Микробиологическая трансформация углерода и азота в лесных почвах Средней Сибири

Автор: Сорокин Н.Д., Александров Д.Е.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Почвоведение
Статья в выпуске: 9, 2013 года.
Бесплатный доступ
Авторами статьи исследованы микробиологические факторы трансформации углерода и азота в лесных почвах Средней Сибири. Установлено, что общая продуктивность бактерий и микромицетов в исследуемых почвах составляет 1,2-3,8 т/га в верхнем слое 0-50 см, расчетный углерод микробной биомассы меняется в пределах 0,7-1,3 т/га, или 0,4-1,7 % от общего углерода. Энергия гумуса при переходе от почв южно-таежной подзоны к северным криогенным почвам уменьшается от 27,9х108 до 1,3х108 ккал/га. Микробиологическая трансформация азота и углерода может служить показателем лесорастительной способности (плодородия) почв.
Экология микроорганизмов, средняя сибирь, трансформация углерода, трансформация азота, плодородие почв
Короткий адрес: https://sciup.org/14083286
IDR: 14083286 | УДК: 630.548:630.114.61(571.51)
Carbon and nitrogen microbiological transformation in Middle Siberia forest soils
The microbiological factors of carbon and nitrogen transformation in Middle Siberia forest soils are researched by the authors of the article. It is established that the general efficiency of bacteria and micromycetes in the studied soils makes 1,2-3,8 t/hectare in the top layer of 0-50 cm, microbic biomass rated carbon changes within 0,7-1,3 t/hectare, or 0,4-1,7 % from the general carbon. Humus energy in transition from southern taiga subzone soils to northern cryogenic soils decreases from 27,9х108 to 1,3х108 kcal/hectare. The nitrogen and carbon microbiological transformation can serve as an indicator of the soil forest vegetation ability (fertility).
Текст научной статьи Микробиологическая трансформация углерода и азота в лесных почвах Средней Сибири
Исследование микробиологической трансформации углерода в лесных экосистемах Сибири имеет большое значение, так как дает возможность количественно оценить биогенную составляющую баланса углерода и его динамику в различных биогоризонтах (почва, подстилка, опад) под влиянием экологических и антропогенных факторов.
В то же время хорошо известно, что микробиологическая мобилизация углеродных соединений тесно связана с динамикой азота почвы. Сопряженные процессы микробиологической трансформации углерода и азота в лесных почвах севера и юга Средней Сибири имеют свои особенности, но однозначно в значительной степени определяют лесорастительную способность почв.
Цель исследований . Провести сравнительный анализ мобилизационной активности микробных комплексов лесных почв Средней Сибири при переходе от ее северной части к центральной и южнотаежной подзоне. Определить количественные микробиологические параметры трансформации углерода и азота.
Объекты и методы исследований . Объектами исследований явились микробные комплексы крио-земов гомогенных лиственничников Центральной Эвенкии, таежных осолоделых почв сосняков Иркутского Приангарья, серых лесных почв пихтарников Нижнего Приангарья и темно-серых лесных почв юга Красноярского края.
Для количественной оценки аккумуляции и эмиссии углерода в почвах лесных экосистем использовался регидратационный метод определения биомассы микроорганизмов [1] и газоаналитический метод СИД [7]. Скорость эмиссии СО 2 из почвы определяли по Очакову и на газовом хроматографе ЛХМ-80. Интенсивность деструкции клетчатки микроорганизмами изучали методом разложения стандартной целлюлозы на поверхности почвы и методом деструкции хлопчатобумажной ткани на глубине 0–20 см. Общую протеазную активность почв определяли по [3].
На основе полученных данных рассчитывали коэффициенты микробиологической активности Кма [5].
Результаты исследований и их обсуждение. Установлено, что в таежных почвах Сибири в период летней вегетации (июль) количество генераций микроорганизмов может достигать 7–9 независимо от зональной принадлежности почв (рис.).
Однако весной и осенью число генераций численности микроорганизмов в почвах северной и южной подзоны резко отличается. При этом биогенность 1 г почвы лесных территорий составляет от 2,3 млрд клеток в условиях Севера Сибири до 6,4 млрд в Приангарье и 8,9 млрд – в предгорьях Западного Саяна, что соответствует их реальной величине трофического коэффициента.
