Мезоэвтрофные ельники таволгово-дернистоосоковые и аконитово-таволговые сфагновые европейской России и Урала

Автор: Кучеров И.Б., Кутенков С.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.30, 2021 года.
Бесплатный доступ
Проведена доминантно-детерминантная классификация мезоэвтрофных ельников таволгово-дернистоосоковых и аконитово-таволговых сфагновых с доминированием Picea abies s.l., Filipendula ulmaria s.l., Sphagnum warnstorfii и (в зависимости от ассоциации) Carex cespitosa , либо Aconitum septentrionale в Европейской России и на Урале. На основе выборки из 123 геоботанических описаний выделено 5 субассоциаций с 2 вариантами ельников таволгово-дернистоосоковых и 2 варианта ельников аконитово-таволговых. Разнообразие синтаксонов обусловлено факторами широтной зональности и континентальности климата, особенностями внутриландшафтной приуроченности сообществ и спецификой почвообразующих пород. Ельники таволгово-вилюйскоосоковые развиты на сульфатных породах в северной тайге бассейнов Кулоя и Пинеги. Леса всех прочих синтаксонов чаще всего встречаются в средней тайге, произрастая как на силикатных, так и на карбонатных породах. Ельникам таволгово-дернистоосоковым типичной субассоциации и аконитово-таволговым свойственны долготные варианты. Для всех синтаксонов характерен высокий уровень видового богатства сосудистых растений и мхов.
Мезоэвтрофное заболачивание, ельники сфагновые, сфагновые мхи, классификация растительности, таежная зона, европейская Россия
Короткий адрес: https://sciup.org/148322408
IDR: 148322408 | УДК: 581.526.426.2: | DOI: 10.24412/2073-1035-2021-10387
Mesoeutrophic meadowsweet-tufted-sedge and wolfsbane-monkshood-meadowsweet peatmoss spruce forests of European Russia and Urals
Mesoeutrophic meadowsweet-tufted sedge and wolfsbane monkshood-meadowsweet peatmoss spruce forests, dominated by Picea abies s.l., Filipendula ulmaria s.l., Sphagnum warnstorfii and (in proper associations) Carex cespitosa or Aconitum septentrionale in the correspondent layers, have been classified in European Russia and the Urals using the dominant-determinant approach to vegetation. Based upon the set of 123 relevés, 5 subassociations with 2 variants are recognized in Piceetum (P.) filipenduloso-caricosum cespitosae together with 2 variants in P. sphagno warnstorfii-aconitosum. The observed syntaxonomic diversity is governed partly by latitudinal zonation and climate continentality changes, partly by community topographic location and bedrock differences. Stands of P. filipenduloso-caricosum cespitosae subass. caricosum juncellae grow on gypsum in the Kuloi R. and Pinega R. basins within the northern-boreal subzone. Forests of all the other recognized syntaxa occur mainly in the middle-boreal subzone on both carbonate and silicate bedrock. Longitudinal variants are distinguished in P. filipenduloso-caricosum typicum as well as in P. sphagno warnstorfii-aconitosum. A high level of both vascular and moss species diversity is typical for all the syntaxa.
Текст научной статьи Мезоэвтрофные ельники таволгово-дернистоосоковые и аконитово-таволговые сфагновые европейской России и Урала
ЦЕНОТИЧЕСКОЕ РАЗНООБРАЗИЕ МЕЗОЭВТРОФНЫХ ЕЛЬНИКОВ ТРАВЯНО-СФАГНОВЫХ И ПОРОЖДАЮЩИЕ ЕГО ПРИЧИНЫ
Мезоэвтрофные ельники (из Picea abies s.l.) травяно-сфагновые развиваются в условиях повышенного минерального богатства почвы при различной степени избыточности проточного увлажнения. Они произрастают по окраинам жестководных болот, в местах разгрузки минерализованных грунтовых вод, по долинам ручьев и в речных поймах. Отличительной чертой всех ельников данной группы является участие (вплоть до согосподства) Filipendula ulmaria s.l. и Sphagnum warnstorfii соответственно в травяном и моховом ярусах сообщества.
Площадь, занимаемая ельниками рассматриваемой группы, как правило, невелика. В северной тайге Карелии на все типы травяносфагновых и логовых ельников (совокупно сфагновых и травяных без сфагнового покрова) приходится не более 3% покрытой лесом площади, в средней тайге – не более 2%, при этом на еловые леса в целом – соответственно 23.5 и 47% [3]. В южной тайге центральной и юго-западной частей Ленинградской обл. еловые леса таволговокисличной, таволговой и болотно-травяной серий покрывают соответственно 4.9, 2.4 и 0.5% от общей площади еловых лесов [33]. Лишь в регионах с близким залеганием известняков площадь, занятая рассматриваемыми ельниками, возрастает [14, 36].
Несмотря на небольшие занимаемые ими площади, рассматриваемым ельникам почти в каждой местности свойствен высокий уровень ценотического разнообразия. Одной из причин тому является высокий уровень видового богатства сосудистых растений и мхов, обусловленный повышенным уровнем минерального богатства почвы и разнообразием микроэкотопов в условиях дифференцированного нанорельефа. Само по себе богатство видов порождает большее число их характерных сочетаний, значимых при классификации. Более важной причиной, однако, видится большое число факторов внешней среды, влияющих на пространственное распределение сообществ. Помимо влажности и богатства почвы, к их числу относится проточность, а в пойменных условиях также длительность затопления. В результате распределение сообществ по градиентам ведущих факторов среды оказывается многомерным, а дифференциацию выделенных синтаксонов с помощью де-терминантных групп трудно отобразить в форме единой таблицы, так как распределение слишком многих диагностических видов оказывается индивидуальным.
Ниже приводится схематический обзор ассоциаций мезоэвтрофных ельников травяно-сфагновых, выделенных доминантно-детерминантным методом (см. ниже) с перечислением видов их диагностических групп (Dt) по ярусам. Доминирующие виды отражены в названиях ассоциаций, а также выделены полужирным шрифтом.
Piceetum (P.) equisetoso-filipendulosum – ельник (е.) таволгово-хвощовый сфагновый. Долины рек, берега ручьев и окраины жестководных болот от южной полосы северной тайги до южной тайги. Dt: Gymnocarpium dryopteris, Equisetum sylvaticum , Carex globularis, Rubus saxatilis, Pla-giomnium ellipticum.
P. sphagno-geraniosum – е. гераневый сфагновый. Долины рек и берега ручьев в полосе крайнесеверной тайги, в том числе в горах. Dt: Juniperus communis s.l., Salix myrsinifolia s.l., S. phylicifolia, Avenella flexuosa s.l., Trollius europaeus, Rubus chamaemorus, Geranium sylvaticum s.l., Empetrum nigrum s.l., Solidago virgaurea s.l., Cirsium heterophyllum, Saussurea alpina.
P. filipenduloso-menyanthosum – е. таволгововахтовый сфагновый. Облесенные жестководные болота и выходы ключей. Dt: Equisetum fluviatile, Caltha palustris, Comarum palustre, Menyanthes trifoliata , Pseudobryum cinclidioides, Calliergon cordifolium.
P. filipenduloso-caricosum cespitosae – е. таволгово-дернистоосоковый сфагновый. Долины ручьев, скрытопроточные понижения и ложбины стока. Dt: Juniperus communis s.l., Rosa acicularis, Padus avium s.l., Equisetum palustre, E. pratense, Carex cespitosa , C. vaginata, Paris quadrifolia, Comarum palustre, Rubus saxatilis, Geum rivale, Geranium sylvaticum s.l. , Angelica sylvestris, Galium palustre, Plagiomnium ellipticum, Climacium dendroides, Calliergon cordifolium.
P. sphagno warnstorfii-aconitosum – е. аконитово-таволговый сфагновый. Окраины жестководных болот, долины ручьев и поймы рек в районах залегания известняков. К диагностическим видам предыдущей ассоциации добавляются Ribes nigrum, Equisetum scirpoides, Aconitum septentrionale , Ranunculus repens, Chrysosplenium alternifolium, Vicia sepium, Rhodobryum roseum, Brachythecium mildeanum, B. rivulare.
Константными видами всех без исключения ассоциаций выступают Picea abies s.l. и Betula pubescens s.l. в составе древостоя и подроста , Calamagrostis purpurea s.l. , Filipendula ulmaria s.l. , Vaccinium vitis-idaea, Orthilia secunda, Linnaea borealis, Sphagnum warnstorfii , Hylocomium splendens. В отдельных ассоциациях к этому списку видов могут добавляться дополнительные константы.
В настоящей статье рассмотрены последние две из упомянутых ассоциаций – ельники таволгово-дернистоосоковые и аконитово-таволговые, – самые флористически богатые из всех синтак-сонов мезоэвтрофных ельников травяносфагновых.
ИСПОЛЬЗОВАННЫЕ ДАННЫЕ И МЕТОДЫ
При классификации ельников таволговодернистоосоковых и аконитово-таволговых использована выборка из 123 геоботанических описаний, из которых 112 выполнены авторами в экспедициях 1996–2019 гг. При этом оценивались проективные покрытия (ПП) видов по ярусам на площади не менее 400 м2, выполнялась почвенная прикопка или измерялась мощность торфа. Опубликовано подробное описание методики [11]. Шесть описаний взяты из литературных источников [1, 6, 7, 25, 29], еще 4 – из рукописи докторской диссертации Ю.П. Юдина [37] в библиотеке БИН РАН, 1 – из отчета Я.Л. Пааля [18] в заповеднике «Кивач».
Как и при выделении синтаксонов ельников черничных и хвощовых сфагновых [12, 13], проведена доминантно-детерминантная классификация. Синтаксоны выделялись по доминантам, затем их объем уточнялся с помощью детерми-нантных групп экологически близких видов. К одной ассоциации отнесены описания с одинаковым набором доминантов и детерминантов на всем протяжении ее ареала. Для субассоциаций могут быть характерны собственные доминанты и детерминанты вдобавок к таковым ассоциации в целом. Географические варианты могут выделяться только по детерминантам. Выделенные 8 синтаксонов разного ранга сведены в фитоцено-тическую таблицу (табл. 1), где содержатся также сведения об общем покрытии и высоте ярусов. Названия синтаксонов даны следуя традиции школы В.Н. Сукачева [32]; в качестве синонимов приводятся названия в форме, использованной в предыдущих работах по доминантно-детерминантной классификации [11].
Номенклатура сосудистых растений дана по С.К. Черепанову [35], листостебельных мхов – по М.С. Игнатову с соавторами [41], печеночных мхов – по А.Д. Потемкину и Е.В. Софроновой [21], лишайников – по «Списку лихенофлоры России» [31]. Европейские таежные популяции ели рассматриваются как единый комплекс Picea abies s.l. [20]), березы пушистой – как Betula pu-bescens s.l. (incl. B. subarctica, B. x aurata [34]). При наименовании синтаксонов использован принцип nomina conservanda, позволяющий ис- пользовать традиционно употребляемые синонимы таксонов. Границы зон и подзон растительности приняты по Т.И. Исаченко и Е.М. Лав-ренко [5] с уточнениями [26]. Зональная обусловленность ареалов синтаксонов оценена с помощью сумм превышений среднесуточных температур над базовой температурой в 10°C («гра-дусо-дней выше 10°C», degree-days above 10°C) по данным глобальной спутниковой метеосъемки [43]. На основе этой же базы вычислены индексы континентальности Конрада [47] (табл. 2). Точечные ареалы синтаксонов (рис. 1, 2) учитывают как совокупность анализируемых описаний, так и дополнительные данные литературы.
ОБЩИЕ ЧЕРТЫ ХАРАКТЕРИЗУЕМЫХ АССОЦИАЦИЙ
Для ельников аконитово-таволговых и для большинства синтаксонов таволгово-дернистоосоковых характерен лишь частично сомкнутый моховой ярус, в составе которого Sphagnum warnstorfii сочетается с таежными и болотными зелеными мхами (табл. 1). Это придает данным синтаксонам пограничный характер между травяно-сфагновой и травяной (без сфагнового покрова) группами ассоциаций еловых лесов, о чем писали и ранее [1. 14]. Многие авторы [9, 17, 22, 23] относят осоково-сфагновые ельники с господством Carex cespitosa к болотно-травяной группе ассоциаций вместе с аконитовотаволговыми, а также таволговыми без сфагнового ковра и торфяной залежи, развитыми на суглинистых отложениях в поймах.
