Микробиоценоз радужной форели в садковых хозяйствах Северной Карелии

Автор: Паршуков Алексей Николаевич, Сидорова Наталья Анатольевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (145) т.1, 2014 года.
Бесплатный доступ
Особенности состава микрофлоры характеризуют физиологический статус разводимых рыб и условия содержания их в садках. При этом у рыб, находящихся в тесном контакте с водой, бактериальная обсемененность внешних и внутренних органов будет зависеть от количества и качества микроорганизмов вокруг. Массовое развитие водных сапрофитных бактерий является стрессовым фактором, на борьбу с которым у рыб затрачиваются дополнительные ресурсы. На фоне неблагоприятных условий обитания снижаются защитные силы организма и появляются различные инфекционные заболевания. Впервые изучены закономерности качественного и количественного распределения микроорганизмов в микробиоценозе радужной форели и водной среды в отдельных форелевых хозяйствах Карелии. Установлено доминирование условно-патогенных бактерий рода Pseudomonas. Анализ состава микробиоценоза рыб демонстрирует высокую значимость полученных данных и возможность их применения в качестве индикаторных тестов для выявления изменений динамического равновесия бактериальной флоры рыб и естественных водных экосистем.
Аквакультура, радужная форель, микробиоценоз
Короткий адрес: https://sciup.org/14750783
IDR: 14750783 | УДК: 579.26:597.552.512:639.31(470.22)
Microbiocenosis of cage farm rainbow trout in Northern Karelia
Unique features of the microflora structure characterize current physiological status of the fish farmed in cages and of their farming conditions. Whilst fish is in close contact with water, bacterial contamination of the fish external and internal organs depends on the quality and quantity of microorganisms around. The mass growth of aquatic saprophytic bacteria is a stress factor, and the fish spends its vital resources to fight it. Due to the adverse environmental conditions, the body resistance of the fish decreases and a variety of infectious diseases appear. For the first time, regularities of qualitative and quantitative distributions of microorganisms in the microbiocenosis of the rainbow trout and water pollution in some trout farms in Karelia are studied. It was established that the opportunistic bacteria of the genus Pseudomonas are dominant in the rainbow trout. Analysis of the fish microbiocenosis composition revealed how important are the obtained data. It also helped to substantiate their possible use as a screening test to detect changes in dynamic equilibrium of the fish bacterial flora and in natural aquatic ecosystems.
Текст научной статьи Микробиоценоз радужной форели в садковых хозяйствах Северной Карелии
В Карелии широкое распространение получило садковое рыбоводство – направление аквакультуры, где производится выращивание радужной форели [11]. Форелевые хозяйства служат своеобразным типом местообитания для аллохтонных микроорганизмов, поступающих с кормом, а также с разводимой рыбой и продуктами ее метаболизма [2], а загрязнение органического происхождения приводит к структурно-функциональным изменениям в составе местной микрофлоры [13].
Негативное влияние симбионтного взаимодействия микроорганизмов с рыбой способствует бурному развитию ассоциаций бактерий, повышающих свои патогенные свойства. Динамика симбионтов в таких условиях постоянно меняется и заложена в высоком адаптационном потенциале как рыбы, так и бактерии. Благодаря высокой приспособляемости к агрессивным факторам среды усиливается ферментная активность бактерий, в результате чего симбионтные отношения в системе меняются в сторону доминирующих групп.
Таким образом, бактериальные показатели приобретают важное индикаторное значение,
позволяя выявить различные источники и типы антропогенного воздействия [1], [9].
Эффективность методов микробиологического анализа при мониторинге водоемов рыбохозяйственного назначения доказана на примере исследований, проводимых в Мурманской области [4]. Учитывая слабую изученность микрофлоры радужной форели в садковых хозяйствах Карелии, подобные исследования даже по одному из таких водоемов представляют научный и практический интерес.