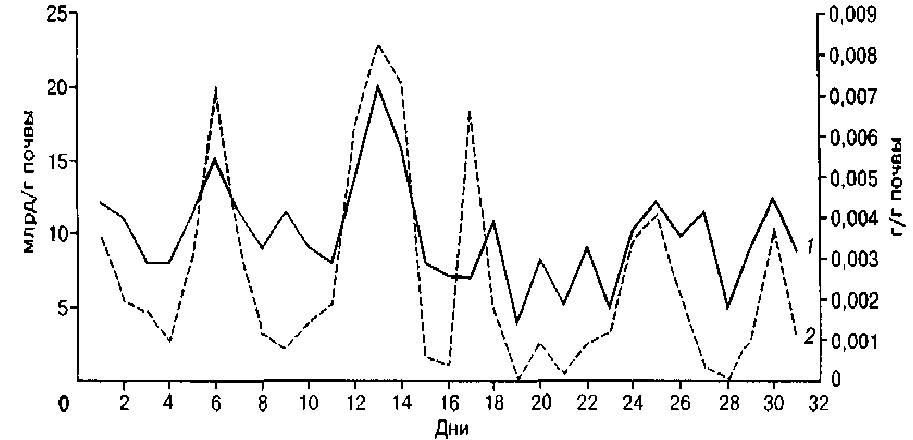
Динамика численности и биомассы бактерий в серой почве сосняка разнотравного (Нижнее Приангарье): 1 – численность бактерий, млрд/г почвы; 2 – биомасса бактерий, г/г почвы
Общая продуктивность бактерий и грибов верхнего гумусированного горизонта почв исследованных биогеоценозов достигает 1,4–4,0 мг/см3 (1,2–3,8 т/га в слое 0–50 см), или 0,3–1,5 % от количества гумуса. Известно, что содержание углерода в микробной клетке колеблется от 48 до 58 % [6]. Следовательно, расчетный углерод микробной биомассы исследованных почв меняется в пределах 0,65–2,0 мг/см3 (0,7–1,3 т/га в слое 0–50 см), или 0,4–1,7 % от общего углерода.
Следует отметить, что энергия, заключенная в микробной биомассе, расходуется клетками на конструктивные или/и на деструкционные процессы. Именно за счет гетеротрофной деструкции органического вещества лес поставляет в атмосферу двуокись углерода в количестве, сопоставимом и даже превышающем антропогенные поступления. Как отмечают И.П. Бабьева и Г.М. Зенова [1], в аэробных условиях грибы дают 2/3, а бактерии – 1/3 выделяющейся различными биогоризонтами углекислоты.
По нашим данным, продукция СО 2 1 га почв таежной зоны составляет 40–62 кг за 24 ч (табл.). Коэффициент окислительной активности, определяемый по соотношению дыхание/микробная биомасса, для исследованных почв равен 3–4. Это означает, что ежегодный приток углекислого газа в атмосферу из лесных почв исследуемых регионов за счет деятельности микробиоты колеблется от 4,8 до 5,5 т/га.
Мобилизационная активность микроорганизмов лесных почв Сибири
|
Район исследований, тип леса и почвы |
Интенсивность выделения СО 2 , кг/га за 1 ч |
Общая протеазная активность, % разрушения желатины за сезон |
Разложение клетчатки, % |
Коэффициент микро-биологической активности, К ма |
|
|
за сезон |
за год |
||||
|
Среднее Приангарье; сосняк брусничнозеленомошный; таежная осолоделая |
2,4±0,3 |
62 |
41 |
77 |
4,6 |
|
Нижнее Приангарье: пихтарник; серая суглинистая |
3,3±0,3 |
68 |
53 |
81 |
5,1 |
|
Западный Саян: пихтарник высокотравнопапоротниковый; горнотаежная бурая |
3,1±0,3 |
74 |
57 |
86 |
5,2 |
|
Центральная Эвенкия: лиственничник кустар-ничково-зеленомошный; крио-зем гомогенный |
0,8±0,1 |
37 |
18 |
37 |
1,7 |
|
Юг Красноярского края: сосняк мертвопокровный; темно-серая лесная |
4,8±0,05 |
72 |
53 |
84 |
5,8 |
Расчет скорости микробного дыхания (БД), биомассы микроорганизмов (МБ) и метаболического коэффициента (qCO 2 ) через субстратиндуцированное дыхание (СИД) свидетельствует о том, что этот метод дает превышение по биомассе микроорганизмов в 1,5 раза, а по интенсивности дыхания – в 2 раза. Таким образом, расчетные данные по эмиссии СО2 в исследуемых почвах должны быть увеличены примерно в 2–2,5 раза.
Следует отметить, что функциональная активность целлюлозоразлагающих микроорганизмов зависит от наличия в почве легкоподвижных азотных соединений, а скорость разрушения клетчатки пропорциональна общей протеазной активности почв. Зная величину общей протеазной и целлюлолитической активности и величину биомассы микроорганизмов, можно посчитать коэффициент микробиологической активности почвы (Кма), который представляет собой отношение суммарного количества разложившейся клетчатки и желатина в граммах к биомассе микроорганизмов за определенный отрезок времени [5]. Коэффициенты микробиологической активности закономерно возрастают при переходе от северных почв к почвам южной подзоны тайги Сибири (см. табл.).