О сходстве ельников таволгово-дернистоосоковых и аконитово-таволговых сфагновых с таволговыми без сфагнового покрова говорит и сопоставимо высокий уровень видового богатства сообществ, существенно превосходящий таковой в описаниях ельников других ассоциаций и групп ассоциаций. На одно описание ельников таволгово-дернистоосоковых, сделанное авторами статьи, приходится 50–110 видов сосудистых растений и мхов, взятых вместе. Такие же цифры характерны для ельников аконитовотаволговых сфагновых, а также собственно таволговых и аконитово-таволговых. Более высокий уровень видового богатства в пределах изучаемой территории отмечен лишь в некоторых ассоциациях широколиственных лесов и в южно- и подтаежных черноольшанниках. В то же время для мезотрофных ельников черничных сфагновых характерно порядка 25–40 видов сосудистых растений и мхов на описание, для черничных зеленомошных – 25–50, для хвощовых сфагновых – 35–50.
Фитоценотическая характеристика мезоэвтрофных ельников таволгово-дернистоосоковых и аконитово-таволговых сфагновых Европейской России и Урала
|
Вид |
Ярус |
Синтаксоны |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
||
Детерминанты субассоциаций и вариантов
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
|
Avenella flexuosa s.l. |
c |
561 |
12+ |
5+ |
4+ |
||||
|
Dicranum majus |
z+d |
441 |
17+ |
33+ |
23+ |
8+ |
33+ |
23+ |
25+ |
|
D. polysetum |
d |
561 |
33+ |
11+ |
8+ |
18+ |
4+ |
||
|
Sphagnum girgensohnii |
d |
3312 |
177 |
111 |
193 |
214 |
5+ |
||
|
Polytrichum commune |
d+z |
331 |
332 |
4+ |
131 |
5+ |
4+ |
||
|
Pinus sylvestris |
a1+2 |
444 |
504 |
672 |
582 |
211 |
1003 |
231 |
|
|
Sphagnum centrale |
d |
44+ |
50 9 |
445 |
354 |
293 |
18+ |
4+ |
|
|
Daphne mezereum |
b |
44+ |
33+ |
44+ |
31+ |
13+ |
50+ |
42+ |
|
|
Vaccinium myrtillus |
c |
10019 |
836 |
892 |
731 |
501 |
864 |
671 |
|
|
Gymnocarpium dryopteris |
c |
672 |
671 |
33+ |
811 |
712 |
50+ |
962 |
|
|
Oxalis acetosella |
c |
893 |
831 |
56+ |
883 |
712 |
955 |
1004 |
|
|
Luzula pilosa |
c |
89+ |
50+ |
44+ |
77+ |
54+ |
82+ |
71+ |
|
|
Dryopteris carthusiana |
c |
781 |
833 |
44+ |
62+ |
29+ |
641 |
71+ |
|
|
Crepis paludosa |
c |
67+ |
331 |
671 |
811 |
581 |
772 |
963 |
|
|
Cirsium heterophyllum |
c |
561 |
67+ |
33+ |
35+ |
461 |
592 |
542 |
|
|
Solidago virgaurea s.l. |
c |
56+ |
33+ |
78+ |
42+ |
29+ |
55+ |
29+ |
|
|
Lycopodium annotinum |
c |
673 |
33+ |
56+ |
15+ |
25+ |
41+ |
33+ |
|
|
Athyrium filix-femina |
c |
44+ |
507 |
33+ |
421 |
29+ |
551 |
38+ |
|
|
Pleurozium schreberi |
d |
10012 |
1009 |
1005 |
964 |
713 |
775 |
1005 |
|
|
Rhizomnium pseudopunctatum |
d+z |
44+ |
33+ |
441 |
621 |
501 |
553 |
922 |
|
|
Dicranum scoparium |
d+z |
44+ |
33+ |
56+ |
65+ |
58+ |
45+ |
67+ |
|
|
Ptilium crista-castrensis |
d+z |
56+ |
33+ |
27+ |
332 |
41+ |
752 |
||
|
Dicranum fuscescens |
z+d |
44+ |
50+ |
11+ |
38+ |
29+ |
14+ |
38+ |
|
|
Salix pentandra |
a 2 +b |
17+ |
22+ |
191 |
8+ |
332 |
5+ |
13+ |
|
|
Caltha palustris |
c |
11+ |
33+ |
22+ |
54+ |
541 |
33+ |
41+ |
631 |
|
Epilobium palustre |
c |
22+ |
50+ |
35+ |
33+ |
33+ |
18+ |
54+ |
|
|
Pseudobryum cinclidioides |
d |
832 |
561 |
693 |
636 |
675 |
271 |
884 |
|
|
Carex loliacea |
c |
11+ |
10012 |
33+ |
46+ |
33+ |
36+ |
50+ |
|
|
C. disperma |
c |
501 |
44+ |
621 |
501 |
45+ |
75+ |
||
|
Deschampsia cespitosa |
c |
33+ |
33+ |
44+ |
311 |
13+ |
18+ |
21+ |
|
|
Carex paupercula |
c |
11+ |
17+ |
56+ |
35+ |
4+ |
4+ |
||
|
Dactylorhiza maculata |
c |
17+ |
56+ |
31+ |
13+ |
18+ |
4+ |
||
|
Phragmites australis |
c |
171 |
563 |
232 |
8+ |
91 |
4+ |
||
|
Molinia caerulea |
c |
10010 |
15+ |
||||||
|
Potentilla erecta |
c |
56+ |
12+ |
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
|
Carex flava |
c |
44+ |
4+ |
||||||
|
Frangula alnus |
b |
331 |
171 |
892 |
691 |
13+ |
23+ |
8+ |
|
|
Convallaria majalis |
c |
33+ |
331 |
892 |
54+ |
4+ |
14+ |
4+ |
|
|
Equisetum fluviatile |
c |
33+ |
33+ |
44+ |
541 |
29+ |
1001 |
271 |
17+ |
|
Carex canescens |
c |
11+ |
17+ |
44+ |
31+ |
421 |
1002 |
9+ |
25+ |
|
Salix myrsinifolia |
a 2 +b |
11+ |
17+ |
44+ |
191 |
81 |
10010 |
9+ |
13+ |
|
Betula pubescens s.l. |
b |
33+ |
171 |
44+ |
731 |
831 |
67+ |
451 |
71+ |
|
Rubus chamaemorus |
c |
11+ |
33+ |
44+ |
19+ |
631 |
67+ |
32+ |
46+ |
|
R. arcticus |
c |
111 |
33+ |
22+ |
54+ |
461 |
1003 |
23+ |
42+ |
|
Pyrola minor |
c |
22+ |
17+ |
44+ |
27+ |
38+ |
33+ |
41+ |
46+ |
|
Moneses uniflora |
c |
11+ |
17+ |
33+ |
27+ |
8+ |
33+ |
41+ |
8+ |
|
Pyrola rotundifolia |
c |
33+ |
44+ |
42+ |
21+ |
1004 |
32+ |
38+ |
|
|
Calliergon giganteum |
d |
22+ |
81 |
251 |
678 |
9+ |
251 |
||
|
Helodium blandowii |
d |
22+ |
19+ |
4+ |
67+ |
5+ |
17+ |
||
|
Calliergonella cuspidata |
d |
22+ |
501 |
332 |
231 |
291 |
|||
|
Galium boreale |
c |
11+ |
44+ |
15+ |
33+ |
67+ |
18+ |
42+ |
|
|
Salix caprea |
a+b |
11+ |
33+ |
8+ |
291 |
672 |
23+ |
38+ |
|
|
S. phylicifolia |
b |
17+ |
11+ |
21+ |
1008 |
9+ |
33+ |
||
|
Bistorta major |
c |
22+ |
33+ |
12+ |
50+ |
67+ |
501 |
711 |
|
|
Lonicera pallasii s.l. |
b |
11+ |
11+ |
27+ |
46+ |
1001 |
23+ |
75+ |
|
|
Veratrum lobelianum |
c |
11+ |
381 |
67+ |
21+ |
||||
|
Atragene sibirica |
c |
21+ |
67+ |
9+ |
33+ |
||||
|
Lathyrus vernus |
c |
11+ |
17+ |
33+ |
31+ |
54+ |
912 |
791 |
|
|
Cirsium oleraceum |
c |
22+ |
11+ |
27+ |
422 |
732 |
794 |
||
|
Ranunculus subborealis |
c |
33+ |
17+ |
631 |
64+ |
88+ |
|||
|
Trollius europaeus |
c |
11+ |
42+ |
33+ |
451 |
631 |
|||
|
Sanionia uncinata |
z+d |
11+ |
17+ |
35+ |
42+ |
331 |
27+ |
711 |
|
|
Salix jenisseensis |
a 2 +b |
4+ |
672 |
||||||
|
S. lapponum |
b |
4+ |
1005 |
||||||
|
S. hastata |
b |
1002 |
|||||||
|
Saussurea alpina |
c |
11+ |
17+ |
4+ |
17+ |
1001 |
5+ |
29+ |
|
|
Naumburgia thyrsiflora |
c |
11+ |
12+ |
8+ |
1002 |
5+ |
13+ |
||
|
Parnassia palustris |
c |
22+ |
15+ |
8+ |
100+ |
||||
|
Carex juncella |
c |
4+ |
10028 |
5+ |
4+ |
||||
|
Veronica longifolia |
c |
8+ |
100+ |
17+ |
|||||
|
Orthilia obtusata |
c |
100+ |
|||||||
|
Cardamine pratensis s.l. |
c |
4+ |
13+ |
67+ |
5+ |
4+ |
|||
|
Ligularia sibirica |
c |
8+ |
671 |
14+ |
4+ |
||||
|
Lathyrus palustris |
c |
8+ |
67+ |
4+ |
|||||
|
Thalictrum kemense |
c |
4+ |
67+ |
||||||
|
Cicuta virosa |
c |
671 |
|||||||
|
Pedicularis sceptrum-carolinum |
c |
67+ |
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
|
Plagiomnium rostratum |
d |
172 |
4+ |
67+ |
182 |
||||
|
Peltigera aphthosa s.l. |
d+z |
11+ |
8+ |
67+ |
4+ |
||||
|
Bryum pseudotriquetrum |
d |
17+ |
22+ |
27+ |
13+ |
67+ |
27+ |
42+ |
|
|
Rhytidiadelphus subpinnatus |
d+z |
11+ |
17+ |
8+ |
17+ |
33+ |
141 |
382 |
|
|
Ribes nigrum |
b |
22+ |
17+ |
11+ |
151 |
33+ |
33+ |
451 |
672 |
|
Aconitum septentrionale |
c |
11+ |
331 |
867 |
1004 |
||||
|
Vicia sepium |
c |
11+ |
33+ |
11+ |
23+ |
33+ |
68+ |
58+ |
|
|
Ranunculus repens |
c |
11+ |
8+ |
29+ |
33+ |
411 |
42+ |
||
|
Equisetum scirpoides |
c |
17+ |
12+ |
25+ |
36+ |
33+ |
|||
|
Chrysosplenium alternifolium |
c |
8+ |
81 |
14+ |
38+ |
||||
|
Brachythecium rivulare |
d |
11+ |
17+ |
22+ |
31+ |
211 |
452 |
63+ |
|
|
Rhodobryum roseum |
d+z |
33+ |
11+ |
35+ |
8+ |
32+ |
42+ |
||
|
Brachythecium mildeanum |
d+z |
12+ |
17+ |
361 |
291 |
||||
|
Rubus humulifolius |
c |
221 |
173 |
231 |
211 |
23+ |
541 |
||
|
Cardamine amara |
c |
11+ |
151 |
21+ |
32+ |
58+ |
|||
|
Galium uliginosum |
c |
17+ |
33+ |
35+ |
331 |
18+ |
50+ |
||
|
Warnstorfia exannulata |
d |
17+ |
4+ |
29+ |
332 |
50+ |
|||
|
Calliergon richardsonii |
d |
11+ |
12+ |
291 |
672 |
||||
|
Bryum weigelii |
d |
11+ |
12+ |
4+ |
18+ |
42+ |
|||
|
Pohlia nutans s.l. |
z+d |
11+ |
33+ |
22+ |
19+ |
25+ |
14+ |
58+ |
|
|
Sciuro-hypnum curtum s.l. |
z+d |
11+ |
33+ |
231 |
33+ |
5+ |
50+ |
||
|
Brachythecium salebrosum |
z+d |
22+ |
17+ |
22+ |
35+ |
17+ |
9+ |
63+ |
|
|
Sciuro-hypnum reflexum |
z+d |
11+ |
33+ |
19+ |
21+ |
50+ |
|||
|
Детерминанты обеих ассоциаций |
|||||||||
|
Padus avium s.l. |
a2+b |
331 |
50+ |
33+ |
42+ |
25+ |
33+ |
45+ |
21+ |
|
Juniperus communis s.l. |
b |
44+ |
33+ |
896 |
771 |
671 |
1008 |
501 |
711 |
|
Rosa acicularis |
b |
781 |
67+ |
781 |
73+ |
751 |
1003 |
73+ |
922 |
|
Rubus saxatilis |
c |
1004 |
501 |
671 |
963 |
541 |
672 |
1003 |
672 |
|
Geranium sylvaticum s.l. |
c |
671 |
33+ |
781 |
46+ |
832 |
1001 |