Цель работы – изучение микробиоценоза радужной форели из садковых хозяйств на севере Карелии, работающих в течение одного года. Для достижения указанной цели поставлены следующие задачи:
-
1. Определить таксономический состав микрофлоры радужной форели в садковых хозяйствах на севере Карелии с небольшим периодом работы.
-
2. Дать численную характеристику гетеротрофных бактерий в микрофлоре рыб.
-
3. Изучить возможность применения бактериальных показателей для оценки степени структурно-функциональных изменений в составе микрофлоры форели.
МАТЕРИАЛЫ И МЕТОДЫ
Отбор проб на рыбоводных предприятиях проведен в период с апреля по сентябрь 2008– 2009 годов согласно определенной нормативнотехнической документации, содержащей необходимые требования, нормативы качества и методы исследования [10]. С помощью бактериологических методов изучен микробиоценоз радужной форели ( Parasalmo mykiss irideus ), выращиваемой на форелевых хозяйствах Карелии в акватории озер Хедо и Муй (система реки Кемь, бассейн Белого моря).
Для выделения бактерий и их дальнейшего изучения у живой рыбы стерильно получены образцы кожи и жабр. Далее проведены посевы на плотные питательные среды общего (рыбопептонный агар, рыбо-пептонный бульон), специального (Эндо, Пешкова) и дифференциальнодиагностического (Кесслера – для обнаружения бактерий группы кишечной палочки) назначения для изучения морфологических, культуральных и биохимических свойств бактерий. Дополнительно использованы среды: висмут-сульфит агар – для обнаружения бактерий, образующих сероводород (сальмонеллы); Плоскирева и Левина – для выявления не разлагающих лактозу патогенных бактерий семейства Enterobacteriaceae ; Сабуро – для селективного выделения дрожжеподобных и плесневых грибов.
Для описания морфологических свойств бактерий мазки окрашены по Граму, а затем исследованы при помощи светового микроскопа с иммерсией. Подвижность бактерий фиксирована при фазоконтрастном микроскопировании и по росту на полужидкой (0,3–0,7 % агара) среде. Гемолитическая активность бактерий проверена по гемолизу на чашках Петри с 5 %-м кровяным агаром. Для дифференциации представителей рода Pseudomonas от бактерий сходных с ними родов определена оксидазная активность культуры, способность расщеплять глюкозу и тип дыхания микроорганизмов в среде Хью-Лейфсона (тест окисления-ферментации).
Для установления систематической принадлежности изучаемых микроорганизмов использован определитель [7].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В формировании микробиоценоза форели, разводимой в хозяйствах с периодом работы один год (озера Муй и Хедо), участвуют 5 групп бактерий, представленных 8 семействами и 5 родами. Диапазон изменений количественных показателей гетеротрофов варьирует в пределах от 101 до 103 КОЕ/мл. Выделенные ассоциации микроорганизмов состоят как из сапрофитных, так и из условно-патогенных бактерий.
В результате анализа данных по бактериальной обсемененности кожных покровов и жабр молоди радужной форели из озера Муй в 2008–
2009 годах достоверно выявлено (P < 0,05), что на коже максимальная численность бактерий (302 ± 113 КОЕ/мл) соответствует периоду май 2009 года (температура воды – 6,5 °С), минимальная (18 ± 4 КОЕ/мл) – декабрь 2008 года (1,4 °С). На жабрах максимальная численность бактерий (1533 ± 62 КОЕ/мл) соответствует периоду сентябрь 2008 года (11,7 °С), минимальная (47 ± 20 КОЕ/мл) – декабрь 2008 года (1,4 °С) (рис. 1).
