лю с максимум плотности (корненасыщенности) на некотором удалении от поверхности (в лесах бореальной и умеренной зон, как правило, 10–40 см) и более или менее постепенным снижением ко личества корней по мере дальнейшего углубления. Количество такого рода данных огромно [Орлов, Кошельков, 1971; Абражко, 1973; Бобкова, 1987; Никонов, 1987; Ярмишко, 1987, 1990; Веселкин, 2002а; Зайцев, 2008; и др.], что свидетельствует об универсальности подобного распределения, но с возможностью широкой модификации актуальны ми экологическими условиями. На фоне объектив ного обилия сведений о распределении в почве ин тегральных параметров корневых систем (биомас сы, корненасыщенности), существенно меньше ин формации о различиях тонких признаков строения корней на разной глубине или в разных гори зонтах, хотя и известно, что такие особенности мо гут быть выраженными, особенно при резких контрастах условий [Орлов, Кошельков, 1971; Се менова, 1980; Веселкин, 2003]. Эктомикоризные грибы – корневые симбионты деревьев, играющие ключевую роль в их почвенном питании, – также распределены в почве неравномерно [Шубин, 1998, 2000; Tedersoo et al., 2003; Genney, Anderson, Alexander, 2006]. В бореальных лесах выделены три частично разобщенные в вертикальном изме рении экологические ниши макромицетов-симбио трофов: первая – лесная подстилка; вторая – гуму сированные горизонты; третья – минеральная часть почвенного профиля [Шубин, 1998, 2000].
В настоящем сообщении на примере сеянцев сосны обыкновенной анализируются макроморфо логические особенности строения недетерминиро ванных корней, располагающихся на разной глуби не. Особое внимание уделяется при этом характери стикам развития микориз. Поставленная задача ак туальна с учетом двух моментов. Во-первых, подоб ный анализ, по нашему мнению, позволит лучше
понять механизмы морфогенеза и функционирова ния подземных органов, которые до настоящего времени изучены несравнимо меньше, чем механиз мы формирования надземных органов. Во-вторых, задача имеет выраженную методическую состав ляющую, так как необходимо ясно представлять степень сравнимости оценок признаков, получен ных для корней с разных глубин, из разных слоев или горизонтов.
Материал и методы
Для решения поставленной задачи проанализи ровано строение и микоризация корней 4-месяч ных особей сосны обыкновенной ( Pinus sylvestris L.), выращенных в разных вариантах вегетаци онного эксперимента [Веселкин, 2009а]. Сеянцы выращивались в пластиковых ящиках с перфори рованным дном, которые экспонировались в неота пливаемой теплице при естественной продолжи тельности светового дня. Субстрат – смесь несте рилизованной дерново-подзолистой среднесугли нистой почвы, песка и верхового торфа. Влаж ность почвы поддерживали на уровне 60% полной полевой влагоемкости. Эксперимент осуществлен как двухфакторный опыт с разной комбинацией уровней снабжения азотом и фосфором и взаимо действием факторов. Азот вносился в форме моче вины в дозах, эквивалентных внесению 0 кг/га (здесь и далее – действующего вещества) («N0»), 60 кг/га («N60»), 180 кг/га («N180»). Фосфор вно сился в тех же дозах («P0», «P60», «P180») в форме двойного суперфосфата.
После выкопки 4-месячных растений у них регистрировали признаки строения боковых кор ней 1-го порядка, отходящих от главных корней на разной глубине, т.е. на разном расстоянии от по верхности почвы. При измерениях главные корни сеянцев были разделены на отдельные фрагменты длиной 40 мм, начиная от уровня поверхности поч вы: 0–40, 40–80, …, 280–320, 320–360 мм. На каж дом таком 40-миллиметровом фрагменте регистри ровали количество отходящих боковых недетерми нированных корней 1-го порядка. Затем измеряли длину каждого бокового корня 1-го порядка, рас считывали плотность расположения коротких кор ней 2-го порядка на боковых корнях 1-го порядка и определяли активность микоризообразования, т.е. долю коротких корней 2-го порядка, заселенных эктомикоризными грибами.