773 |
1002 |
|
Equisetum palustre |
c |
561 |
671 |
561 |
923 |
793 |
1002 |
681 |
831 |
|
Geum rivale |
c |
441 |
50+ |
782 |
652 |
712 |
33+ |
862 |
963 |
|
Carex cespitosa |
c |
561 |
503 |
672 |
10020 |
10010 |
1003 |
731 |
71+ |
|
C. vaginata |
c |
33+ |
501 |
891 |
692 |
29+ |
1002 |
50+ |
67+ |
|
Angelica sylvestris |
c |
331 |
33+ |
78+ |
50+ |
58+ |
33+ |
64+ |
71+ |
|
Equisetum pratense |
c |
565 |
50+ |
331 |
27+ |
421 |
331 |
452 |
671 |
|
Paris quadrifolia |
c |
22+ |
33+ |
67+ |
65+ |
58+ |
33+ |
77+ |
96+ |
|
Comarum palustre |
c |
22+ |
50+ |
33+ |
651 |
633 |
1005 |
18+ |
58+ |
|
Galium palustre |
c |
22+ |
331 |
44+ |
62+ |
29+ |
67+ |
45+ |
42+ |
|
Plagiomnium ellipticum |
d+z |
332 |
33+ |
672 |
773 |
463 |
332 |
685 |
967 |
|
Calliergon cordifolium |
d+z |
22+ |
673 |
22+ |
732 |
501 |
676 |
412 |
672 |
|
Climacium dendroides |
d+z |
44+ |
33+ |
33+ |
621 |
422 |
1002 |
411 |
501 |
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
|
Константные виды |
|||||||||
|
Picea abies s.l. |
a1 |
10050 |
10039 |
10029 |
10030 |
10033 |
10027 |
10039 |
10038 |
|
Betula pubescens s.l. |
a 1 |
898 |
8318 |
10016 |
9615 |
9610 |
10011 |
9111 |
8813 |
|
Picea abies s.l. |
a2 |
6710 |
173 |
221 |
427 |
292 |
10015 |
555 |
293 |
|
Betula pubescens s.l. |
a 2 |
222 |
221 |
231 |
13+ |
10022 |
271 |
211 |
|
|
Sorbus aucuparia s.l. |
a 2 +b |
1004 |
832 |
89+ |
811 |
461 |
100+ |
912 |
712 |
|
Alnus incana |
a2+b |
442 |
502 |
44+ |
812 |
461 |
10022 |
452 |
581 |
|
Picea abies s.l. |
b |
8912 |
675 |
673 |
966 |
964 |
1003 |
915 |
1006 |
|
Vaccinium vitis-idaea |
c |
1005 |
10014 |
1003 |
1004 |
882 |
1004 |
952 |
881 |
|
Trientalis europaea |
c |
892 |
67+ |
89+ |
1001 |
751 |
1002 |
1001 |
1001 |
|
Linnaea borealis |
c |
893 |
1001 |
56+ |
811 |
631 |
33+ |
861 |
1001 |
|
Filipendula ulmaria s.l. |
c |
673 |
831 |
899 |
9612 |
9618 |
1007 |
9520 |
9615 |
|
Maianthemum bifolium |
c |
891 |
832 |
100+ |
881 |
751 |
671 |
952 |
921 |
|
Orthilia secunda |
c |
1001 |
50+ |
67+ |
85+ |
63+ |
33+ |
77+ |
71+ |
|
Equisetum sylvaticum |
c |
782 |
834 |
33+ |
421 |
543 |
33+ |
825 |
752 |
|
Calamagrostis canescens + purpurea s.l. |
c |
552 |
833 |
1001 |
962 |
923 |
1005 |
73+ |
922 |
|
Viola epipsila |
c |
562 |
832 |
891 |
923 |
793 |
1004 |
861 |
882 |
|
Chamaenerion angustifolium |
c |
44+ |
17+ |
56+ |
35+ |
631 |
67+ |
41+ |
67+ |
|
Melica nutans |
c |
56+ |
33+ |
54+ |
25+ |
33+ |
68+ |
50+ |
|
|
Hylocomium splendens |
d |
10017 |
1006 |
894 |
925 |
754 |
10013 |
866 |
1008 |
|
Sphagnum warnstorfii |
d |
442 |
5012 |
10032 |
9222 |
7516 |
10045 |
9525 |
10015 |
|
Aulacomnium palustre |
d |
331 |
50+ |
67+ |
58+ |
631 |
33+ |
55+ |
67+ |
|
Rhytidiadelphus triquetrus |
d+z |
447 |
831 |
441 |
733 |
675 |
1005 |
7312 |
969 |
|
Прочие виды |
|||||||||
|
Pinus sylvestris |
b |
11+ |
17+ |
22+ |
23+ |
5+ |
4+ |
||
|
Ribes spicatum |
b |
11+ |
17+ |
12+ |
29+ |
27+ |
33+ |
||
|
Rubus idaeus |
b |
11+ |
17+ |
8+ |
17+ |
18+ |
21+ |
||
|
Viburnum opulus |
b |
22+ |
11+ |
15+ |
4+ |
9+ |
4+ |
||
|
Rosa majalis |
b |
12+ |
4+ |
33+ |
8+ |
||||
|
Abies sibirica |
b |
111 |
21+ |
171 |
|||||
|
Betula pendula |
b |
331 |
|||||||
|
Carex globularis |
c |
332 |
331 |
33+ |
23+ |
211 |
32+ |
33+ |
|
|
C. rhynchophysa |
c |
11+ |
171 |
11+ |
17+ |
332 |
9+ |
21+ |
|
|
Melampyrum sylvaticum s.l. |
c |
22+ |
17+ |
33+ |
15+ |
32+ |
13+ |
||
|
Milium effusum |
c |
11+ |
17+ |
15+ |
29+ |
18+ |
25+ |
||
|
Listera cordata |
c |
22+ |
17+ |
4+ |
4+ |
9+ |
17+ |
||
|
Phegopteris connectilis |
c |
22+ |
11+ |
23+ |
13+ |
36+ |
21+ |
||
|
Melampyrum pratense |
c |
22+ |
11+ |
12+ |
17+ |
9+ |
13+ |
||
|
Listera ovata |
c |
11+ |
11+ |
12+ |
8+ |
36+ |
17+ |
||
|
Goodyera repens |
c |
33+ |
17+ |
11+ |
4+ |
9+ |
|||
|
Dactylorhiza hebridensis |
c |
33+ |
11+ |
19+ |
8+ |
18+ |
|||
|
Dryopteris expansa s.l. |
c |
11+ |
4+ |
21+ |
9+ |
25+ |
|||
|
Cypripedium calceolus |
c |
11+ |
121 |
8+ |
321 |
17+ |
|||
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
|
Fragaria vesca |
c |
17+ |
15+ |
13+ |
23+ |
4+ |
|||
|
Rumex acetosa s.l. |
c |
22+ |
4+ |
8+ |
5+ |
8+ |
|||
|
Ledum palustre |
c |
33+ |
22+ |
15+ |
8+ |
||||
|
Lathyrus pratensis |
c |
11+ |
21+ |
18+ |
21+ |
||||
|
Carex appropinquata |
c |
11+ |
33+ |
12+ |
|||||
|
Calamagrostis arundinacea |
c |
33+ |
18+ |
4+ |
|||||
|
Carex dioica |
c |
17+ |
22+ |
15+ |
|||||
|
C. chordorrhiza |
c |
17+ |
22+ |
12+ |
|||||
|
C. lasiocarpa |
c |
17+ |
22+ |
4+ |
|||||
|
Stellaria graminea |
c |
4+ |
8+ |
21+ |
|||||
|
Hieracium agg. vulgatum |
c |
22+ |
5+ |
||||||
|
Oxycoccus palustris |
c |
33+ |
15+ |
||||||
|
Vaccinium uliginosum |
c |
22+ |
13+ |
||||||
|
Carex aquatilis |
c |
131 |
33+ |
||||||
|
Sphagnum squarrosum |
d |
11+ |
33+ |
111 |
31+ |
21+ |
5+ |
8+ |
|
|
S. teres |
d |
11+ |
331 |
22+ |
151 |
4+ |
|||
|
S. russowii |
d |
339 |
11+ |
41 |
4+ |
||||
|
Straminergon stramineum |
d |
33+ |
11+ |
27+ |
21+ |
||||
|
Campylium stellatum |
d |
33+ |
8+ |
5+ |
8+ |
||||
|
Sphagnum wulfianum |
d |
33+ |
17+ |
12+ |
|||||
|
Drepanocladus polygamus |
d |
4+ |
4+ |
338 |
|||||
|
Campylium protensum |
d |
23+ |
8+ |
33+ |
|||||
|
Sphagnum capillifolium |
d |
222 |
|||||||
|
Rhizomnium punctatum |
d+z |
11+ |
331 |
11+ |
15+ |
4+ |
14+ |
21+ |
|
|
Plagiothecium denticulatum |
d+z |
33+ |
11+ |
8+ |
4+ |
9+ |
33+ |
||
|
Plagiochila asplenioides |
d+z |
221 |
8+ |
171 |
9+ |
4+ |
|||
|
Plagiomnium cuspidatum |
d+z |
11+ |
211 |
9+ |
29+ |
||||
|
P. affine |
d+z |
4+ |
212 |
4+ |
|||||
|
Plagiothecium laetum s.l. |
z+d |
11+ |
171 |
4+ |
13+ |
9+ |
25+ |
||
|
Breidleria pratensis |
z+d |
17+ |
33+ |
31+ |
13+ |
5+ |
17+ |
||
|
Ptilidium pulcherrimum |
z+d |
33+ |
19+ |
17+ |
9+ |
||||
|
Cladonia coniocraea |
z+d |
22+ |
15+ |
17+ |
4+ |
||||
|
Cirriphyllum piliferum |
z+d |
19+ |
8+ |
27+ |
33+ |
||||
|
Mnium stellare |
z+d |
4+ |
13+ |
5+ |
25+ |
||||
|
Tetraphis pellucida |
z |
11+ |
33+ |
12+ |
17+ |
14+ |
33+ |
||
|
Pleurozium schreberi |
z |
111 |
15+ |
251 |
33+ |
231 |
|||
|
Dicranum polysetum |
z |
17+ |
15+ |
4+ |
23+ |
13+ |
|||
|
Средние сомкнутос |
ть или п |
роектив |
ное пок |
ытие (% |
) ярусо |
в: |
|||
|
– 1-го яруса древостоя |
0.7 |
0.7 |
0.5 |
0.5 |
0.5 |
0.4 |
0.5 |
0.5 |
|
|
– 2-го яруса древостоя |
0.2 |
0.2 |
0.1 |
0.2 |
0.1 |
0.5 |
0.1 |
0.1 |
|
|
– подроста и подлеска |
20 |
10 |
15 |
15 |
15 |
45 |
15 |
15 |
|
|
– травяного |
60 |
60 |
45 |
70 |
70 |
75 |
75 |
60 |
|
|
– мохового |
70 |
65 |
65 |
60 |
65 |
90 |
70 |
65 |
|
|
Таблица 1 (окончание) |
||||||||
|
I II Ср |
III едняя в |
IV ысота яр |
V усов, м: |
VI |
VII |
VIII |
IX |
X |
|
– 1-го яруса древостоя |
21 |
21 |
20 |
20 |
19 |
18 |
23 |
23 |
|
– 2-го яруса древостоя |
13 |
8 |
11 |
12 |
12 |
10 |
11 |
17 |
|
– подроста и подлеска в целом |
2.1 |
3.0 |
3.1 |
4.0 |
2.7 |
2.8 |
2.7 |
3.5 |
|
– 1-го подъяруса |
3.7 |
3.8 |
? |
5.8 |
? |
? |
? |
? |
|
– 2-го подъяруса |
1.3 |
0.9 |
? |
1.7 |
? |
? |
? |
? |
|
– 3-го подъяруса |
0.5 |
— |
? |
0.5 |
? |
? |
? |
? |
|
Бонитет древостоя |
III |
III |
III |
III |
III |
IV |
II |
II |
|
Мощность торфа, см |
30 |
125 |
215 |
120 |
95 |
40 |
90 |
95 |
|
Число описаний |
9 |
6 |
9 |
26 |
24 |
3 |
22 |
24 |
Примечания . Синтаксоны: 1–6 – Piceetum (P.) filipenduloso-caricosum cespitosae: 1 – myrtillosum; 2 – caricosum loliaceae; 3 – moliniosum caeruleae; 4–5 – typicum: 4 – var. Frangula alnus, 5 – var. Ranunculus subboreal-is; 6 – caricosum juncellae; 7–8 – P. sphagno warnstorfii-aconitosum: 7 – var. typica, 8 – var. Calliergon richardsonii. Ярусы: a1 и a2 – 1-й и 2-й ярусы древостоя, b – подрост и подлесок, c – кустарничково-травяной, d – моховой, z – эпифитные и эпиксильные мхи и лишайники. Для видов приводятся постоянство (%) и (в надстрочном регистре) среднее проективное покрытие (%). Детерминантные группы выделены серым фоном и жирной рамкой; виды в их пределах сортированы по ярусам, далее по убыванию встречаемости во всем массиве описаний таблицы. Для доминирующих видов значения покрытия и постоянства даны полужирным шрифтом. Среднее проективное покрытие видов менее 0.5% отмечено плюсом «+». Прочерк «–» – отсутствие подъяруса, знак вопроса «?» – отсутствие данных. Исключены сопутствующие виды со встречаемостью менее 20% в каждом из синтак-сонов, всходы деревьев и кустарников.