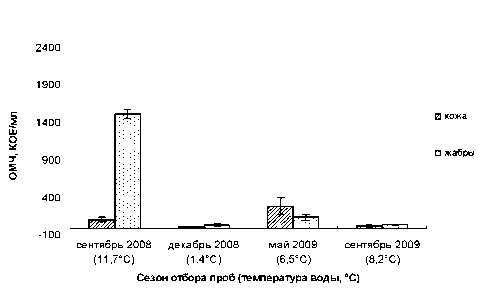
Рис. 1. Бактериальная обсемененность кожных покровов и жабр у форели (озеро Муй)
Полученные данные согласуются с общим представлением о том, что температурный фактор определяет интенсивность всех микробиологических процессов в воде, в том числе значение показателей численности [12], где невысокая бактериальная обсемененность совпадает с наступившим в это время холодным сезоном года (декабрь). Весенний подъем численности микробного числа характеризует прогревание водоема до благоприятных температур. Для рыбных хозяйств приведенные данные могут означать следующее: нетипичное низкое обсеменение в летний и осенний периоды, когда развитие бактериопланктона достигает своих максимальных значений, следует рассматривать как сигнал неблагополучного влияния на водоем со стороны самого предприятия, например из-за применяющихся в лечении рыб антибактериальных препаратов широкого спектра действия и др. Так, к примеру, анализ проб воды из садка на одном из форелевых хозяйств в южной части Карелии в летний период позволил определить численность, не превышающую 101 КОЕ/мл.
В период наиболее высокой численности бактерий в воде (сентябрь 2008 года) кожа рыб контаминирована незначительно – 123 ± 30 КОЕ/мл, в то время как на жабрах отмечена максимально высокая бактериальная численность – 1533 ± 62 КОЕ/мл. В течение всего исследования жабры были более обсемененными, чем кожа (P < 0,05). В сентябре 2008 года на жабрах грамположитель-ная микрофлора (66,7 %) доминирует над грамот-рицательной (33,3 %). По литературным данным [17], микробное число на жабрах может варьировать в широких пределах с 6×102 до 2,2×106. Считаем, что в сентябре 2008 года у исследованной группы рыб была превышена численность бактерий 1,5×103 КОЕ/мл по сравнению с остальными сезонами. Причиной тому могло послужить внешнее повреждение жаберного эпителия в результате воздействия различных стрессовых факторов – загрязнение воды органическими веществами, скученность, низкое содержание кислорода, повышенное количество аммиака, сортировка рыбы [8]. Известно, что бактериальная обсемененность кожных покровов и жабр, а также внутренних органов тесным образом связана с условиями содержания рыб в садках. При неудовлетворительном состоянии воды из-за применяемых мер по интенсификации производства повышается уровень бактериального загрязнения. Одна из причин – высокая плотность посадки рыб, которая способствует поступлению в водоем большого количества продуктов их метаболизма (органическое вещество), а несъеден-ные остатки корма только дополняют картину [1], [2], [6]. К примеру, из литературы известно, что на одном из рыбоводных хозяйств у карпа отмечалась значительная обсемененность покровов и органов, вызванная изначальным пребыванием посадочного материала в условиях предприятия с проводимыми на нем интенсификационными мероприятиями [5]. Не исключена также возможность инвазии тканей жаберного аппарата. К примеру, по результатам паразитологических и микробиологических исследований промысловых рыб дельты Волги [8] было установлено, что на фоне увеличения интенсивности инвазии наблюдалось повышение численности бактерий у рыб, однако качественные характеристики микробиоценоза оставались неизменными и соответствовали составу окружающей водной среды.
В ходе исследований установлено, что качественный состав микробиоценоза молоди форели из озера Муй представлен 7 семействами ( Pseudomonadaceae, Enterobacteriaceae, Mi-crococcaceae, Listeriaceae, Vibrionaceae, Bacil-laceae, Neisseriaceae ), 4 родами ( Pseudomonas, Micrococcus, Bacillus, Listeria ). В целом полученные результаты соответствуют литературным данным. По сообщениям ряда авторов, в составе микрофлоры рыб часто встречаются условнопатогенные микроорганизмы, среди которых представители семейств Pseudomonadaceae (род Pseudomonas ), Vibrionaceae (род Aeromo-nas ) и Enterobacteriaceae [1], [2], [3], [6], [14]. На коже и жабрах распространены бактерии родов Aeromonas, Pseudomonas, Citrobacter, Proteus, Enterobacter, Escherichia, Cytophaga, Flavobacte-rium, Micrococcus, Staphylococcus, Enterococcus [15], [16].
