Микробиологические индикаторы раннеголоценовой притеррасной почвы долины реки Мзымта черноморского побережья Кавказа

Автор: Рогожина Е.В., Моисеева Е.В., Худокормов А.А., Захарихина Л.В.
Журнал: Бюллетень Почвенного института им. В.В. Докучаева @byulleten-esoil
Рубрика: Статьи
Статья в выпуске: 120, 2024 года.
Бесплатный доступ
На примере изучения раннеголоценовой палеоаллювиальной почвы (стоянка древнего человека в гроте Ахцу, надпойменная терраса среднего течения реки Мзымта Сочинского Причерноморья) показана эффективность применения комплекса микробиологических методов, раскрывающих условия формирования древнего палеообьекта и генезис геоморфологии речной долины. Исследование морфологии и мезоморфологии палеопочвы позволило сделать предположение о возможном ее формировании в условиях существенно более гидроморфных в сравнении с современными. Вероятно, ранее, в начале голоцена, данный участок являлся не террасой, а притеррасной переувлажненной частью поймы реки. Микробиологические характеристики подтвердили данное предположение. Из выделенной почвенной ДНК прокариотных микроорганизмов (бактерий и архей) методом ДНК-метабаркодинга идентифицирован состав микробного сообщества. Это представители доминантного в составе микробиома (более 1% от общей ДНК) филума Planctomycetes классов Phycisphaerae и Planctomycetia и цианобактерии класса Chloroplast порядков Streptophyta, Stramenopiles и Pseudanabaenale. Микробиологическими индикаторами переувлажненных условий формирования палеопочв в раннеголоценовом периоде являлся вид архей (0.5% от общей ДНК) филума Euryarchaeota; класса Thermoplasmata; семейства Methanomassiliicoccaceae, способный к метаногенезу. В микробном комплексе современной фоновой почвы данный вид не обнаружен. Выявленные в составе прокариотного сообщества палеопочв индикаторные микроорганизмы могут потенциально обладать полезными прикладными свойствами. Отмечено отсутствие культивируемых форм аэробных и анаэробных целлюлозолитиков, что обусловлено, возможно, отсутствием субстрата (клетчатки).
Притеррасные палепочвы, микроорганизмы, биоиндикаторы, метагеном, днк-метабаркодинг, сочинское причерноморье
Короткий адрес: https://sciup.org/143183586
IDR: 143183586 | DOI: 10.19047/0136-1694-2024-120-84-106
Microbiological indicators of Early Holocene near-terrace soil in the valley of the Mzymta river on the Black Sea coast of the Caucasus
By the example of studying the Early Holocene paleoalluvial soil (ancient man's site in Akhtsu Grotto, an overflow terrace of the middle reaches of the Mzymta River, Sochi Black Sea region), the efficiency of using a set of microbiological methods revealing the conditions of formation of the ancient paleoobject and the genesis of the geomorphology of the river valley was shown. The study of morphology and mesomorphology of the palaeo-soil allowed to assume its possible formation under the conditions significantly more hydromorphic in comparison with the modern ones. It is probable that earlier, at the beginning of the Holocene, this area was not a terrace, but a near-terrace, over-watered part of the river floodplain. Microbiological characteristics confirmed this assumption. The composition of the microbial community was identified from the isolated soil DNA of prokaryotic microorganisms (bacteria and archaea) by DNA metabarcoding. Groups and species of microorganisms-indicators reflecting the specificity of genesis of the studied soils were identified. Microbiological indicators of overwatered conditions were representatives of the phylum Planctomycetes of Phycisphaerae and Planctomycetia classes dominant in the microbiome (more than 1% of total DNA) and cyanobacteria of Chloroplast class of Streptophyta, Stramenopiles and Pseudanabaenale orders, as well as archaea (0.5% of total DNA) of Euryarchaeota phylum; Thermoplasmata class; Methanomassiliicoccaceae family, capable of methanogenesis. This species was not found in the microbial complex of the modern background soil. Indicator microorganisms identified as part of the prokaryotic community of palaeo-soils may potentially have useful applicable properties.