Несмотря на сходство рассматриваемых ассоциаций с ельниками травяными, авторы затрудняются с окончательным выводом об их принадлежности к травяно-сфагновой либо травяной группе. Для этого нужны результаты завершенной классификации в каждой из групп.
Благодаря высокому уровню минерального богатства почв древостои ельников обеих ассоциаций отличают повышенные для заболоченных лесов сомкнутость и производительность: в лесах большинства подчиненных синтаксонов бонитет достигает II–III. Древостой дифференцирован по высоте на два, в южной тайге иногда на три яруса. В 1-м ярусе выражена примесь Betula pubescens s.l. к Picea abies s.l. от 1 до 3–4 единиц по составу в зависимости от синтаксона. Стратификация на 3 подъяруса типична и для подроста и подлеска. Однако высота подъярусов оценена не для всех синтаксонов. Нижние подъярусы подлеска умеренно густые, богаты видами орнитохорных кустарников (Frangula alnus, Ribes nigrum, R. spicatum, R. alpinum, Lonicera pallasii s.l., L. xylosteum, Viburnum opulus). Подрост еловый с примесью Betula pubescens s.l., как правило, немногочисленный. ПП травяного яруса не превышает 60–75% (табл. 1) в отличие от ельников травяных, где оно часто достигает 90% и более [10, 14, 42].
Таблица 2
Амплитуды среднемноголетних (1983–2004) значений метеопараметров в пределах ареалов синтаксонов мезоэвтрофных ельников таволгово-дернистоосоковых и аконитово-таволговых сфагновых
|
Синтаксоны |
Метеопараметры |
|
|
GDD>10, °C |
K |
|
|
1. Piceetum (P.) filipenduloso-caricosum cespitosae |
468–686 |
29.33–44.44 |
|
subass. myrtillosum |
||
|
2. Caricosum loliaceae |
484–596 |
35.22–40.19 |
|
3. Moliniosum caeruleae |
460–546 |
33.81–35.24 |
|
4. Typicum var. Frangula alnus |
285–727 |
33.81–40.32 |
|
5. Var. Ranunculus subborealis |
245–702 |
38.35–46.25 |
|
6. Caricosum juncellae |
447–452 |
37.8–39.12 |
|
7. P. sphagno warnstorfii-aconitosum var. typica |
484–596 |
35.65–39.52 |
|
8. Var. Calliergon richardsonii |
447–589 |
37.37–43.37 |
Примечания . GDD>10 - сумма градусо-дней выше 10 ° С [43], K - коэффициент континентальности Конрада [47]. Амплитуды рассчитаны с учетом как имеющейся совокупности описаний, так и данных литературы.
Детерминанты, свойственные обеим ассоциациям, подразделяются на три экологические группы. К первой относятся мезотрофные ( Rosa acicularis, Equisetum pratense, Carex vaginata, Rubus saxatilis, Geranium sylvaticum s.l.) и ме-зоэвтрофные ( Juniperus communis s.l., Paris quadrifolia ) лесные мезофиты. Это бореальные, отчасти также бореонеморальные и полизональ-ные виды, приуроченные к повышениям нанорельефа. Ко второй группе относятся бореонемо-ральные мезоэвтрофные лесные и лугово-лесные гигромезофиты ( Padus avium s.l., Geum rivale, Angelica sylvestris ), к третьей – полизональные мезотрофные ( Comarum palustre, Plagiomnium ellipticum, Climacium dendroides, Calliergon cordifolium ) и мезоэвтрофные ( Carex cespitosa, Galium palustre ) болотно-лесные мезогигрофиты и гигрофиты. Экологическая неоднородность детерминантов обусловлена изменчивостью экологических режимов по элементам дифференцированного нанорельефа, хорологическая – указывает на многоэтапность формирования соответствующих ценофлор в голоцене [15].
К списку константных видов (см. выше) добавляются Sorbus aucuparia s.l., Alnus incana , Equisetum sylvaticum, Melica nutans, Maianthemum bifolium, Viola epipsila, Chamaenerion angustifoli-um, Trientalis europaea, Aulacomnium palustre, Rhytidiadelphus triquetrus (табл. 1). Константные доминанты приземных ярусов – болотно-лесные ( Calamagrostis purpurea s.l., Sphagnum warnstorfii ) и лугово-лесные ( Filipendula ulmaria s.l.) гигромезо- и мезогигрофиты, в основном мезоэвтрофы. Среди прочих констант, включая доминирующие виды древесного яруса, по числу видов преобладают бореально-лесные мезофиты, растения темнохвойной тайги.
Торфяная залежь полностью либо только в верхней части сложена низинными древесными торфами высокой (40–50% и более) степени разложения. Ведущим торфообразователем является Picea abies s.l., обильны также остатки Betula pubescens s.l., Carex cespitosa и Equisetum spp. На Sphagnum warnstorfii, Filipendula ulmaria s.l. и Calamagrostis purpurea s.l. может приходиться значительная доля растительных остатков в относительно слаборазложенном поверхностном слое залежи. Однако по мере усиления разложения с глубиной доля остатков этих видов снижается, в силу чего их нельзя относить к основным торфообразователям в сообществах данного типа. Неглубокие (до 1 м) залежи обычно сверху донизу сложены древесным и древесноосоковым (с участием Carex cespitosa ) торфом.
Более глубокие залежи также могут быть целиком сложены данными торфами, но чаще с глубиной последние сменяются низинными древесно-травяно-сфагновыми, осоково- и травяносфагновыми, травяно-гипновыми и иными разностями торфов меньшей степени разложения, свидетельствующими о том, что на месте современных ельников некогда существовали безлесные низинные болота. В этом случае можно говорить об имевшей место эндэкзогенной смене болотного сообщества на лесное, связанной с перераспределением болотного стока, формированием ручьевых русел и последующим развитием дренажа.
«Центр тяжести» ареалов обеих ассоциаций находится в средней тайге, хотя сообщества ряда подчиненных синтаксонов проникают и в северную часть южнотаежной подзоны и/или в южную полосу подзоны северной тайги (рис. 1, 2).
ЕЛЬНИКИ ТАВОЛГОВОДЕРНИСТООСОКОВЫЕ СФАГНОВЫЕ
Piceetum (P.) filipenduloso-caricosum cespi-tosae (Filipendulo-Carici cespitosae-P.) – ельник (е.) таволгово-дернистоосоковый сфагновый (табл. 1, рис. 1: № 1–6). Леса большинства подчиненных синтаксонов с равной частотой произрастают на силикатных и на карбонатных породах. Почвы от дерново-подзолисто-грунтовоглеевых [16], торфянисто- и торфяно-глеевых до болотных низинных на маломощных и среднемощных, реже на мощных перегнойных торфах [4, 27], слабокислые (pH H2O 5.8–6.0 [2]). Нанорельеф кочковатый, обычно трехкомпонентный с участками моховых ковров и сырыми западинами, реже двухкомпонентный без западин. Его формированию способствуют как нарастание кочек Carex cespitosa и других видов кочкообразующих осок, так и развитие ветровальнопочвенного комплекса. Бонитет древостоя обычно III. Дифференциация ассоциации от ельников аконитово-таволговых сфагновых в основном негативная – по отсутствию видов из группы Ribes nigrum–Aconitum septentrionale (см. выше). Собственная детерминантная группа ассоциации включает олигомезотрофные виды, более обычные в ельниках хвощовых со Sphagnum girgen-sohnii [13] и таволгово-хвощовых сфагновых. Это Pinus sylvestris и Sphagnum centrale (табл. 1) .
В южной части подзоны южной тайги и подтайге ельники таволгово-дернистоосоковые, как и многие другие ассоциации мезоэвтрофных ельников травяно-сфагновых, замещаются чер-ноольшанниками. За пределами России ассоциация известна из Финляндии [40] и Эстонии [45], но не отмечена ни в Западной Фенноскандии [46], ни в бореально-лесном поясе гор Средней Европы [39, 44, 48].
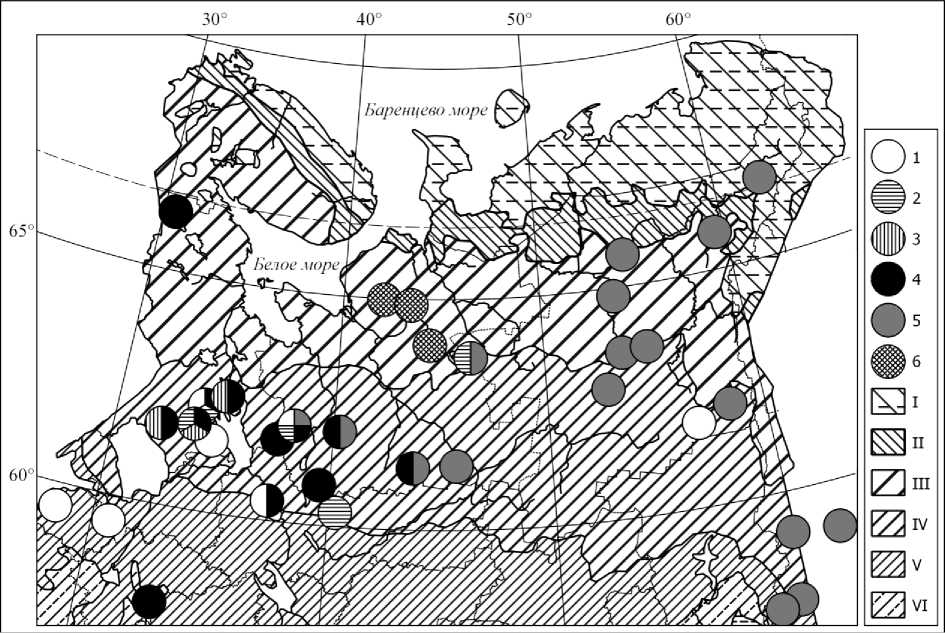
Рис. 1. Распространение ельников дернистоосоково-черничных (1), таволгово-плевельноосоковых (2), таволгово-молиниевых (3), таволгово-дернистоосоковых сфагновых западного (4) и восточного (5) вариантов и ивняковых таволгово-вилюйскоосоковых (6) в Европейской России и на Урале.
Зоны и подзоны: I – тундра, II – лесотундра и подгольцовые редколесья, III – северная, IV – средняя, V – южная тайга, VI – подтайга (картооснова [5, 26] объединяет зональные выделы и их высотно-поясные аналоги)
В южной части подзоны южной тайги и подтайге ельники таволгово-дернистоосоковые, как и многие другие ассоциации мезоэвтрофных ельников травяно-сфагновых, замещаются чер-ноольшанниками. За пределами России ассоциация известна из Финляндии [40] и Эстонии [45], но не отмечена ни в Западной Фенноскандии [46], ни в бореально-лесном поясе гор Средней Европы [39, 44, 48].
В составе ассоциации выделяются пять субассоциаций.