Результаты и их обсуждение
Общие результаты эксперимента в части оцен ки влияния разных уровней биогенных элементов на особенности строения и микоризации корней частично опубликованы [Веселкин, 2009а]. Поэто му ниже основное внимание будет уделено особен ностям распределения признаков корневых систем по слоям глубины на примере двух эксперимен тальных вариантов («N60P0» и «N180P60»), а эф фекты биогенов будут обсуждаться в меньшей сте пени.
Формирование боковых недетерминированных корней
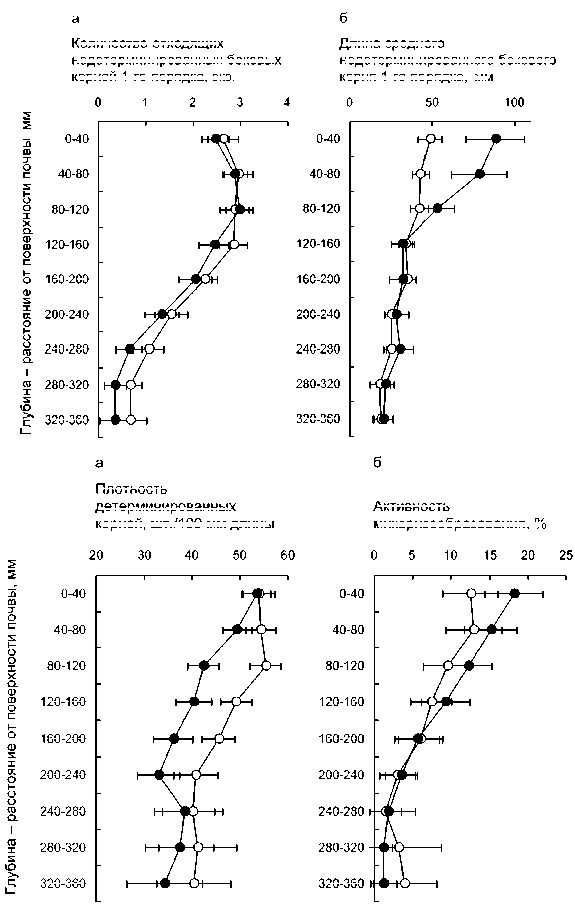
Активнее всего боковые недетерминированные (проводящие) корни 1-го порядка закладываются на глубине 40–160 мм от поверхности почвы (рис. 1а). В этом диапазоне глубин на каждых 40 мм главного корня возникает обычно 2.5–3.5 боковых недетер минированных корня 1-го порядка, в то время как на глубине более 200 мм на каждых 40 мм главных корней закладывается не более 0.5–1.0 бокового не детерминированного корня 1-го порядка. Эти раз личия, как и различия между двумя использованными для работы вариантами вегетационного опыта, стати стически значимы (результаты двухфакторного ANOVA: F глубина (8;954) = 88.94; P < 0.001; F вариант опыта (1;954) = 9.57; P = 0.002; F глубина х вариант (8;954) = 0.59; P = 0.791). В среднем несколько более активное за ложение боковых недетерминированных корней на блюдается при относительно меньшей обеспеченно сти элементами минерального питания – в варианте опыта N60P0, по сравнению с вариантом N180P60.
Длина среднего бокового недетерминированно го корня 1-го порядка напротив, несколько выше при обильном снабжении биогенами, в основном за счет мощного развития корней в верхних слоях суб страта – на глубине до 80 мм (рис. 1б). Но в обоих случаях хорошо просматривается последовательное убывание размера среднего бокового корня с глуби ной: от 40–50 мм в варианте N60P0 и 60–120 мм в варианте N180P60 до 10–30 мм на глубинах больше 200 мм в обоих вариантах. Все обсуждаемые раз личия длины среднего корня, т.е. и различия между вариантами опыта, и различия, связанные с глуби ной, статистически значимы ( F глубина (8;659) = 11.69; P < 0.001; F вариант опыта (1;659) = 6.31; P = 0.012; F глубина х вариант (8;659) = 4.33; P < 0.001).