Текст научной статьи Микробиологические индикаторы раннеголоценовой притеррасной почвы долины реки Мзымта черноморского побережья Кавказа
354002, Сочи, ул. Я. Фабрициуса, 2/28, *, e-mail:
2/28 Ya. Fabricius Str., Sochi 354002, Russian Federation, *, e-mail:
149 Stavropolskaya Str., Krasnodar 350040, Russian Federation, **, e-mail: , ***
Успехи палеомикробиологии имели большое значение для формирования представлений об эволюции климата и условий почвообразования на Земле. Погребенные микробные сообщества палеопочв не только жизнеспособны, но и отражают природные условия, существовавшие в древности (Frindte et al., 2020). Эволюция конкретного почвенного ареала определяет состояние микробных сообществ почвы в тот или иной исторический период, поэтому возникает локальная специфика в структурнофункциональной организации микробного сообщества (Дёмкина, 2020).
Древние почвы довольно часто бывают обнаружены на территории речных террас (Hsu et al., 2021; Kuznetsova et al., 2020; Sedov et al., 2022), в том числе заселенных в древности человеком (Плеханова., 2019; Ovchinnikov et al., 2022). Речные террасы являются древними образованиями, сформированными двумя природными процессами - периодическим затоплением полыми водами и отложением аллювия (наилка). При этом процессе происходит естественное орошение и влагозарядка почв. Аллювиальные почвы наследуют от отлагаемого аллювия гранулометрический, петрографический, биолитный состав. Органические вещества поступают за счет привносимого наилка, так как он богат органическим углеродом, соединениями калия и фосфора, а также содержит разнообразную микрофлору, которая формирует впоследствии экосистему почвы или входит в состав уже существующего сообщества (Добровольский, 2011). Формирование палеоаллювиальных почв Кавказа также происходило под влиянием гидрорежима рек. Согласно литературным данным, аллювиальные почвы долин горных рек целесообразно использовать для восстановления кратковременных (голоценовых) событий в циклах потепления-похолодания, так как из-за высокой скорости течения годовая мощность отложений в поймах горных рек невелика (Kovaleva, Stolpnikova, 2020). Однако древние микробные сообщества формируются не только под влиянием климата эпохи, но и обусловлены спецификой биотопа, в котором шел процесс почвообразования. Гидрогеологические процессы (изменение русла реки, образование речных террас) приводят к сукцессии экосистем в пойме реки. В результате древние почвы сохраняют в своем составе микроорганизмы-индикаторы ранее существующих экотопов (Hsu et al., 2021). Исследования палеопочв с целью выделения микроор- ганизмов-хроноиндикаторов и экомаркеров проводятся сегодня не только традиционно – посевом почвенной суспензии на питательные среды, но и с привлечением молекулярно-биологических и биоинформационных (выделение тотальных ДНК и РНК из почвы, количественная ПЦР с детекцией в реальном времени, высокопроизводительное секвенирование) методов. Результаты исследований отражают численность генов основных групп почвенных микроорганизмов, содержание функциональных генов и таксономическую структуру микробиомов (Chernov et al., 2018). В совокупности использование химико-аналитических и количественных микробиологических, а также молекулярно-генетических исследований позволяет изучать не только текущее состояние микробиома почвы, но и следы древнего сообщества, которое неактивно или слабо выражается на фоне культивируемого сообщества.
В публикациях последних лет палеопочвы рассматривают как банк древних микроорганизмов, обладающих хозяйственно ценными свойствами – антимикробной, противовирусной, рост-стимулирующей активностью, а также способностью к синтезу важных биополимеров, биологически активных веществ (ферментов, витаминов) или к гидролизу ксенобиотиков, углеводородов, полимеров (Manucharova et al., 2020). В этой связи представлял интерес анализ прикладных свойств видов-индикаторов, обнаруженных в микробиоме. В Сочинском Причерноморье подобные исследования проводятся впервые.
Таким образом, целью исследований являлось определение групп и видов микроорганизмов-индикаторов, способных раскрыть историю формирования раннеголоценовой палеопочвы, обнаруженной в долине реки Мзымта Черноморского побережья Кавказа в месте стоянки древнего человека, а также выполнение анализа возможных прикладных свойств в области биотехнологии, выявленных в палеообъекте видов микроорганизмов.
ОБЪЕКТЫ И МЕТОДЫ
Исследования выполнялись на археологическом объекте Сочинского Причерноморья, который был отнесен к памятникам каменного века и получил наименование “пещерная стоянка в гроте Ахцу”. Грот Ахцу расположен в 20 км от береговой линии моря
(43°56´N; 39°99´E) примерно в 70 м от уреза р. Мзымта, абсолютная высота его над уровнем моря составляет 133 м, высота над уровнем реки 12 м (рис. 1).