Subass. myrtillosum – е. дернистоосоковочерничный сфагново-зеленомошный (табл. 1, рис. 1: № 1). Сообщества соответствуют начальному этапу мезоэвтрофного заболачивания, развиты в нижней части и на шлейфах пологих склонов водоразделов и боровых террас, реже близ выходов ключей на надпойменных террасах. Почвы различного гранулометрического состава от песков до глин. Торфяная залежь обычно неглубокая. Нанорельеф с малым числом западин либо без них; по площади могут преобладать как низкие (10–40 см) кочки, так и ковры. Сомкнутость 1-го яруса древостоя близка к нормальной. Примесь Betula pubescens s.l. к Picea abies s.l. до 1, Pinus sylvestris – менее 1 единицы по составу яруса. Во втором ярусе и подросте почти исключительно Picea abies s.l. Подрост невысокий, но вдвое более густой, чем в лесах остальных обсуждаемых синтаксонов. В подлеске обильнее всего Sorbus aucuparia s.l.; в нижнем подъярусе постоянна Rosa acicularis. Прочие кустарники встречаются эпизодически. В травянокустарничковом ярусе микроповышений преобладает Vaccinium myrtillus в сопровождении V. vitis-idaea, Rubus saxatilis, Oxalis acetosella, Linnaea borealis, Gymnocarpium dryopteris, Dryopteris carthusiana. На участках ковров постоянны, но не обильны Filipendula ulmaria s.l., Carex cespitosa, Geranium sylvaticum s.l., Equisetum palustre, E. sylvaticum, Calamagrostis purpurea s.l., Crepis paludosa, Cirsium heterophyllum. Иногда субдоминантом становится Equisetum pratense. В ярусе мхов согоспод-ствуют Hylocomium splendens и Pleurozium schreberi с примесью Rhytidiadelphus triquetrus. Sphagnum warnstorfii может замещаться на S. girgensohnii, достигающий при этом большего обилия, нежели первый вид.
Детерминанты – олигомезотрофные пустошно-лесные (Avenella flexuosa s.l.) и мезотрофные лесные (Dicranum majus) мезофиты. В группу, сближающую субассоциацию с ельниками таволгово-плевельноосоковыми, входят лесные мезофильные (D. polysetum) и болотно-лесные гигромезофильные (Sphagnum girgensohnii, Polytrichum commune) мхи, от олиготрофов до мезотрофов, типичные для мезотрофных ельников сфагновых [12, 13]. Негативная дифференциация субассоциации обусловлена отсутствием видов из групп Salix pentandra–Caltha palustris и Carex disperma–C. loliacea (табл. 1).
Большинство описаний ельников субассоциации тяготеет к средней тайге Карелии, юго-запада Архангельской и северо-запада Вологодской областей. Одно из описаний, однако, было выполнено в Печорском Предуралье. Это говорит как о протяженном ареале синтаксона, так и о повсеместной редкости его сообществ (возможно, в силу их сукцессионной природы). Вероятно, аналогичные леса могут быть обнаружены и в других среднетаежных регионах Республики Коми, а также севера Кировской обл. В южной тайге аналогичные ельники описаны на юго-западе [29] и в центральной части Ленинградской обл.
Subass. caricosum loliaceae – е. таволговоплевельноосоковый сфагновый (табл. 1, рис. 1: № 2). Сравнительно редкие сообщества логовых понижений и окраин болот. Почвы супесчаные или суглинистые, обычно с мощной (до 2 м и более) торфяной залежью. Нанорельеф трехкомпонентный с преобладанием ковров и развитыми западинами, что отражает сильную степень заболоченности, но высота кочек при этом как в предыдущем синтаксоне. В древостое примесь Betula pubescens s.l. возрастает до 3 единиц по составу 1-го яруса; 2-й ярус низкий. Подрост Picea abies s.l. несколько выше, но вдвое менее густой, чем в ельниках дернистоосоковочерничных. Снижается и общее покрытие подроста и подлеска, а также ПП Sorbus aucuparia s.l. при одновременном росте встречаемости Padus avium. В остальном характеристики древостоя, подроста и подлеска как в предыдущей субассоциации.
Доминантами травяно-кустарничкового яруса выступают Carex loliacea и Vaccinium vitis-idaea ; V. myrtillus переходит на роль субдоминанта наряду с Athyrium filix-femina . Filipendula ulmaria s.l. и Carex cespitosa остаются в роли не слишком обильных сопутствующих видов, так же как Oxalis acetosella, Rubus saxatilis, Dryopteris carthusiana , Equisetum sylvaticum и Calamagrostis purpurea s.l. В моховом ярусе микроповышений и ковров господствуют соответственно Pleurozium schreberi и Sphagnum warnstorfii ; последний, как и в предыдущем синтаксоне, изредка замещается на S. girgensohnii . S. centrale становится одним из субдоминантов вместе с Hylo-comium splendens . В западинах появляются
Pseudobryum cinclidioides и Calliergon cordifolium .
От ельников дернистоосоково-черничных субассоциацию отличают две группы детерминантов, одновременно сближающие ее с ельниками таволгово-молиниевыми. Первая из групп объединяет синтаксон и со всеми прочими сильно заболоченными (сфагновыми) субассоциациями и вариантами ельников, обсуждаемых в данной статье. Ее формируют мезотрофные ( Epilobium palustre ) и мезоэвтрофные ( Salix pentandra, Caltha palustris, Pseudobryum cinclidioides ) болотно-луговые и болотно-лесные гигрофиты. Ценоспектр таежно-лесных мезотрофных ( Carex disperma ) и мезоэвтрофных ( C. loliacea ) гигромезофитов из состава второй группы не столь широк: они «выпадают» в северотаежных ельниках таволгово-вилюйскоосоковых. В то же время субассоциацию сближает с ельниками дернисто-осоково-черничными и отличает от таволгово-молиниевых группа Dicranum polysetum– Sphagnum girgensohnii (табл. 1).
Ареал субассоциации выявлен не полностью. Основная часть относящихся к ней ельников описана в средней тайге Карелии (в том числе в заповеднике «Кивач» [18]) и юго-запада Архангельской обл. (национальный парк (НП) «Кено-зерский»), где растительность изучалась наиболее интенсивно. Отдельные описания сделаны также в северной тайге Архангельской обл. (бассейн р. Нюхчи, притока р. Пинеги) и в южной тайге Вологодской обл. на водоразделе рек Во-жега и Сить [2].
Subass. moliniosum caeruleae – е. таволгово-молиниевый сфагновый (табл. 1, рис. 1: № 3). Сообщества средней тайги Карелии. Развиваются в скрытопроточных межозовых и межсельговых понижениях, а также вдоль ручьев, текущих по краю болот, всегда по мощной (до 5 м) торфяной залежи поверх песка со щебнем или скального водоупора. Нанорельеф с преобладанием ковров; западины отсутствуют или их мало. Кочки невысокие (20–40 см). По сравнению с субассоциациями, описанными выше, сомкнутость и средняя высота 1-го яруса древостоя несколько снижены. Примесь Betula pubescens s.l. к Picea abies s.l. приближается к 4 единицам по составу яруса, Pinus sylvestris – единична. Второй ярус древостоя выражен слабо. ПП яруса подроста и подлеска промежуточное между таковым в двух предыдущих субассоциациях. Подрост березовоеловый, малочисленный. В подлеске наибольшее ПП у Juniperus communis; постоянны, но не обильны Sorbus aucuparia s.l., Frangula alnus и Rosa acicularis. Доминанты травяного яруса – Molinia caerulea и Filipendula ulmaria s.l. Carex cespitosa – по-прежнему сопутствующий вид на моховых коврах вместе с Geum rivale, Crepis paludosa, Calamagrostis phragmitoides либо C. canescens и (иногда) Phragmites australis. На микроповышениях постоянны Vaccinium vitis-idaea, V. myrtillus, Carex vaginata вместе с Convallaria majalis, Geranium sylvaticum и Rubus saxatilis; умеренно обильны Pleurozium schreberi и Hylocomium splendens. В моховом ярусе ковров к доминирующему Sphagnum warnstorfii эпизодически примешиваются S. centrale и S. angustifolium; S. girgensohnii при этом нехарактерен. Plagiomnium ellipticum, Rhizomnium pseudopunctatum и Pseudobryum cinclidioides в малом обилии присутствуют в западинах, первые два вида – также в примеси в ковре.
Ельникам таволгово-молиниевым свойственна многосторонняя дифференциация. «Ядро» группы их детерминантов составляют полизо-нальные мезоэвтрофные ( Molinia caerulea, Deschampsia cespitosa, Carex flava ) и бореальные мезотрофные ( Dactylorhiza maculata, Potentilla erecta ) лугово- и болотно-лесные гигромезофиты, в основном субокеанические виды с западноевропейско-поволжско-южнозападносибирским ареалом [11]. К группе тяготеют и виды иной экологии – от олиготрофных и олигомезотроф-ных мезогигрофитов ( Carex paupercula, Sphagnum angustifolium ) до эвтрофных гигрофитов ( Phragmites australis ). Бореонеморальные ме-зотрофные мезофиты ( Convallaria majalis ) и мезогигрофиты ( Frangula alnus ) сближают субассоциацию с западным вариантом типичных ельников таволгово-дернистоосоковых, западинные мезотрофные ( Carex canescens ) и мезоэвтрофные ( Equisetum fluviatile ) гигрофиты – также с восточным их вариантом и с ельниками таволгово-вилюйскоосоковыми, мезоэвтрофный гигромезофит Salix myrsinifolia – только с последними.
Как с типичными ельниками таволговодернистоосоковыми, так и с аконитовотаволговыми синтаксон объединяет экологически неоднородная группа видов, в составе которой преобладают олигомезотрофные (Rubus chamaemorus, Moneses uniflora) и мезотрофные (Rubus arcticus, Pyrola minor, P. rotundifolia, подрост Betula pubescens s.l.) мезо- и гигромезофиты микроповышений. Однако в эту же группу входят и западинные мхи, мезогигрофиты (Helodium blandowii) и гигрофиты (Calliergon giganteum, Calliergonella cuspidata). Наконец, мезоэвтроф-ный луговой мезофит Galium boreale служит детерминантом как ельников таволгово-молиниевых, так и таволгово- вилюйскоосоковых, а также восточных вариантов аконитово-таволговых и типичной субассоциации таволгово-дернистоосоковых. Отсутствие видов из группы Dicranum polysetum–
Sphagnum girgensohnii отличает рассматриваемую субассоциацию от плевельноосоковой, из группы Ptilium crista-castrensis–Dicranum fuscescens – также от западного варианта типичных ельников таволгово-дернистоосоковых (табл. 1).
Ельники таволгово-молиниевые описаны в Пряжинском, отчасти также в Кондопожском и Медвежьегорском районах Карелии. В бывшем природном парке «Хийсъярви» и близ п. Колатсельга они встречаются наряду с олиготрофными ельниками деренно-морошковыми сфагновыми, где также бывает обильна Molinia caerulea [12, 38]. В Финляндии мезоэвтрофные ельники с молинией отмечены не были [40, 46].
Subass. typicum – собственно е. таволговодернистоосоковый сфагновый (табл. 1, рис. 1: № 4–5). Центральная субассоциация, дифференциация которой негативна – по отсутствию детерминантов как ельников таволгово-молиниевых, так и вилюйскоосоковых. Почвы различного гранулометрического состава от песчаных до глинистых; мощность торфа варьирует от 20–40 см до 2–4 м. Высота кочек 20–70 см. Примесь Betula pubescens s.l. к Picea abies s.l. до 2–3 единиц по составу 1-го яруса древостоя. Сомкнутость и высота яруса – как в предыдущем синтаксоне, так же как ПП подроста и подлеска. При этом сомкнутость 2-го яруса древостоя – скорее как в ельниках дернистоосоковочерничных и плевельноосоковых. Подрост малочисленный, еловый с примесью Betula pubescens s.l. В подлеске постоянны Juniperus communis и Rosa acicularis . В травяном ярусе Carex cespitosa становится одним из доминантов, разделяя господство с Filipendula ulmaria s.l. В числе сопутствующих видов яруса обычны Calamagrostis purpurea s.l., Crepis paludosa , Viola epipsila , Angelica sylvestris , Equisetum palustre , Caltha palustris , Comarum palustre , на микроповышениях – Vaccinium vitis-idaea, V. myrtillus, Oxalis acetosella, Gymnocarpium dryopteris, Trientalis europaea, Maianthemum bifolium, Linnaea borealis, Orthilia secunda, в средней тайге Rubus arcticus, Carex disperma. В моховом ярусе микроповышений преобладают Hylocomium splendens и Pleurozium schreberi с примесью Rhytidiadelphus triquetrus, Dicranum scoparium и Climacium dendroides . К Sphagnum warnstorfii , формирующему моховые ковры, примешивается Plagiomnium ellipticum , редко также Sphagnum centrale . В западинах постоянны Calliergon cordifolium и Pseudobryum cinclidioides (табл. 1).