Таким образом, в условиях вегетационного опы та у сеянцев Pinus sylvestris большая часть боковых проводящих корней закладывается в верхней части главного корня и здесь же, в верхних слоях суб страта, формируются в среднем наиболее длинные боковые проводящие корни.
Формирование поглощающих структур
Наряду с большим количеством и большими раз мерами, боковые корни, расположенные на глубинах до 80 мм, заметно лучше оснащены детерминирован ными поглощающими корнями, которые, к тому же, заметно активнее заселяются эктомикоризными гри бами, по сравнению с более глубоко расположенными слоями субстрата (рис. 2). Различия, связанные с глу биной, высоко значимы как в отношении плотности расположения детерминированных корней (F глуби- на (8;659) = 17.95; P < 0.001; F вариант опыта (1;659) = 24.98; P < 0.001; F глубина х вариант (8;659) = 2.27; P = 0.021), так и в
11.77; P < 0.001; F
глубина х вариант (8;659)
вариант опыта (1;659)
1.16; P = 0.320).
2.51
ина (8;659) = отношении активности формирования эктомикориз
120-160
160-200
160-200
200-240
200-240
240-280
240-28С
280-320
280-320
320-36С
320-360
Плотность
120-160
120-160
160-200
160-200
200-240
200-240
240-280
240-280
280-320
280-320
320-360
320-360
Количество отходящих недетерминированных боковых корней 1 -го порядка, экз.
Длина среднего недетерминированного бокового корня 1-го порядка, мм
Активность микоризообразования,% детерминированных корней, шт./100 мм длины
Рис. 1. Количество (а) и длина (б) недетерминированных боковых корней 1-го порядка в зависимости от расстояния до поверхности почвы у 4-месячных сеянцев Pinus sylvestris в вегетационном опыте при двух уровнях внесения биогенных элементов.
Варианты опыта здесь и на рисунках 2–3: ○ – N60P0, • – N180P60; горизонтальные линии – 95%-й доверительный интервал
Рис. 2. Плотность расположения детерминированных корней (а) и активность микоризообразования (б) в зависимости от расстояния до поверхности почвы у 4-месячных сеянцев Pinus sylvestris в вегетаци онном опыте при двух уровнях вне сения биогенных элементов. Все измерения относятся к недетерминированным боковым корням 1-го порядка
Общее количество корневых структур
Результатом интеграции рассмотренных част ных реакций на уровне особи является преимуще ственная локализация всех структур – недетермини рованных корней, коротких детерминированных корней, эктомикориз в верхних слоях субстрата (рис. 3). Изменения всех этих признаков с глубиной статистически значимы на высоких уровнях (во всех случаях P < 0,001). Следует отметить очень значительную контрастность абсолютных показа телей развития подземных органов между верхними и нижними слоями субстрата. Так, например, сум марная длина боковых проводящих корней в верх нем (0–80 мм) слое варьирует в интервале 250–450 мм, а в слое 200–360 мм – лишь в интервале 100–
150 мм. Суммарное количество коротких корней у особи в слое субстрата 0–80 мм варьирует в диапазоне 120–260 экз., а глубже 200 мм расположены лишь 30–85 коротких корней. Суммарное количество эктомикориз у особи в слое субстрата 0–80 мм составляет 15–40 экз., а в слое 200–360 мм расположены лишь единицы (1–5) эктомикоризных корней, а иногда они вообще отсутствуют.
Функциональные зоны корневых систем
Прежде чем переходить к обсуждению изложенных фактов, отметим, что, как большинство экспериментов, опыт с разным уровнем обеспеченности биогенными элементами лишь частично моделирует естественные условия.