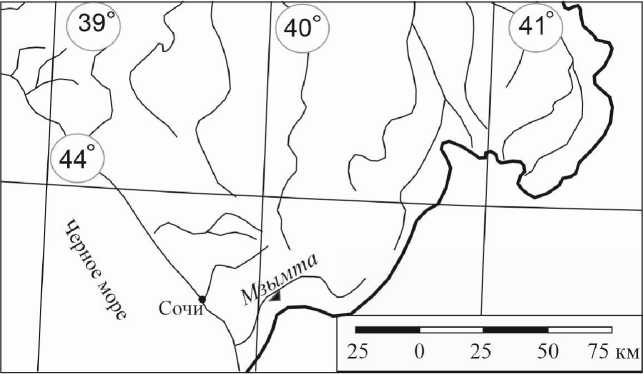
Рис. 1 . Обзорная схема расположения грота Ахцу.
Fig. 1 . Overview of the location of the Akhtsu grotto.
В открытом археологическом раскопе (заложенном при археологических раскопках в 2014–2015 гг.) была обнаружена палеопочва. Возраст слоя, в котором она располагалась, установленный археологами радиоуглеродным методом по кости древних животных, составляет 8.3 тыс. лет (Кулаков и др., 2017). Палеопочва залегала на глубине 12–30 см. Перекрыта она была мелко раздробленной породой пещерных отложений, имевших очень плотное, сцементированное сложение. Педогенное происхождение палеообъекта подтверждалось наличием отдельных фрагментов, хорошо сохранившихся ризолитов (древних корней) и высоким содержанием в нем органического углерода (2.3%).
Установленный почвенный палеогоризонт имел очень темную, фактически черную окраску, рыхлое аморфное сложение. Под лупой (×10) он представлял собой рыхло-сложенный темно бурый аморфный материал, скрепленный в комковатые отдельно- сти, напоминающий по сложению перегнойный горизонт H. Все морфологические и мезоморфологические особенности свидетельствовали в пользу гидроморфного происхождения палеопочвы. Кроме того, очень большая для погребенной почвы мощность горизонта (около 20 см) также указывала на то, что, вероятнее всего, в период раннего голоцена это была не минеральная почва, а, скорее, органогенная – либо аллювиальная с мощным перегнойным горизонтом H, сформированным прибрежной высокотравной растительностью, либо вовсе низинная торфяная почва. Поверхностные органоминеральные горизонты минеральных почв, имеющие, как правило, мощность при современном (дневном) залегании около 10–15 см, переходя в погребенное состояние приобретают небольшую мощность – не более 1–2 см (Zakharikhina et al., 2021).
В пользу органогенно-гидроморфного происхождения палеопочвы не свидетельствовало лишь ее очень сухое состояние. Она не имела мажущуюся консистенцию, характерную для перегнойных и низинных торфяных почв. Вероятно, ранее, в начале голоцена данный участок являлся не террасой, а притеррасной переувлажненной частью поймы реки. Такие низинные переувлажненные участки, часто образованные маломощными низинными торфяными почвами, характерны и в современности для долин рек региона. Позже, когда русло реки сместилось, пойма изменила геоморфологическое положение, перейдя в надпойменную террасу. Условия увлажнения здесь существенно изменились, стало суше, и гидроморфная органогенная почва значительно трансформировалась по степени увлажнения, сохранив при этом свое характерное сложение.
В целях сравнительного анализа современные аналоги палеопочвы (фон), имеющие схожие свойства и мезоморфологические характеристики, были опробованы в непосредственной близости от грота под высокотравной и разнотравной растительностью в расщелинах крупноглыбистой породы.
В сравнении с фоновыми почвами палеопочва грота имела пониженные содержания органического углерода (фон – 2.9%, палеопочва – 2.3%), подвижных форм калия (фон – 757 мг/кг, палеопочва – 209 мг/кг) и повышенные концентрации доступного фосфора (фон – 69 мг/кг, палеопочва – 219 мг/кг). Степень насыщенности основаниями в ней составляла 99%, что соответствовало показателям фона. Кислотность (рНKCl) палеопочвы и фона имела сходные значения и была равна 7.0 ед. Влажность палеопочвы была незначительно ниже (18%) по сравнению с фоном (24%) (Zakharikhina et al., 2021).
Морфология, свойства и местоположение в рельефе позволило классифицировать почву как аллювиальную темногумусовую (Шишов., 2008). В соответствии c World Reference Base for Soil Resources (World Reference Base…, 2014) и специальной па-леопочвенной классификацией Г. Мака (Mack et al., 1993), почва диагностировалась как Histosoils (Folic).