У исследованных рыб на коже доминируют представители семейства Pseudomonadaceae (род Pseudomonas – 79,2 %). Выделенные штаммы – прямые палочки, окрашиваются по Граму отрицательно, обладают подвижностью. Аэробы, метаболизм чисто дыхательного типа (тест O/F в среде Хью-Лейфсона). Оксидазо- и каталазоположительные, не декарбоксилируют лизин и орнитин. Вторыми по частоте встречаемости являются бактерии из семейства Enterobacteriaceae (8,3 %) – прямые палочки, грамотрицательные. Обладают и дыхательным, и бродильным типом метаболизма (среда Хью-Лейфсона). Оксидазоотрицательные. Остальные группы Vibrionaceae, Bacillaceae (род Bacillus) и Listeriaceae (род Listeria) выделяются с одинаковой частотой (4,2 %). На жабрах микробиоценоз в основном представлен бактериями из семейства Pseudomonadaceae (род Pseudomonas – 72,7 %), затем следуют представители семейства Enterobacteriaceae (9,1 %), а семейства Bacillaceae (род Bacillus), Micrococ-caceae (род Micrococcus), Listeriaceae (род Listeria) и Neisseriaceae составляют по 4,5 % соответственно.
Во все периоды отбора проб на молоди форели из озера Муй доминирующее положение занимали представители семейства Pseudomonadaceae (род Pseudomonas ).
Осенью 2008 года на долю псевдомонад приходится 50 %, а к концу года – 76,5 %. К весне 2009 года количество псевдомонад возрастает до 77,8 %, а к осени 2009 года они полностью (100 %) составляют микробиоценоз рыб, потеснив остальных представителей (рис. 2).

Рис. 2. Сезонная встречаемость бактерий на коже и жабрах форели (озеро Муй)
Для псевдомонад зарегистрирован максимальный коэффициент Симпсона (C), который составляет от 0,25 до 1. Сопутствующая псевдомонадам микрофлора меняется в сезон. В сентябре 2008 года представлены семейства Bacillaceae (род Bacillus – 33,3 %) и Micrococcaceae (род Micrococcus – 16,7 %) с коэффициентами Симпсона 0,11 и 0,028 соответственно. В декабре 2008 года встречаются представители семейства Entero-bacteriaceae (5,9 %, коэффициент 0,003), и только в этот период, когда покровы рыб сильно контаминированы грибами, выявлены представители семейств Neisseriaceae и Listeriaceae (род Listeria ) (5,9 % и 11,8 % с коэффициентами 0,003 и 0,014). Весной 2009 года впервые появляются бактерии из семейства Vibrionaceae (5,6 %, коэффициент 0,003).
В микробиоценозе кожи и жабр представители семейства Pseudomonadaceae (род Pseudomonas ) в течение года не только доминируют над остальными бактериями, но к сентябрю 2009 года вытесняют их полностью. Доминирование палочковидных форм бактерий у рыб выступает индикатором загрязненности водоема и усиленных процессов эвтрофикации. В начале исследований отмечен высокий удельный вес бактериальных штаммов с гемолитической активностью и подвижностью, что обусловливает их патогенность.