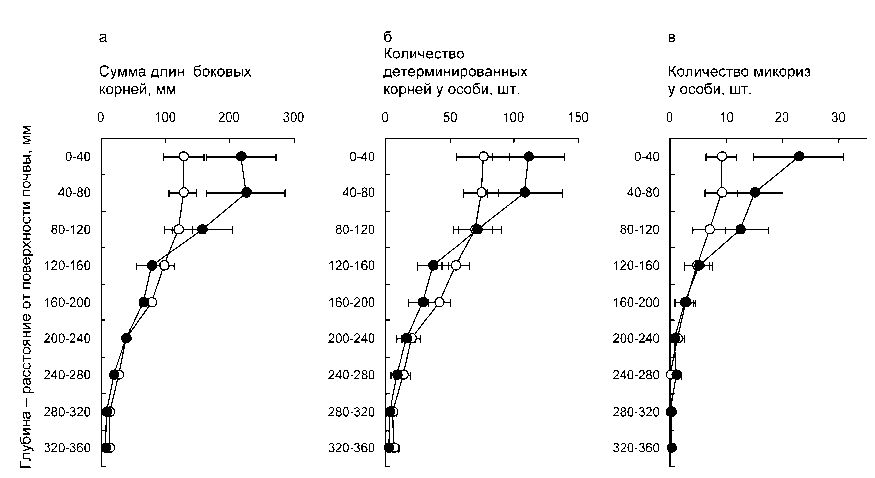
Рис. 3 . Суммарная длина боковых недетерминированных корней (а), абсолютное количество детерминированных корней (б) и микориз (в) в зависимости от расстояния до поверхности почвы у 4-месячных сеянцев Pinus sylvestris в вегетационном опыте при двух уровнях внесения биогенных элементов. Все измерения относятся только к недетерминированным боковым корням 1-го порядка.
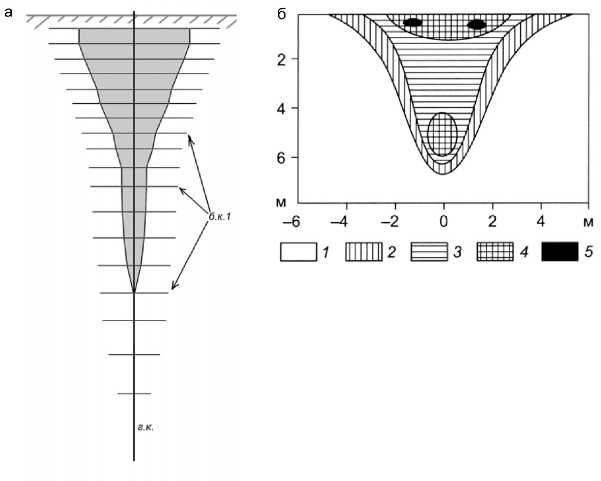
Рис. 4. Схематическое изображение функциональных зон корневой системы сеянца сосны (а) и 7-летней яблони (б). а: серая область – «зона поглощения»; незакрашенная область распространения боковых кор ней – «зона роста»; г.к. – главный корень; б.к.1 – недетерминированные боковые корни 1-го порядка; б: плотность поглощающих корней: 1 – <0,05; 2 – 0,05–0,5; 3 – 0,5–2,0; 4 – 2,0–5,0; 5 – 5,0–5,5 тыс. шт./м3 [по:
Ковалевский, Ковалевская, 2010]
В результате этого, например, размеры корне вых систем в эксперименте в 5–15 раз (!) больше, чем у всходов сравнимого возраста из ненарушен ных лесов [Veselkin, Sannikov, Sannikova, 2010] и в 2–5 раз больше, чем у однолетних сеянцев из лес ных питомников [Веселкин, 2006б; 2010]. Не ме нее контрастно различается и активность микори зообразования. В разных вариантах опыта она практически на порядок ниже, чем в естественных лесах, и в 2–5 раз ниже, чем в питомниках. Таким образом, при экстраполяции обсуждаемых далее закономерностей на более широкий круг условий необходима известная осторожность. Еще одно ограничение состоит в том, что наши данные лишь косвенно описывают изменчивость, связанную с глубиной, поскольку часть различий можно интер претировать как различия между участками корне вых систем разного возраста. Ясно, что участки корневых систем, расположенные глубоко или на периферии корневой системы, моложе, по сравне нию с участками, расположенными ближе к по верхности почвы или ближе к главному корню.