Качественное и количественное определение физиологических групп микроорганизмов проводили стандартными микробиологическими методиками с высевом на питательные среды (Практикум по микробиологии, 2005). Для количественной оценки содержания бактерий, мобилизующих органические источники азота, использовали мясо-пептонный агар (МПА), для учета олигокарбофилов – стандартную среду МПА, разбавленную в 10 раз, нитрификаторов – нитритный агар, свободноживущих азотфикси-рующих микроорганизмов – среду Эшби, анаэробных целлюлозо-литиков – жидкую среду Имшенецкого, аэробных – почвенную вытяжку с карбоксиметилцеллюлозой. Для культивирования фо-тотрофных серобактерий использовали накопительную среду Ван-Ниля. Для выявления серобактерий различных экологических групп применяли более универсальную среду Бейеринка.
Отбор почвенной пробы для проведения микробиологических посевов и молекулярно-генетического анализа был произведен с соблюдением условий стерильности. Образец погребенных почв отбирался с помощью cтерильных перчаток и ножа, затем помещался в стерильный полиэтиленовый пакет и пластиковую пробирку с герметичной крышкой, которую замораживали в морозильной камере при температуре -20 °C до отправки в специализированную лабораторию генетического анализа. Транспортировали образец в охлаждающей камере. ДНК-метабаркодинг состоял из следующих этапов: выделение из почвы суммарной ДНК, амплификация участков генов и идентификация ампликонов. Подго- товку геномных библиотек осуществляли согласно протоколу “Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System”. Амплификацию V3-V4 региона 16S рРНК проводили с использованием прокариотических праймеров: прямой – TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGG NGGCWGCAG; обратный – GTCTCGTGGGCTCGGAGATGTGT-ATAAGAGAC AGGACTACHVGGGTATCTAATCC, – с последующей индексацией ампликонов. Секвенирование выполнялось на платформе Illumina MiSeq (США) (режим парного чтения 300 пар нуклеотидов). При обработке данных секвенирования использовали алгоритм классификации операционных таксономических единиц (ОТЕ) с открытым референсом (Open-reference OTU), порог классификации – 97% сходства, что обычно соответствует виду. Визуализация данных проведена с помощью построения диаграмм потоков (Sankey diagram библиотека plotly, python).
Химические показатели были определены по общепринятым стандартным методикам (Практикум по агрохимии, 2001): pH KCl – потенциометрически (ионометр рН-121); органический углерод – по Тюрину в модификации Орлова и Гриндель; фосфор подвижный – по Олсену (прибор УСФ 01); калий обменный – по Масловой со спектрометрическим окончанием (прибор КВАНТ - АФА); содержание обменных форм Са2+ и Мg2+ в солевой вытяжке NaCl – трилонометрическим методом; гидролитическая кислотность – по Каппену; влажность – весовым методом.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Проведенные исследования аллювиальной темногумусовой почвы раннеголоценнового периода (грот Ахцу) показали, что численность в ней аммонифицирующих бактерий на порядок ниже, по сравнению с современной фоновой почвой (1.7*105 и 7.5*106 КОЕ/г соответственно), а содержание олиготрофов (1.7*106 и 3.6*106 КОЕ/г соответственно) и азотфиксаторов (5.2*106 и 3.6*106 КОЕ/г соответственно) соразмерно. Более активное развитие в палеопочве олиготрофов, по сравнению с аммонификаторами, является характерной особенностью погребенных почв (Демкин и др., 2013; Демкина, 2020) и может свидетельствовать о низком содержании в исследуемом образце органических соединений азота. Высокое (на уровне фона) содержание азотфик-саторов в микробных комплексах палеопочв может быть связано с недостатком доступных форм азота (Чернов, Железова, 2020). Численность нитрификаторов в палеопочве грота и фона различалась на 3 порядка и составила 6.0*103 и 1.4*106 КОЕ/г соответственно. На численность нитрификаторов, способных к разложению сложных азотсодержащих соединений (в том числе гумуса), возможно, в большей степени повлияли процессы диагенеза – трансформации палеопочв, происходящей при их погребении (Zakharikhina et al., 2021).