Исходя из этого мы считаем, что подобная видовая сукцессия служит сигналом неблагополучия в экосистеме. Несмотря на то, что псевдомонады относятся к представителям нормальной микрофлоры рыб [6], при определенных условиях, будучи условно-патогенными, вызывают эпизоотии, особенно когда занимают доминирующее положение. Как отмечают исследователи [3], к псевдомонадам восприимчивы большинство видов пресноводных и морских рыб. В условиях бесконтрольного применения антибиотиков на рыбоводных хозяйствах были зафиксированы случаи вспышек псевдомоноза. Особое значение приобрели бактерии Ps. fluorescens var. capsulata , которые за счет своей капсулы оказались более устойчивыми к применению антибактериальных препаратов, потеснив многие виды сапрофитных микроорганизмов. В результате псевдомонады прочно заняли доминирующее положение в водоеме, резко ухудшив эпизоотическую ситуацию.
Изучение бактериальной обсемененности кожных покровов и жабр молоди радужной форели из озера Хедо позволяет достоверно (P < 0,05) установить, что на коже максимальная численность бактерий (2396 ± 8 КОЕ/мл) соответствует периоду сентябрь 2008 года (температура воды – 11,7 °С), минимальная (39 ± 12 КОЕ/ мл) – сентябрь 2009 года (10 °С). На жабрах максимальная численность бактерий (2132 ± 31 КОЕ/ мл) соответствует периоду сентябрь 2008 года (11,7 °С), минимальная (58 ± 14 КОЕ/мл) – декабрь 2008 года (1,5 °С) (рис. 3).
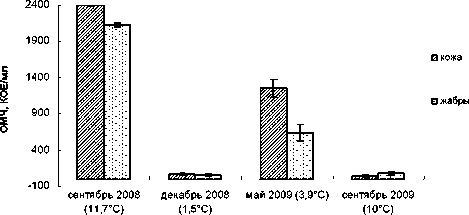
Сезон отбора проб (температура воды, °C)
Рис. 3. Бактериальная обсемененность кожных покровов и жабр у форели (озеро Хедо)
Температурный фактор здесь также сыграл ключевую роль в определении уровня обсеменен-ности – небольшая численность отмечена в под- ледный период, с последующим повышением за счет прогревания водоема. В период посадки молоди форели в садки (сентябрь 2008 года) на озере Хедо отмечена наивысшая численность бактерий (более 2 тыс. КОЕ/мл) на коже и жабрах по сравнению с другими сезонами (рис. 3). По нашему мнению, причиной такой разницы могло быть стрессированное состояние, в котором находилась рыба после недавней перевозки. Возможно, этим же объясняется дальнейшее более высокое микробное число в течение исследования у форели из озера Хедо, чем у форели из озера Муй. Низкая численность бактерий (менее 70 КОЕ/мл) выявлена в декабре 2008 года, что соответствует наиболее холодному сезону года (температура воды 1,5 °C). Однако в сентябре 2009 года при более высокой температуре воды (10 °C) установлена такая же низкая бактериальная обсемененность форели, как в декабре 2008 года, но, возможно, уже из-за антропогенного воздействия, поскольку санитарное качество воды к концу исследований становится все менее удовлетворительным. Самые низкие показатели численности зафиксированы в зимний период при температуре воды 1,4 °С и осенний 2009 года при температуре воды 10 °С.
По результатам исследований получены данные, характеризующие качественный состав микробиоценоза молоди форели из озера Хедо. Микрофлора рыб представлена 6 семействами ( Pseudomonadaceae, Enterobacteriaceae, Micrococ-caceae, Listeriaceae, Vibrionaceae, Saccharomyce-taceae ), 3 родами ( Pseudomonas, Staphylococcus, Listeria ).
В микробиоценозе кожи доминируют представители семейства Pseudomonadaceae (род Pseudomonas – 41,7 %) – грамотрицательные прямые палочки, подвижные. Аэробы, метаболизм чисто дыхательного типа (тест O/F в среде Хью-Лей-фсона). Оксидазо- и каталазоположительные, не декарбоксилируют лизин и орнитин. Обладают аргининдегидролазой. Далее идут семейства Vib-rionaceae (25 %) и Enterobacteriaceae (20,8 %). Доминирующим в микробиоценозе жабр является семейство Pseudomonadaceae (род Pseudomonas – 55,9 %). Представители семейства Vibrionaceae составляют 17,6 %, а семейства Enterobacteria-ceae – 11,8 %.