В пределах корневой системы можно выделить функционально различные зоны, структурно состо ящие из корней разных порядков (рис. 4а; соотно шение размеров корней разных порядков и корней, расположенных на разной глубине, близко к реаль ным). Поглощающие органы – короткие детерми нированные корни и эктомикоризы – в наи большем количестве сосредоточены в зоне, охва тывающей проксимальные части главного и бо ковых проводящих корней. Это связано, во-пер вых, со снятием апикального доминирования в силу удаленности растущих кончиков корней 0 и 1-го порядков, во-вторых, прямо с фактором воз раста корней, так как процесс формирования экто микориз требует времени. Зону, охватывающую проксимальные части главного и боковых прово дящих корней, можно обозначить как «зону погло щения» по наиболее характерной функции слагаю щих ее органов (на рис. 4а отмечена серым цве том). В зоне, включающей дистальные фрагменты главного и боковых корней, преобладающим про цессом является линейный рост корней с соответ ствующим проявлением апикального доминирова ния. Поэтому ее можно обозначить как «зону ро ста». Подобная дифференциация участков корне вых систем на основании квазифункциональных характеристик возможна с большей дробностью при условии более детальных исследований. Например, на основании количественной оценки плотности поглощающих корней возможна диффе ренциация объема, занимаемого корневой систе мой, на несколько классов (рис. 4б).
Пространственная разобщенность проявления функций роста и поглощения позволяет предпола гать их частичную независимость. Такое предположение может оказаться перспективным для решения некоторых вопросов экологии растений. Например, по нашим оценкам, в условиях загрязнения тяжелыми металлами критичным для выживания всходов является обеспечение именно роста недетерминированных корней. Размер корневых систем всходов в условиях загрязнения существенно снижается притом, что формирование поглощающих корней и их трансформация в эктомикоризы происходят с фоновой интенсивностью или даже активизируются [Веселкин, 2002б, 2006а].
Выраженность вертикальной неоднородности строения корневых систем свидетельствует об ограниченности описательных возможностей подхода, апеллирующего исключительно к различиям между корнями разного порядка [Усольцев, 2007; Веселкин, 2013]. Ясно, что признаки, важные для понимания и предсказания структуры корней, не связаны только с порядком их ветвления, как, впрочем, и не связаны исключительно с положением анализируемого фрагмента в градиенте между проксимальным и дистальным полюсами. Более правдоподобным является консенсусное описание способности корней к освоению объема и эксплуатации ресурсов почвы с объединением представлений о: (1) различии между корнями разных порядков и (2) существовании функциональных зон корневых систем (рис. 5).
Зоны преимущественной реализации функции роста I I поглощения дистальные фрагменты ? проксимальные фрагменты недетерминированных корней недетерминированных боковых
0-го (главный корень сеянца) и корней 1-го порядка с
1-го порядков с редко относительно часто расположенными расположенными детерминированными корнями и детерминированными корнями и слабым микоризообразованием активным микоризообразованием
Ослабление ростовой и усиление поглощающей активности
Рис. 5 . Участие разных корневых структур в формировании функциональных зон корневых систем. Вопросительный знак указывает на возможность выделения функциональных зон с промежуточными характеристиками
Последовательное разрастание корневой систе мы с постепенным отодвиганием зоны преимуще ственного размещения активных корней все дальше от основания ствола дерева – понятный и неоднократно описанный феномен [Колесников, 1962; Красильников, 1983], определяемый как ак ропетальный ростовой процесс [Усольцев, 2007]. Такой процесс последовательной смены функций, выражающийся в постепенном ослаблении одних функций и усилении других, носит для подземных органов универсальный характер вплоть до самых тонких корней. В частности, он ясно подтвержда ется при анализе абсолютных размеров и пропор ций между симбионтами в эктомикоризных корнях [Веселкин, 2009б].
Методические следствия
В методическом отношении важны следующие следствия из рассмотренных материалов.