В результате исследования целлюлозолитической активности микроорганизмов палеопочвы разложения целлюлозы в анаэробных условиях не обнаружено. После 30-дневного культивирования в жидкой питательной среде заметного изменения плотности бумаги не наблюдалось, однако появлялся зеленый налет на бумаге и стенках пробирок, который связан с возможным заселением фототрофных анаэробных бактерий или водорослей. В аэробной среде наблюдали единичные колонии бактерий, разлагающих клетчатку, в то время как в фоновой почве их содержание соответствовало 1.1*105 КОЕ/г. Культивируемые формы аэробных и анаэробных целлюлозолитиков практически отсутствуют в палеопочве, что, возможно, связано с отсутствием клетчатки, гидролизованной в процессе диагенеза. В результате культивирования на среде Ван-Ниля были обнаружены обитатели природных вод – фототрофные серобактерии в виде единичных колоний на плотной среде. Столь незначительное их содержание в палеопочве, вероятно, являлось результатом контаминации в периоды паводков. Подтверждало данное предположение отсутствие серобактерий на среде Бейеринка.
По результатам секвенирования 16S рРНК прокариотное сообщество палеопочв грота и современных фоновых почв представлено преимущественно видами домена Bacteria (295 и 325 видов соответственно). Домен Archaea являлся минорным (3 и 2 вида соответственно). Своего рода индикатором палеопочвы грота Ахцу, в которой процессы минерализации органического вещества протекали в переувлажненных условиях, создающих анаэробные условия, выступает вид домена Archaea; филума Euryarchaeota ;
класса Thermoplasmata ; семейства Methanomassiliicoccaceae. Высокая доля содержания в микробоценозе палеопочвы данного вида (0.5% от общей ДНК) предполагает значительное участие Methanomassiliicocales в метаногенезе – анаэробном окислении метана . В микробном комплексе современной фоновой почвы данный вид не обнаружен. Согласно литературным данным, гены 16S рРНК, ассоциированные с Methanomassiliicoccales , были обнаружены в широком диапазоне бескислородных сред (Borrel et al., 2013), включая морскую среду обитания, рисовые поля и кишечные тракты термитов, млекопитающих, людей. Совсем недавно Зёллингер и др. (Sollinger et al., 2016) подтвердили, что Methanomassiliicoccales предпочитают анаэробную среду обитания, и разделили этот отряд на основе филогенетического и геномного анализа метаногенных последовательностей, выделенных из водно-болотных угодий и кишечного тракта животных, на 2 группы: “экологическую” и “желудочно-кишечного тракта”. Авторы также сообщили о более крупных геномах и бόльшем количестве генов, кодирующих антиоксидантные ферменты в “экологической” группе, что делает их интересными для использования в биотехнологии. С большой долей вероятности можно предположить, что в палеопочве грота присутствует экологическая группа метаномассиликокковых бактерий, распространенная в заболоченных местах обитания.
Все эти данные являются подтверждением выдвинутой выше гипотезы об изначальном гидроморфном происхождении палеопочвы и свидетельствуют в пользу того, что формирование почвы на дневной поверхности в начале голоцена протекало в переувлажненных условиях.
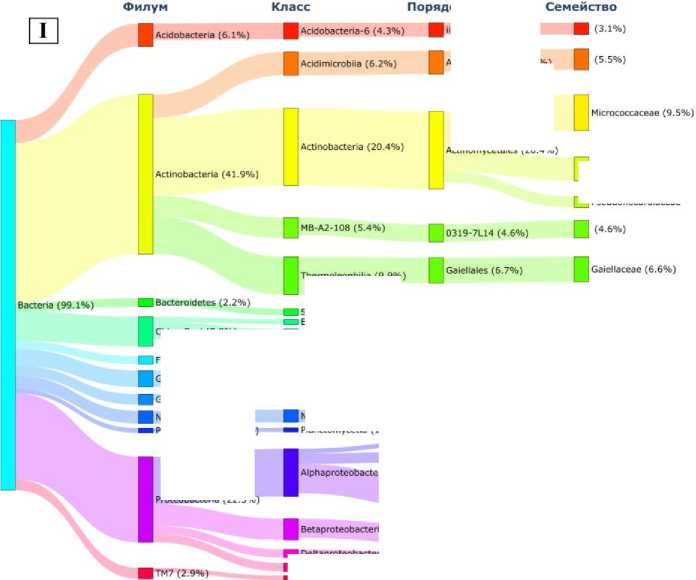
Генетическое сходство микробиомов палеопочвы и фона подтверждалось доминированием (встречаемость ДНК более 1% от общего) в прокариотных сообществах 8 филумов бактерий (рис. 3). В меньшей степени различалось содержание ДНК представителей филумов Actinobacteria (41.9 и 44.9% соответственно), Proteobacteria (22.5 и 26.3% соответственно), Bacteroidetes (2.2 и 1.8% соответственно), Gemmatimonadetes (2.8 и 2.3%, соответственно) , Nitrospirae (3.3 и 2.3% соответственно) , Planctomycetes (1.3 и 2.2%, соответственно), в большей степени – Acidobacteria