Преимущественное использование микробоценозами и фитоценозами лесных сообществ аммонийных форм азота выражается в преобладающем развитии в исследуемых почвах аммонифицирующих микроорганизмов. Энергия размножения аммонификаторов выражается в аммонифицирующей способности почв. Накопление поглощенного аммония наиболее активно происходит в серых лесных и дерново-подзолистых почвах приенисейской части Сибири и наименее активно в супесчаной почве Среднего Приангарья. В первом случае накапливается до 17–20 мг NH 4 на 100 г почвы, во втором – 3,5 мг.
Накопление нитратных форм азота не имеет прямой зависимости от числа нитрифицирующих бактерий, но находится в большей связи с общей численностью микроорганизмов, участвующих в метаболизме азота. Нитрификаторы в исследуемых почвах развиты слабо вследствие неблагоприятных для них почвенно-экологических условий. Но согласно результатам Докстадера и Александера [8], нитрификация может осуществляться гетеротрофными организмами. В частности, гетеротрофные бактерии, актиномицеты и грибы продуцируют нитраты из различных восстановленных форм азота. Отсутствие соответствия между чис- лом нитрификаторов и содержанием нитратов в почве может являться именно этим фактом, и кроме того, другой причиной является поглощение нитратов высшими растениями и денитрификация.
Исследования показывают, что процессы трансформации легкогидролизуемых форм органики наиболее полно протекают в серых лесных почвах Нижнего Приангарья. Здесь при определенных экологических условиях в летний период может происходить биологическое окисление солей аммония в нитратную форму. В остальных почвах процесс трансформации азота заканчивается на стадии образования аммония. При этом возможны потери азота в газообразной форме за счет активного развития денитрификаторов или за счет иммобилизации его другими группами микроорганизмов, которые после отмирания и лизиса возвращают поглощенный азот в почву в более сложной органической форме.
Обобщая полученные данные по трансформации азота и углерода в лесных почвах Средней Сибири, мы имеем возможность провести оценку плодородия почв региона, поскольку, как было показано, азот является в данном случае основным компонентом органического вещества, лимитирующим плодородие.
В.А. Ковда [2] и И.И. Свентицкий [4] предлагают оценивать потенциальное плодородие почв по одному комплексному показателю – количеству энергии, заключенной в органическом веществе почв, поскольку и почвообразование в целом, и гумусообразование являются энергетическими процессами.
Если принять запасы гумуса и энергии за уровень потенциального плодородия, то исследуемые почвы можно расположить в следующий нисходящий ряд: дерново-перегнойные суглинистые – серые оподзо-ленные – дерново-подзолистые суглинистые – перегнойно-поверхностно-глеевые осолоделые – серые осолоделые – таежные осолоделые красно-бурые – дерново-карбонатные – дерново-подзолистые супесчаные. Энергия гумуса в этом ряду убывает от 27,9х108 до 1,3х108ккал/га. По запасам энергии в микробной массе такой закономерности не наблюдается. Самые низкие и довольно близкие между собой величины отмечены в дерново-перегнойных суглинистых почвах и дерново-подзолистых супесчаных – 1,2х106 и 1,3х106ккал/г соответственно. Причем доля микробной массы в энергии органических соединений в первом случае самая низкая, а во втором самая высокая среди исследованных почв.
Заключение . При переходе от криогенных почв севера Сибири (Центральная Эвенкия) к длительно сезоннопромерзающим почвам центральной и южной части Красноярского края резко возрастает продукция микробной массы (от 1,2 до 3,8 т/га) и дыхательная активность микробиоты. Сопряженно в 3–5 раз увеличивается аммонифицирующая способность микроорганизмов и деструкция целлюлозы. Показателем мобилизационной активности гетеротрофных микроорганизмов является коэффициент микробиологической активности – Kма, который закономерно возрастает в почвах юга Красноярского края по сравнению с северными почвами от 1,7 до 5,8.
По всем параметрам микробиологической трансформации углерода и азота (сбалансированности процессов деструкции и синтеза клетчатки, накоплению микробной биомассы, эмиссии СО 2 , процессам аммонификации, нитрификации и др.), определяющим почвенное плодородие, лучшие условия создаются в лесных экосистемах юга Красноярского края и Нижнего Приангарья. Самой низкой лесорастительной способностью обладают почвы северной и восточной части Средней Сибири (Центральная Эвенкия и Иркутское Приангарье).