Выделяются два варианта, замещающих друг друга с запада на восток.
Var. Frangula alnus – западный вариант (табл. 1, рис. 1: № 4). Сообщества скрытопроточ- ных понижений, реже встречаются по окраинам ключевых болот на надпойменных террасах и по берегам ручьев. Как правило, заболочены умеренно, реже сильно. В структуре нанорельефа по площади могут преобладать как кочки, так и ковры; западин обычно мало. Второй ярус древостоя сравнительно высокий, как в ельниках дернистоосоково-черничных. Еловый подрост отличается хорошим ходом роста, его верхний подъярус превышает 5 м. К списку постоянных видов подлеска добавляются Sorbus aucuparia, Frangula alnus, Alnus incana, Padus avium. Аналогичный список сопутствующих видов травяного яруса пополняют Solidago virgaurea, Dryopteris carthusiana, Rubus saxatilis, Convallaria majalis, Melica nutans и Carex vaginata на приствольных повышениях, Athyrium filix-femina на коврах, Equisetum fluviatile и Galium palustre в западинах. Calamagrostis phragmitoides может замещаться на С. canescens.
От восточного варианта западный отличают как отсутствие видов из детерминантных групп, свойственных первому, так и группа Frangula alnus–Convallaria majalis. Дифференциация от ельников таволгово-молиниевых – по отсутствию их детерминантов, а также по нетипично-сти Galium boreale и Salix myrsinifolia (табл. 1).
Ельники западного варианта повсеместны в средней тайге Карелии, а также Архангельской обл. к западу от р. Северной Двины. В частности, опубликовано описание с окраины водораздельного болота в Коношском р-не (P. caricoso-sphagnosum [36]). На северо-западе Вологодской обл. (Вашкинский р-н) сообщества отмечены как в средней, так и в южной тайге, соответственно при основании склона флювиогляциальной гряды и на карбонатной морене. Второе южнотаежное описание сделано в НП «Валдайский» на юге Новгородской обл. (Climacio-Piceetum typicum var. typica Korotk. 1991 [7]). Кроме того, вариант приводится для всей южной тайги Ленинградской обл. как «ельник дернистоосоковый» [17]. Аналогичные сообщества с Carex cespitosa, Fili-pendula ulmaria, Geum rivale, Sphagnum warn-storfii, Calliergon cordifolium, Pseudobryum cin-clidioides, Plagiomnium ellipticum известны из Финляндии (включая и северную тайгу) как ground-water-influenced herb and grass birch-spruce mires [40], а из Эстонии – как Filipendulo-Piceetum (Filipendula site type [45]). Изолированное местонахождение синтаксона отмечено в горной тайге НП «Паанаярви» на северо-западе Карелии.
Var. Ranunculus subborealis – восточный вариант (табл. 1, рис. 1: № 5). По окраинам болот и заболоченным долинам ручьев, на Урале в нижней части и на шлейфах горных склонов. Нано- рельеф двухкомпонентный, без ковров, с сопоставимой долей кочек и западин, реже трехкомпонентный с преобладанием кочек. По сравнению с предыдущим вариантом сообщества более обводнены; подрост ели низкий и более разреженный. Второй ярус древостоя также невысокий, как в ельниках плевельноосоковых. Sorbus aucuparia s.l., Alnus incana и Padus avium встречаются эпизодически; Frangula alnus и Cala-magrostis canescens нехарактерны. К списку постоянных сопутствующих видов травяного яруса, свойственных субассоциации в целом, добавляются Geranium sylvaticum s.l., Equisetum sylvati-cum и Rubus chamaemorus. В западинах возрастает ПП Pseudobryum cinclidioides.
Группа мезотрофных ( Lonicera pallasii s.l.) и мезоэвтрофных ( Veratrum lobelianum, Atragene sibirica ) лесных и лугово-лесных мезофитов сближает вариант с ельниками вилюйскоосоко-выми и с хорологически сходным восточным вариантом ельников аконитово-таволговых. Другая, объединяющая мезотрофные мезо- и гигро-мезофильные виды ив ( Salix caprea, S. phylicifolia ) с мезоэвтрофным гигромезофитом Bistorta major , характерна и для западного варианта последних. С ассоциацией ельников аконитово-таволговых в целом вариант объединяют мезоэвтрофные лугово-лесные и лесные мезофиты ( Lathyrus vernus ) и гигромезофиты ( Trollius europaeus, Ranunculus subborealis, Cirsium oleraceum ), к которым присоединяется эпифитный (в сообществах данного синтаксона) мезо-трофный мезофильный мох Sanionia uncinata. Одновременно названные виды отличают син-таксон от ельников вилюйскоосоковых наряду с отсутствием детерминантов последних. Виды всех трех названных детерминантных групп, а также Galium boreale дифференцируют восточный вариант от западного, равно как и отсутствие видов из группы Frangula alnus–Convallaria majalis (табл. 1).
Леса восточного варианта обычны в средней тайге и в южной полосе подзоны северной тайги Архангельской обл. и Республики Коми. В первом из регионов большая часть описаний сделана к востоку от Северной Двины. Однако отдельные регистрации сообществ есть и к западу от нее, где ареалы вариантов перекрываются, – в средней тайге Устьянского и Няндомского районов и в НП «Кенозерский» на юго-западе области. В Республике Коми ряд находок сделан и в крайнесеверной тайге Предуралья (Воркута-Вом) и Приполярного Урала (Кожим; P. caespitosi-caricoso-herbo-sphagnosum (Yudin, 1948)). В южной полосе северной тайги вариант был описан на Среднем Тимане (P. filipenduloso-sphagnosum со Sphagnum girgensohnii [1], P. mixto-herboso- sphagnosum [37]) и Средней Печоре (P. caricosum [25]), а в средней тайге – в бассейне р. Нившеры (P. filipenduloso-sphagnosum [37]). На Верхней Печоре леса синтаксона распространены вплоть до предгорий Северного Урала. В средней тайге Среднего Урала и равнинного Зауралья они отмечены и в Свердловской обл., где встречаются в понижениях пойм, у выходов ключей, по окраинам осоковых болот. Для них характерны Carex cespitosa, Filipendula ulmaria, Calamagrostis lang-sdorffii, Equisetum sylvaticum, Geum rivale, Vera-trum lobelianum, Caltha palustris, Thalictrum ke-mense (P. caricosum, Alneto-Betuleto-Piceetum caricosum [19]). Ельники таволговодернистоосоковые с мощным ковром сфагновых мхов (P. caricoso-sphagnosum) описаны также в южной тайге области: в Восточном Предуралье и в Висимском заповеднике на Среднем Урале [23].
Без деления на варианты сообщества субассоциации приводятся для всей средней и южной тайги Европейской России как P. filipenduloso-sphagnosum [23], однако под этим же названием приводятся и ельники таволгово-хвощовой ассоциации P. equisetoso-filipendulosum (см. выше). Южносибирский P. caespiti-caricoso-sphagnosum [23], судя по доминированию Sphagnum girgen-sohnii и значимому покрытию Equisetum sylvati-cum и Calamagrostis langsdorffii , также должен быть отнесен к ельникам таволгово-хвощовым сфагновым.
Subass. caricosum juncellae – е. ивняковый таволгово-вилюйскоосоковый сфагновый (табл. 1, рис. 1: № 6). В отличие от других син-таксонов, леса этой субассоциации встречаются лишь в северной тайге Архангельской обл. в районе залегания сульфатных и карбонатных пород. Они описаны на легко- и среднесуглинистых почвах в заболоченных понижениях надпойменной террасы р. Кулой [10], также известны из надпоймы других рек Келдинско-Полтинского и Пинежско-Мезенского ландшафтов (P. equisetoso-caricosum [24]). Торфяная залежь в конце июня с мерзлотой на глубине 30– 50 см; в первой декаде июля мерзлота протаивает. Выражен двухкомпонентный нанорельеф с преобладанием западин и отстоящими друг от друга осоковыми кочками и буграми высотой 50–150 см. Первый ярус древостоя разрежен, высота его снижена; в отличие от других субассоциаций бонитет снижается до IV. Примесь Betula pubescens s.l. к Picea abies s.l. около 3 единиц по составу яруса. Сомкнутость 2-го яруса древостоя выше, чем у 1-го; этот ярус сформирован Alnus incana, Salix myrsinifolia, S. jenisseensis и другими видами ив, в основном мезоэвтрофными. В густом подлеске также преобладают виды Salix
( S. phylicifolia, S. lapponum, S. hastata ). Вместе с тем обилен и Juniperus sibirica ; постоянна Lonic-era pallasii s.l. Еловый подрост малочисленный, низкий. В травяном ярусе господствует Carex juncella , тогда как C. cespitosa переходит в число сопутствующих констант. Filipendula ulmaria выступает доминантом 2-го порядка, при этом растет на кочках Carex juncella. Таволге сопутствуют виды евросибирского высокотравья ( Veratrum lobelianum, Atragene sibirica, Ligularia sibirica, Thalictrum kemense ), а также Pyrola ro-tundifolia, Carex vaginata, Rubus arcticus, Saus-surea alpina, Parnassia palustris, Veronica longifolia . Еще один доминант 2-го порядка – за-падинный Comarum palustre , сопровождаемый Naumburgia thyrsiflora, Equisetum fluviatile и Cicuta virosa . В моховом покрове обводненных западин обильны мхи-гигрофиты ( Calliergon giganteum, C. cordifolium, Pseudobryum cinclidioides ). Доминирующий в составе яруса Sphagnum warnstorfii произрастает как в западинах при основании кочек и бугров, так и по стенкам последних совместно с Plagiomnium ellipticum и P. rostratum . На вершинах бугров и при стволах обилен Hylocomium splendens ; ему сопутствуют Rhytidiadelphus triquetrus и Climacium dendroides .
В числе детерминантов субассоциации мезо-трофные (Salix lapponum, Carex juncella, Cicuta virosa, Naumburgia thyrsiflora, Pedicularis sceptrum-carolinum) и мезоэвтрофные (Cardamine dentata, Veronica longifolia) мезогигрофиты и за-падинные гигрофиты сочетаются с мезоэвтроф-ными гигромезофитами (Salix jenisseensis, S. hastata, Parnassia palustris, Lathyrus palustris, Ligularia sibirica, Saussurea alpina, Plagiomnium rostratum) и мезофитами (Thalictrum kemense). К группе примыкают также олигомезотрофные (Orthilia obtusata) и мезотрофные (Peltigera aphthosa s.l.) мезофиты, растущие на микроповышениях. Мхи Bryum pseudotriquetrum (мезоэв-трофный мезогигрофит) и Rhytidiadelphus subpinnatus (мезотрофный гигромезофит) объединяют синтаксон с восточным вариантом ельников аконитово-таволговых. В то же время наблюдается многосторонняя негативная детерминация субассоциации, что отражает ее экологическую и географическую специфику. Так, для нее нехарактерны многие олигомезотрофные (Gymnocarpium dryopteris, Lycopodium annotinum, Vaccinium myrtillus, Pleurozium schreberi) и мезо-трофные (Daphne mezereum, Dryopteris carthusiana, Luzula pilosa, Oxalis acetosella, Solidago virgaurea s.l., Dicranum scoparium) лесные мезофиты микроповышений, а также ряд гигромезофитов (Athyrium filix-femina, Cirsium heterophyllum, Crepis paludosa, Rhizomnium pseudopunctatum). Все эти виды постоянны во всех остальных рассматриваемых синтаксонах. Отсутствуют и мхи-мезофиты из группы Ptilium crista-castrensis–Dicranum fuscescens, равно как и виды из групп Carex disperma–C. loliacea и Lathy-rus vernus–Cirsium oleraceum (табл. 1).
Сообщества произрастают в сочетании с пойменными ельниками ивняковыми таволгово-вилюйскоосоковыми с моховым покровом из Cal-liergonella lindbergii , C. cuspidata, Plagiomnium elatum и Climacium dendroides без участия Sphagnum warnstorfii и развитой торфяной залежи. Их отличает сомкнутый 2-й ярус древостоя из Salix pyrolifolia, S. cinerea, S. jenisseensis и Alnus incana , а также согосподство Filipendula ulmaria, Carex juncella и Comarum palustre при участии Caltha palustris, Naumburgia thyrsiflora , Equisetum fluviatile и (на кочках) E. pratense в травяном ярусе (Fil-ipendulo-P. comaretosum palustris различных вариантов [10]). Пока не совсем ясно, относятся ли эти сообществ к одной ассоциации с вышерассмотренной subass. caricosum juncellae, представляя собой лишь разные подчиненные синтаксоны, или же к разным ассоциациям и разным их группам.