Во-первых, на разной глубине относительные характеристики, описывающие развитие поглоща ющих органов, весьма существенно различаются. В большей степени это относится к такому пара метру? как «активность микоризообразования», ко торая между разными слоями почвы различается до 3–9 раз. Более консервативной, различающейся на разных глубинах не более чем в 1.5 раза, являет ся характеристика плотности детерминированных корней, что, вероятно, является следствием высо кой автоматизации закладки боковых корней [Бар лоу, Люк, 2008].
Для того, чтобы получать несмещенные оценки при сравнении разных особей или их групп по сте пени развития подземных органов, необходимо специально контролировать глубину отбора проб. При отсутствии такого контроля погрешности из мерений могут быть весьма существенными и до стигать нескольких раз. Многие методические сложности исследования корневых систем общеиз вестны. Например, непростой задачей является из влечение интактных корневых мочек из верхних слоев почвы, которые в естественных условиях за частую представляют плотную дернину. При рабо те в подобных условиях особенно необходимо стремиться стандартизировать глубину взятия об разцов для обеспечения сопоставимости результа тов.
Второе обстоятельство методического плана, заслуживающее упоминания, состоит в следую щем. Основные различия между эксперименталь ными вариантами выращивания растений, которые рассмотрены в настоящем сообщении, проявля лись лишь в верхнем 10-сантиметровом слое суб страта. В нижележащих слоях строение корневых систем между вариантами N60P0 и N180P60 прак тически не различалось. Этот пример демонстри рует, что с разной глубиной изменяются не просто особенности строения корней, но и характерная амплитуда их реакций на внешние факторы. Поэтому при проведении экологических или фи зиологических исследований учет глубины взятых образцов корней может помочь яснее представить степень проявления исследуемых эффектов.
Заключение
С увеличением глубины почвенного слоя в кор невых системах древесных растений, особенно, по видимому, у ювенильных растений, закономерно изменяются значения признаков, характеризующие ростовую и поглощающую активность. Причины этих различий обусловлены значительно автомати зированными морфогенетическими процессами. При их реализации в отдельных частях корневых систем последовательно сменяется выраженность признаков, свидетельствующих сначала о ростовой активности, а затем – о возрастающей специализа ции в направлении выполнения поглощающей функции.
Различия по способности формировать поглоща ющие корни и эктомикоризы между участками кор ней, расположенными на разной глубине, имеет большое методическое значение. Эти различия необходимо учитывать при планировании схем на блюдений в естественных и экспериментальных условиях для получения несмещенных оценок таких стандартных количественных характеристик мико ризообразования как «плотность микориз» или «ак тивность микоризообразования».
Список литературы Методические особенности оценки формирования эктомикориз: изменчивость в связи с глубиной
- Абражко М.А. Закономерности распределения и фракционный состав биомассы подземных частей//Структура и продуктивность еловых лесов южной тайги. Л., 1973. С. 109-117.
- Барлоу П.В., Люк Ж. Ритмический характер морфогенеза растений на примере повторяющегося образования клеток идиобластов//Физиология растений. 2008. Т. 55. № 2. С. 163-183.
- Бобкова К.С. Биологическая продуктивность хвойных лесов Европейского Северо-Востока. Л.: Наука, 1987. 156 с.
- Веселкин Д.В. Распределение тонких корней хвойных деревьев по почвенному профилю в условиях загрязнения выбросами медеплавильного производства//Экология. 2002а. № 4. С. 250-253.
- Веселкин Д.В. Строение и микоризация корней сеянцев ели и пихты при изменении почвенного субстрата//Лесоведение. 2002б. № 3. С. 12-17.
- Веселкин Д.В. Снижение длины поглощающих корней ели сибирской и пихты сибирской в условиях загрязнения тяжелыми металлами и S02//Лесоведение. 2003. № 3. C. 65-68.
- Веселкин Д.В. Морфология корневых систем и микоризообразование у ювенильных пихты сибирской и ели сибирской в условиях воздействия выбросов медеплавильного комбината//Лесоведение. 2006а. № 4. С. 52-60.