По результатам сезонной встречаемости бактерий на коже и жабрах молоди форели установлено разнообразие микробиоценоза во все периоды исследования, кроме начального (сентябрь 2008 года), где выделяются исключительно представители семейства Pseudomonadaceae (род Pseudomonas – 100 %) (рис. 4).
Значение показателя Симпсона у псевдомонад в сентябре 2008 года составляет 1 (для Ps. aeruginosa и Ps. alcaligenes С = 0,005; Ps. stutzeri С = 0,08; Ps. syringae С = 0,02; для остальных псевдомонад С = 0,184). В остальные сезоны (декабрь

Рис. 4. Сезонная встречаемость бактерий на коже и жабрах форели (озеро Хедо)
2008 – сентябрь 2009), наряду с псевдомонадами (декабрь 2008 год – 47,4 %; май 2009 года – 52,6 %; сентябрь 2009 года – 33,3 %), встречаются представители семейства Enterobacteriaceae (декабрь 2008 года – 21,1 %; май 2009 года – 10,5 %; сентябрь 2009 года – 20 %). В мае и сентябре 2009 года стабильно присутствуют бактерии семейства Vibrionaceae (удельный вес 36,8 и 33,3 % соответственно). Бактерии семейств Micrococ-caceae (род Staphylococcus ) и Listeriaceae (род Listeria ) обнаружены лишь в декабре 2008 года (коэффициенты Симпсона 0,011 и 0,044).
В начале исследований удельный вес бактериальных штаммов с гемолитической активностью и подвижностью отмечен как самый высокий, что обусловливает их патогенность, а в микробиоценозе форели установлено видовое однообразие, где доминируют только представители семейства Pseudomonadaceae (род Pseudomonas ), дополнительно указывая на негативную ситуацию в акватории. Похожая картина отмечена для микрофлоры форели из озера Муй, где также преобладают псевдомонады и отмечена их эпизоотическая и экологическая роль.
Таким образом, определена наиболее низкая бактериальная обсемененность молоди форели из водоема Муй, где посадка малька производи- лась на несколько месяцев раньше, чем у рыб из водоема Хедо. Качественный состав микробиоценоза более разнообразен у форели, выпущенной в садки раньше (озеро Муй), и отличается присутствием представителей семейств Vibrionaceae, Bacillaceae и Neisseriaceae. Схожими составы оказываются в семействах Pseudomonadaceae, Enterobacteriaceae, Micrococcaceae и Listeriaceae. В микрофлоре рыб доминируют представители семейства Pseudomonadaceae (род Pseudomonas), но больше их отмечено для форели из озера Муй (76,1 %), чем из озера Хедо (58,5 %).
ВЫВОДЫ
Анализ полученных материалов показал, что таксономический состав микрофлоры рыб представлен 8 семействами ( Pseudomonadaceae, Enterobacteriaceae, Vibrionaceae, Neisseriaceae, Micrococcaceae, Bacillaceae, Listeriaceae, Saccha-romycetaceae ), 5 родами бактерий ( Pseudomonas, Micrococcus, Staphylococcus, Bacillus, Listeria ).
На изученных форелевых фермах в микробиоценозе радужной форели доминируют условно-патогенные бактерии семейства Pseudomonadaceae рода Pseudomonas . При создании неблагоприятных условий для макроорганизма они могут повышать свою вирулентность и способны инфицировать стрессированную (ослабленную) рыбу.
Численность бактерий на коже и жабрах радужной форели из хозяйств с периодом работы около одного года (озеро Муй и Хедо) варьирует в пределах 101–103 КОЕ/мл при разных сезонных условиях среды и особенностях рыбоводных техник. Это отражает высокую значимость температурного фактора в развитии бактериопланктона и индикаторной роли значений численности микроорганизмов для определения качества водной среды и эффективности проводимых профилактических мероприятий.