Сходные с subass. caricosum juncellae типы сообществ известны из северной тайги Республики Коми [37], но в целом распространение их не выявлено. Редкостойные ельники вилюйско-осоковые сфагновые с восточного макросклона Полярного Урала с согосподством Carex juncella и Calamagrostis purpurea с примесью Bistorta el-liptica, Sanguisorba officinalis, а также Sphagnum fallax к S. warnstorfii в моховом ковре (Grumi-piceetum wiluicosum [30]), вероятно, относятся к иному синтаксону. В частности, они произрастают на силикатах и лишены подлеска из мезоэв-трофных видов Salix . То же справедливо и в отношении кольских ельников вилюйскоосоковых, отмеченных в Хибинах (Sparse-Piceetum fontinale [8]) и в Лапландском заповеднике. Окончательное выявление объема субассоциации caricosum juncellae требует привлечения дополнительных данных.
ЕЛЬНИКИ АКОНИТОВО-ТАВОЛГОВЫЕ СФАГНОВЫЕ
P. sphagno warnstorfii-aconitosum (Sphagno warnstorfii-Aconito-P.) – е. аконитово-таволговый сфагновый (табл. 1, рис. 2: № 7–8). Синоним: Filipendulo-P. aconitetosum var. Sphagnum warnstorfii [14]. Основная часть ареала ассоциации приурочена к области залегания карбонатных пород на северном макросклоне Русской равнины. Чаще всего рассматриваемые ельники встречаются именно на этих породах, но в поймах рек могут произрастать и на силикатных отложениях. Почвы от песчаных до тяжелосугли- нистых, умеренно кислые (pHH2O 5.4) пойменные иловато-торфянисто-глеевые [2] и перегнойноглеевые, реже торфянисто- и торфяно-глеевые низинные [16, 27]. Торф обычно неглубокий, но по окраинам болот может достигать мощности 2–4 м. При обилии Aconitum septentrionale его остатки распознаются в верхних слоях залежи наряду с Filipendula ulmaria s.l. Нанорельеф бывает как выровненным, так и кочковатым с высотой кочек 30–80 см, с преобладанием ковров и малочисленными западинами, реже с преобладанием западин и без ковров. Первый ярус древостоя той же сомкнутости, что и в ельниках таволгово-дернистоосоковых типичной субассоциации, но выше; бонитет древостоя достигает II. Примесь Betula pubescens s.l. к Picea abies s.l. чуть более 2 единиц по составу яруса; примесь Pinus sylvestris нехарактерна. Второй ярус смешанный березово-еловый. Он сильно разрежен, как в восточном варианте ельников таволговодернистоосоковых. ПП яруса подроста и подлеска как в ельниках таволгово-дернистоосоковой субассоциации в целом. Еловый подрост рассеянный, малочисленный, но хорошей жизнеспособности. Основную роль в формировании подлеска играет Ribes nigrum, в меньшей степени Sorbus aucuparia s.l., Alnus incana, Rosa acicularis. В травяном ярусе выраженно доминирует Filipendula ulmaria s.l. Доминанты 2-го порядка – Aconitum septentrionale и (при стволах и на кочках) Oxalis acetosella, которую сопровождают Maianthemum bifolium и иные виды мезофильного лесного мелкотравья, а также Vaccinium vitis-idaea вместе с Rubus saxatilis и Lathyrus vernus. Из сопутствующих видов гигро-мезофильного высокотравья наиболее характерны Crepis paludosa, Cirsium oleraceum, C. heterophyllum, в меньшей степени Trollius europaeus. На участках ковров постоянны также Geum rivale, Geranium sylvaticum s.l., Viola epipsila и Ranunculus subborealis, в западинах – Caltha palustris. Постоянна, но малообильна и Carex cespitosa. В моховом ярусе согосподству-ют Sphagnum warnstorfii и Rhytidiadelphus triquetrus; субдоминантами выступают Hylocomium splendens, Pleurozium schreberi и Plagiomnium ellipticum.
Среди собственных детерминантов ассоциации преобладают мезоэвтрофные гигромезофиты (Aconitum septentrionale, Ranunculus repens) и мезогигрофиты (Ribes nigrum, Chrysosplenium alternifolium, Brachythecium rivulare), отчасти также мезофиты (Vicia sepium, Rhodobryum roseum). Из мезотрофов к группе присоединяются Equisetum scirpoides и Brachythecium mildeanum (табл. 1). Экологическая характеристика детерминантов, общих с ельниками тавол- гово-дернистоосоковыми, приведена выше. Одновременно выражена негативная дифференциация от последних по отсутствию видов из группы Pinus sylvestris–Sphagnum centrale (табл. 1).
Данную ассоциацию также можно разбить на два варианта, замещающих друг друга в долготном направлении.
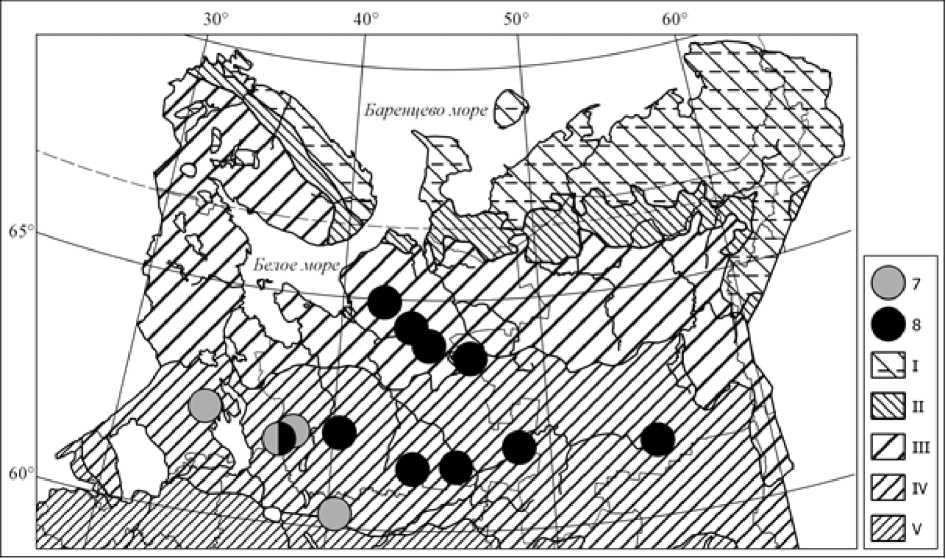
Рис. 2. Распространение ельников аконитово-таволговых сфагновых западного (типичного) (7) и восточного (пойменно-топяного) (8) вариантов в Европейской России и на Урале (картооснова как на рис. 1)
Var. typica – западный (типичный) вариант (табл. 1, рис. 2: № 7). Чаще всего встречается по окраинам жестководных болот и на выходах ключей при основании озовых гряд, реже по ложбинам стока и вдоль ручьев. Второй ярус древостоя, а также подрост и подлесок такой же высоты, как и в большинстве других рассматриваемых синтаксонов. Сравнительно с восточным вариантом, в травяном ярусе повышены ПП Equisetum sylvaticum, встречаемость Athyrium filix-femina и Solidago virgaurea . Дифференциация от восточного варианта негативная (табл. 1).
Основная часть описаний синтаксона приходится на среднюю тайгу Пудожской Карелии и Каргополья (НП «Кенозерский»), где леса произрастают в условиях близкого залегания известняков либо разгрузки минерализованных грунтовых вод. Отдельные описания, однако, сделаны и западнее, в сельговых ландшафтах заповедника «Кивач», также при наличии жестководного подтока. В южной тайге сходные ельники («травяноболотный» и «крупнотравно-приручейниковый») отмечены в поймах рек Вожега и Сить в районе залегания карбонатной морены в Вологодской обл. [2].
Родственные (но не строго аналогичные) сообщества ельников с Filipendula ulmaria, Crepis paludosa, Cirsium heterophyllum, Geum rivale представлены также в Южной Финляндии (true thin-peated herb-rich forests [40]), где тоже есть участки перемещенной карбонатной морены.
Var. Calliergon richardsonii – восточный (пойменно-топяной) вариант (табл. 1, рис. 2: № 8). Сообщества понижений в поймах рек и ручьев, относящихся к бассейну Северной Двины и ее притоков. Реже встречаются по окраинам жестководных болот. Второй ярус древостоя существенно выше, чем в лесах западного варианта и всех остальных рассматриваемых синтаксонов; возрастает и средняя высота подроста. Возможно, это следствие старовозрастности и малой нарушенности лесов Пинежско-Мезенского массива, в пределах которого выполнена значительная часть описаний. В травяном ярусе сравнительно с западным вариантом возрастает встречаемость ряда мезогигрофитов и западинных гигрофитов ( Comarum palustre, Epilobium palustre, Chrysosplenium alternifolium ). В моховом ярусе у Pseudobryum cinclidioides также наблюдается некоторый рост покрытия. Более высокая встречаемость характерна для ряда сопутствующих видов мезофитов ( Equisetum pratense, Geranium sylvaticum s.l., Ptilium crista-castrensis ) и гигромезофитов ( Rhizomnium pseudopunctatum ).
По сравнению с типичным (болотным) вариантом в пойменно-топяном существенно возрастает видовое богатство: 70–110 видов сосудистых растений и мхов на описание, тогда как в типичном варианте – 50–80.
Детерминанты варианта – мезоэвтрофные за-падинные мезогигрофиты и гигрофиты ( Cardamine amara, Galium uliginosum, Bryum weigelii, Calliergon richardsonii, Warnstorfia exannulata ). К ним тяготеют также Rubus humulifolius (мезотрофный гигромезофит микроповышений) и эпифитные мхи-мезофиты ( Pohlia nutans s.l., Brachythecium salebrosum, Sciuro-hypnum curtum s.l., S. reflexum ; возможно, вхождение эпифитов в эту группу случайно). С восточным вариантом типичных ельников таволгово-дернистоосоковых синтаксон сближают виды из групп Salix caprea–Bistorta major, Lonicera pal-lasii–Veratrum lobelianum и Lathyrus vernus– Cirsium oleraceum, а также Galium boreale , с ви-люйскоосоковыми – мхи из группы Bryum pseudotriquetrum–Rhytidiadelphus subpinnatus. Все группы, кроме Salix caprea–Bistorta major и Lathyrus vernus–Cirsium oleraceum, отличают восточный вариант от западного (табл. 1).
Ельники восточного варианта описаны в северной и средней тайге Архангельской обл. В первой из подзон они известны на Средней Пи-неге, в Келдинско-Полтинском и Пинежско-Мезенском ландшафтах, где произрастают не только на карбонатных породах, но и на гипсах [24]. Большое число описаний выполнено также в пойме р. Нюхчи на Верхней Пинеге. В средней тайге сообщества повсеместны в понижениях высокой поймы в центральной части, на юге и востоке области. В Республике Коми они отмечены в верховьях Вычегды [6], но пока не выявлены ни восточнее (в бассейне Печоры), ни южнее (в южной тайге).
В тех же регионах, что и ельники аконитовотаволговые сфагновые, в поймах малых рек и лесных ручьев обычны сходные ельники без участия Sphagnum warnstorfii и других видов сфагновых мхов в сложении мохового яруса [42]. Детерминантами этих сообществ выступают Cirsium oleraceum, Chrysosplenium alternifolium , Circaea alpina, Cinna latifolia , Brachythecium rivulare, Mni-um stellare, Conocephalum conicum . Не выражены ни торфяная залежь, ни даже связная лесная подстилка; моховой ковер из Brachythecium rivulare и Plagiomnium spp. формируется на тонком слое опада, покрывающем суглинок. Бонитет древостоя часто достигает I. Ареал подобных ельников шире, чем у аконитово-таволговых сфагновых. Они встречаются по всей таежной зоне Двино-Печорского региона, включая и полосу крайнесеверной тайги, а также на севере Кировской [28] и
(возможно) Костромской областей. Ранее данные сообщества были описаны из НП «Кенозерский» под именем Filipendulo-P. aconitetosum var. Circaea alpina. Аконитово-таволговые сфагновые ельники при этом рассматривались в качестве варианта Sphagnum warnstorfii той же субассоциации [14]. Безусловно, многое говорит о родстве этих двух синтаксонов. Однако при объединении их в одну ассоциацию возникает парадокс: два синтаксона, подчиненных одной и той же ассоциации, оказываются принадлежащими к разным группам ассоциаций еловых лесов – травяно-сфагновой и травяной. Аргументом против объединения служит и наличие торфяной залежи в ельниках со сфагновым покровом. Тем не менее, можно констатировать подлинно пограничный характер ассоциации ельников аконитово-таволговых сфагновых, находящейся на рубеже между двумя группами ассоциаций в пространстве факторов среды.