- Веселкин Д.В. Функциональное значение микоризообразования у однолетних сеянцев сосны и ели в лесных питомниках//Вестник Оренбургского государственного университета. 2006б. № 4(54). С. 12-18.
- Веселкин Д.В. Влияние уровня обеспеченности азотом и фосфором на структуру биомассы и развитие эктомикориз у всходов сосны обыкновенной//Аграрная Россия. 2009а. Спец. вып. С. 53-54.
- Веселкин Д.В. Возрастные изменения эктомикоризных корней Abies sibirica//Вестник Тверского государственного университета. Серия Биология и экология. 2009б. № 37. С. 119-126.
- Веселкин Д.В. Оценка влияния несимбиотических и симбиотических параметров подземных органов на развитие надземных органов всходов Pinus sylvestris//Экология. 2010. № 6. С. 414-419.
- Веселкин Д.В. Методические особенности оценки формирования эктомикориз: изменчивость в связи с порядком ветвления корней//Вестник Оренбургского государственного педагогического университета. Электрон. науч. журн. 2013. № 3 (7). С. 18-25.
- Зайцев Г.А. Адаптация корневых систем хвойных древесных растений к экстремальным лесорастительным условиям: автореф. дис.. д-ра биол. наук. Тольятти, 2008. 39 с.
- Карпачевский Л.О. Лес и лесные почвы. М.: Лесн. пром-сть, 1981. 264 с.
- Ковалевский А.Л., Ковалевская О.М. Биогеохимия урановых месторождений и методические основы их поиска. Новосибирск: Гео, 2010. 362 с.
- Колесников В.А. Корневая система плодовых и ягодных растений и методы ее изучения. М.: Сельхозгиз, 1962. 191 с.
- Красильников П.К. Методика полевого изучения подземных частей растений: c учетом специфики ресурсоведческих исследований. Л.: Наука, 1983. 208 с.
- Никонов В. В. Почвообразование на северном пределе сосновых биогеоценозов. Л.: Наука, 1987. 142 с.
- Орлов А.Я., Кошельков С.П. Почвенная экология сосны. М.: Наука, 1971. 322 с.
- Семенова Л.А. Особенности экологии микориз сосны обыкновенной в зависимости от эдафических условий и географической широты//Микоризные грибы и микоризы лесообразующих пород Севера. Петрозаводск, 1980. С. 133-147.
- Усольцев В.А. Биологическая продуктивность лесов Северной Евразии. Методы, база данных и ее приложения. Екатеринбург: УрО РАН, 2007. 635 c.
- Шубин В.И. Экологические ниши и сукцессии макромицетов-симбиотрофов в лесных экосистемах таежной зоны. I. Экологические ниши//Микология и фитопатология. 1998. Т. 32, вып. 6. С. 32-37.
- Шубин В.И. Сукцессии макромицетов-симбиотрофов в лесных экосистемах таежной зоны//Грибные сообщества лесных экосистем. М.; Петрозаводск, 2000. С. 181-206.
- Ярмишко В.Т. Корневая система как индикатор техногенного загрязнения//Ботанический журнал. 1987. Т. 72, № 3. С. 340-346.
- Ярмишко В.Т. Особенности развития корневых систем сосны//Влияние промышленного атмосферного загрязнения на сосновые леса Кольского полуострова. Л.: Наука, 1990. С. 84-94.
- Genney D.R., Anderson I.C., Alexander I.J. Fine-scale distribution of pine ectomycorrhizas and their extramatrical mycelium//New Phytologist. 2006. Vol. 170. P. 381-390.
- Tedersoo L. et al. Fine scale distribution of ectomycorrhizal fungi and roots across substrate layers including coarse woody debris in a mixed forest//New Phytologist. 2003. Vol. 159, № 1. P. 153-165.
- Veselkin D.V., Sannikov S.N., Sannikova N.S. Specific features of root system morphology and mycorrhiza formation in Scots pine seedlings from burned-out areas//Russian Journal of Ecology. 2010. Vol. 41, № 2. P. 139-146.