MICROBIOCENOSIS OF CAGE FARM RAINBOW TROUT IN NORTHERN KARELIA
Список литературы Микробиоценоз радужной форели в садковых хозяйствах Северной Карелии
- Бычкова Л. И. Микробиоценоз радужной форели (Oncorhynchus mykiss Walbaum) и водной среды при садковом выращивании: Автореф. дисс.. канд. биол. наук. М., 2002. 27 с.
- Бычкова Л. И., Юхименко Л. Н., Можарова А. И. Микробиоценоз как индикатор экологического состояния водной среды и рыбы//Проблемы охраны здоровья рыб в аквакультуре: Тез. докл. научно-практической конф. М., 2000. С. 42-43.
- Головина Н. А., Стрелков Ю. А., Воронин В. Н., Головин П. П., Евдокимова Е. Б., Юхименко Л. Н. Ихтиопатология/Под ред. Н. А. Головиной, О. Н. Бауера. М.: Мир, 2003. 448 с.
- Карасева Т. А. Проблемы здоровья рыб в аквакультуре Севера России: на примере Кольского полуострова: Дисс.. канд. биол. наук. Мурманск, 2003. 168 с.
- Котлярчук М. Ю. Микробный пейзаж карпа (Cyprinus carpio L.) при выращивании в установке с замкнутым циклом водообеспечения: Автореф. дисс.. канд. биол. наук. Калининград, 2004. 23 с.
- Конев Н. В. Нормальная микрофлора рыб и ее роль в возникновении бактериальных заболеваний, вызванных стрессом: Научные тетради. СПб.: ГосНиОрХ, 1996. Вып. № 4. 46 с.
- Определитель бактерий Берджи: В 2 т./Под ред. Дж. Хоулта, Н. Крига, П. Снита, Дж. Стейли, С. Уильямса. М.: Мир, 1997. 368 с.
- Проскурина В. В., Лисицкая И. А. Паразиты и микрофлора промысловых рыб дельты реки Волга и их патогенное влияние на представителей ихтиофауны//Рыбное хозяйство. 2007. № 3. С. 69-71.
- Руководство по гидробиологическому мониторингу пресноводных экосистем/Под ред. В. А. Абакумова. СПб.: Гидрометеоиздат, 1992. 319 с.
- Руководство по проведению ветеринарного контроля над болезнями рыб и отбору проб для рыбоводных хозяйств северо-запада России/П. Коски. Оулу, 2006. 42 с.
- Рыжков Л. П., Нечаева Т. А., Евсеева Н. В. Садковое рыбоводство -проблемы здоровья рыб. Петрозаводск: Изд-во ПетрГУ, 2007. 120 с.
- Семушин А. В. Экология гетеротрофного бактериопланктонного сообщества прибрежных поверхностных вод Соловецкого архипелага: Автореф. дисс.. канд. биол. наук. Сыктывкар, 2003. 138 с.
- Широкова Л. С. Пространственно-временная структура бактериопланктона и его роль в самоочищении малых озер Архангельской области: Автореф. дисс.. канд. биол. наук. Архангельск, 2007. 189 с.
- Юхименко Л. Н., Койдан Г. С., Бычкова Л. И., Смирнов Л. П. Биологические свойства аэромонад и их роль в патологии рыб//Рыбное хозяйство. Сер. Болезни гидробионтов в аквакультуре. 2001. Вып. 1. С. 1-10.
- Buller N. B. Bacteria from fish and other aquatic animals: a practical identification manual. Cambridge, 2004. 361 p.
- Cahill M. M. Bacterial flora of fishes: a review//Microbial Ecology. 1990. Vol. 19. № 1. P. 21-41.
- Trust T. J. Bacteria associated with the gills of salmonid fishes in freshwater//Applied bacteriology. 1975. Vol. 38. P. 225-233.