ЗАКЛЮЧЕНИЕ
Всего в Европейской России и на Урале нами описаны 5 субассоциаций и 2 варианта мезоэв-трофных ельников таволгово-дернистоосоковых сфагновых и 2 варианта ельников аконитовотаволговых сфагновых, итого 8 синтаксонов. Из них лишь ельники таволгово-вилюйскоосоковые являются сугубо северотаежным синтаксоном, остальные семь встречаются преимущественно в средней тайге. Амплитуда зональной приуроченности краевых местонахождений синтаксонов при этом может быть различной. Если ельники таволгово-молиниевые произрастают исключительно в средней тайге, то аконитово-таволговые восточного варианта – также в южной полосе северотаежной подзоны, тогда как западного – в средней и южной тайге, равно как и ельники дернистоосоково-черничные. Ельники таволгово-плевельноосоковые встречаются от южной тайги до южной полосы северной тайги, а оба варианта ельников таволгово-дернистоосоковых типичной субассоциации – до крайнесеверной тайги, в указанных пределах демонстрируя интразональность. Сказанное подтверждают и амплитуды толерантности ельников соответствующих синтаксонов по отношению к фактору теп-лообеспеченности (табл. 2).
С точки зрения долготного простирания ареала наиболее широко распространены ельники дернистоосоково-черничные с восточноевропейским ареалом от Карелии до Предуралья. Ельники таволгово-плевельноосоковые демонстрируют не столь обширный карельско-западносеверодвинский ареал. Для двух синтаксонов – ельников аконитово-таволговых и таволговодернистоосоковых типичной субассоциации – характерны замещающие долготные варианты, представленные соответственно в карельско- западносеверодвинском и северодвинско-печорско-уральском секторах России, с областью перекрытия ареалов в левобережной части бассейна Северной Двины. Ельникам таволгово-молиниевым и таволгово-вилюйскоосоковым свойственны локальные долготные ареалы, соответственно карельский и пинежско-кулойский.
Меридиональное распространение синтаксо-нов со сравнительно узким долготным ареалом может быть обусловлено не только градиентом континентальности, как в случае ельников тавол-гово-молиниевых (табл. 2), но и географическим распространением почвообразующих пород, от которых зависит требуемое минеральное богатство почвы. В частности, ельники таволгово-вилюйскоосоковые приурочены преимущественно к сульфатным почвообразующим породам пермского периода, чьи залежи распространены субмеридионально.
Пойменно-топяные ельники аконитовотаволговые восточного варианта могут встречаться как на силикатных, так и на карбонатных и сульфатных породах, проявляя нейтральность по отношению к данному фактору. Сообщества всех остальных синтаксонов произрастают как на силикатных, так и на карбонатных породах с равной частотой либо (как ельники аконитовотаволговые западного варианта), предпочитая более плодородные карбонатные, но не отмечены в области залегания сульфатов.
Авторы признательны к.б.н. Е.Ю. Чураковой (ФИЦКИА РАН) и к.б.н. В.Н. Мамонтову (НП «Вод-лозерский») за участие в совместных полевых исследованиях, к.б.н. М.А. Бойчук и к.б.н. А.И. Максимову (ИБ КарНЦ РАН) за определение критических сборов мхов, проф. Я. Паалю (J. Paal, Univ. Tartu, Estonia) за предоставление неопубликованных описаний. Работа выполнена в рамках действующих государственных заданий БИН РАН по теме АААА-А19-119030690058-2 «Разнообразие, динамика и принципы организации растительных сообществ Европейской России» и ИБ КарНЦ РАН по теме АААА-А19-119062590056-0 «Оценка разнообразия болотных и луговых экосистем, их динамика и история формирования на Европейском Севере».
Список литературы Мезоэвтрофные ельники таволгово-дернистоосоковые и аконитово-таволговые сфагновые европейской России и Урала
- Андреев В.Н. Лесная растительность южного Тимана // Тр. Полярной комиссии СОПС АН СССР. 1935. Т. 24. С. 7-64.
- Гаврилов К.А., Карпов В.Г. Главнейшие типы леса и почвы Вологодской области в районе распространения карбонатной морены // Тр. ин-та леса и древесины. 1962. Т. 52. С. 5-119.
- Громцев А.Н. Основы ландшафтной экологии европейских таежных лесов России. Петрозаводск: КарНЦ РАН, 2008. 238 с.
- Забоева И.А. Почвы и земельные ресурсы Коми АССР. Сыктывкар: Коми кн. изд-во, 1975. 344 с.
- Исаченко Т.И., Лавренко Е.М. Ботанико-географическое районирование // Растительность европейской части СССР. Л.: Наука, 1980. С. 10-22.
- Колесников Б.П. Лесная растительность юго-восточной части бассейна Вычегды. Л.: Наука, 1985. 216 с.
- Коротков К.О. Леса Валдая. М.: Наука, 1991. 160 с.
- Коровкин А.А. Геоботанический очерк Хибинского массива // Изв. Гос. Геогр. о-ва. 1934. Т. 66, № 6. С. 787-825.
- Кутенков С.А., Кузнецов О.Л. Разнообразие и динамика заболоченных и болотных лесов Европейского Севера России // Разнообразие и динамика лесных экосистем России. Кн. 2. М.: Товарищество научных изданий КМК, 2013. С. 152-206.
- Кучеров И.Б. Еловые леса верховий р. Кулой // Современные проблемы притундровых лесов: Материалы Всерос. конф. с междунар. участием (Архангельск, 4-9 сентября 2012 г.). Архангельск: САФУ, 2012. С. 162-172.
- Кучеров И.Б. Ценотическое и экологическое разнообразие светлохвойных лесов средней и северной тайги Европейской России. СПб.: Марафон, 2019. 568 с.
- Кучеров И.Б., Кутенков С.А. Мезотрофные кустарничковые сфагновые и сфагново-зеленомошные ельники Европейской России и Урала // Бот. журн. 2019. Т. 104, № 6. С. 859-887.
- Кучеров И.Б., Кутенков С.А. Мезотрофные ельники хвощовые, крупнопапоротниково- и хмеле-нично-хвощовые сфагновые средней и южной тайги Европейской России и Урала // Бот. журн. 2020. Т. 105, № 5. С. 438-457.
- Кучеров И.Б., Разумовская А.В., Чуракова Е.Ю. Еловые леса национального парка «Кенозер-ский» (Архангельская область) // Бот. журн. 2010. Т. 95, № 9. С. 1268-1301.
- Миняев Н.А. История развития флоры северо-запада европейской части СССР с конца плейстоцена: Автореф. дис. ... докт. биол. наук. Л.: ЛГУ, 1966. 38 с.
- Морозова Р.М. Лесные почвы Карелии. Л.: Наука, 1991. 184 с.
- Ниценко А.А. Еловые леса Ленинградской области // Вестн. Ленингр. ун-та. Сер. 3: Биол. 1960. Т. 2, № 21. С. 5-16.
- Пааль Я.Л. Первичные геоботанические данные: схемы расположения пробных площадей, обилие видов растений по отдельным ярусам, мощность генетических горизонтов почвенного профиля. Кивач, 1978. 223 с. (Рукопись в архиве заповедника «Ки-вач»).
- Полуяхтов К.К. Растительность Свердловской области // Учен. записки Смоленск. пед. ин-та. 1958. Вып. 6. С. 3-317.
- Попов П.П. Ель европейская и сибирская: структура, интерградация и дифференциация популя-ционных систем. Новосибирск: Наука, 2005. 231 с.
- Потемкин А.Д., Софронова Е.В. Печеночники и антоцеротовые России. СПб.; Якутск: Бостон-Спектр, 2009. Т. 1. 368 с.
- Пьявченко Н.И. Лесное болотоведение. М.: Наука, 1963. 192 с.
- Рысин Л.П., Савельева Л.И. Еловые леса России. М.: Наука, 2002. 335 с.
- Сабуров Д.Н. Леса Пинеги. Л.: Наука, 1972. 173 с.
- Самбук Ф.В. Печорские леса // Тр. Бот. музея АН СССР. 1932. Т. 24. С. 63-245.
- Сафронова И.Н., Юрковская Т.К. 2015. Зональные закономерности растительного покрова равнин Европейской России и их отображение на карте // Бот. журн. Т. 100, № 11. С. 1121-1141.
- Скляров Г.А., Шарова А.С. Почвы лесов Европейского Севера. М.: Наука, 1970. 272 с.
- Смирнова А.Д. Типы еловых лесов крайнего севера Кировской области. Ч. II // Учен. записки Горьковск. ун-та. 1954. Т. 25. С. 191-226.
- Смирнова З.Н. Лесные ассоциации северозападной части Ленинградской области // Тр. Петергоф. естеств.-науч. ин-та. 1928. № 5. С. 119-258.
- Сочава В.Б. Ботанический очерк лесов Полярного Урала от р. Нельки до р. Хулги // Тр. Бот. музея АН СССР. 1927. Т. 21. С. 1-78.
- Список лихенофлоры России /Урбанавичюс Г.П., сост.; Андреев М.П., отв. ред./ СПб.: Наука, 2010. 194 с.
- Сукачев В.Н. Руководство к исследованию типов леса. 3-е изд. М.: Сельхозгиз, 1931. 328 с.
- Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. Лесные экосистемы северо-западных районов России: Типология, динамика, хозяйственные особенности. СПб.: СПбНИИЛХ, 2005. 382 с.
- Цвелев Н.Н. Род Береза - Betula L. // Флора Восточной Европы. М.; СПб.: Товарищество научных изданий КМК, 2004. Т. 11. С. 65-85.
- Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья-95, 1995. 991 с.
- Шиманюк А.П. Опыт изучения северных лесов. М.; Л.: Сельхозгиз, 1931. 104 с.
- Юдин Ю.П. 1948. Темнохвойные леса Коми АССР (геоботаническая характеристика): Дис. ... докт. биол. наук. Сыктывкар: Коми филиал АН СССР. 323 с. (Рукопись в библиотеке БИН РАН.)
- Brandt A. Hiisjarven luonnonpuiston kasvil-lisuudesta // Silva Fennica. 1933. V. 32. 108 s.
- Chytry M., Zeleny D., Návratilová J., Sádlo J. Boreokontinentální jehlicnaté lesy // Vegetace Ceské re-publiky. T. 4: Lesní a krovinná vegetace. Praha: Academia, 2013. S. 380-432.
- Eurola S., Hicks S., Kaakinen E. Key to Finnish mire types // European Mires. L.: Acad. Press, 1984. 117 p.
- Ignatov M.S., Afonina O.M., Ignatova E.A. Check-list of mosses of East Europe and North Asia // Arctoa: Бриологический журнал. 2006. V. 15. P. 1-130.
- Kucherov I.B. Chorological pattern in the Aco-nito-Piceetum in middle and northern taiga of European Russia // Abhandl. Westfälisch. Museum für Naturkunde. Hf 3/4: Aspekte der Geobotanik - From Local to Global. 2008. S. 313-324.
- NASA surface meteorology and solar energy. 2006. http://eosweЫarc.nasa.gоv (accessed on 15.05.2006).
- Oberdorfer E. Süddeutsche Pflanzengesellschaften. Teil 4: Wälder und Gebüsche. 2 Aufl. Jena: Gustav Fischer, 1992. A. Textband. 282 S. - B. Tabellenband. 580 S.
- Paal J. Eesti taimkatte kasvukohatüüpide klassifi-katsioon. Tartu: Tartu Ülikool bot. ökol. inst., 1999. 202 lk.
- Páhlsson L. (ed.) Vegetationstyper i Norden. Kö-penhavn: Nordiska Ministerrädet, 1994. 627 s.
- Tukhanen S. Climatic parameters and indices in plant geography // Acta Phytogeogr. Suec. 1980. V. 67. 105 p.
- Willner W., Grabherr G. (eds.) Die Wälder und Gebüsche Österreichs. Ein Bestimmungswerk mit Tabellen. Im 2 Bände. München: Spektrum, 2007. 608 S.