(6.1 и 11.1% соответственно), Chloroflexi (7.8 и 4.4% соответственно). Снижение содержания представителей Acidobacteria (на 5.0%) и увеличение Chloroflexi (на 3.4%) в погребенных почвах вряд ли может служить индикатором временной (эволюционной) перестройки структуры микробиома. Это связано с тем, что изменения содержания таксонов в микробиоме в пределах 5% ДНК соответствуют уровню сезонных колебаний содержания этих групп микроорганизмов в почвах (Чернов, Железова, 2020). Еще 3 филума ( ТМ7 (2.9%), GAL15 (4.3%), Firmicutes (2.0%)) доминировали в микробиоме грота, а в фоновых почвах являлись минорными компонентами. Роль первых двух до конца не изучена, их рассматривают как кандидатов в филумы. Представители Firmicutes относятся к классу Bacilli (род Bacillus, Paenibacillus, Lactobacillus, Streptococcus ). Бациллы широко встречаются в природе, предпочитают богатые органические субстраты и их доминирование в палеопочве может быть связано с деятельностью древнего человека-охотника, использующего грот в качестве стоянки для разделки туш животных, о чем свидетельствуют многочисленные артефакты, найденные в погребенной почве (Кулаков и др., 2017).
Большинство представленных на диаграмме доминантных филумов встречаются в составе прокариотных сообществ различных типов почв, образуя ассоциации и комплексно участвуя в деструкции органического вещества (Чирак и др., 2013; Chernov et al., 2018), что является еще одним подтверждением педогенной природы погребенной в гроте палеопочвы.
Однако филум Planctomycetes, являясь одной из важных и численно репрезентативных групп бактерий в доминантном микробном сообществе палеопочвы грота и фона, может преобладать только в гидроморфных условиях. Изначально планктомицеты были описаны как класс водных бактерий, который обнаруживался в образцах водорослей в морской воде (Elshahed et al., 2007). Однако с введением в практику методов молекулярной экологии они были обнаружены в торфяных сфагновых болотах (Kuli-chevskaia et al., 2006), в анаэробных источниках, богатых соединениями серы (Elshahed et al., 2007), в почвенных сообществах микроорганизмов.
Illi 15 (3-7%)
Acidimicrobiales (6.2%)
Sphingobacteriia (1.8%)
Chloroflexi (7.8%)
Firmicutes (2.1%)
SAL15 (4.3%)
Gemmatimonadetes (2.8%)
IGemm-1 (1.7%)
Nitrospira (3.3%)
Planctomycetla (1.1%)
Sphingomonadales (7.7%)
Sphingomonadaceae (6.1%)
ITM7-3 (1.2%)
— (1.2%)
Д Sollrubrobacterales (3.2%) □ (2.9%)
И] Sphingobacteriales (1.8%) ИС Sphingobacteriaceae (1.7%)
" (1.7%)
| (4.3%)
■ Rhlzoblales (2.8%) (12.6%)
^И Comamonadaceae (1.7%)
■ Oxalobacteraceae (3.3%)
■ Nitrospirales (3.3%)
^e Cauiobacteraies (1.2%)
(5.6%) ■ Burkholderiales (5.0%)
Ш Deltaproteobacteria (2.2%) e Syntrophobacterales (1.6%)
■ Gammaproteobacteria (2.2(M^HXanthomanadales (2.0%)
| (4.3%)
M (1.7%)
Tiiermoleophilia (9.9%)
ЕП1П6529 (1.4%)
■|G'tt-GS-136 (2.3%)
О SOBS (2.3%)
О Bacilli (2.1%)
| (4.3%)
Nitrospirae (3.3%)
Planrtomycetes (1.3%)
Actinomvcetales (20.4%)
| Nocardioidaceae (6.3%)
1=3 (1.4%)
О (2.3%)
О (2.3%)
EZJ Bacillales (2.0%)
П Pseudonocardiaceae (3.0%)
M (1.4%)
□ (2.3%)
■ (2.3%)
EZ3 Bacillaceae (1.3%)
И0319-6А21 (3.0%)
-
■и Caulobacteraceae (1.2%)
^И Hyphomicrobiaceae (1.3%)
-
■■ (1.4%)
Proteobacteria (22.5%)
i Hi Syntrophobacteraceae (1,6%) ■ Xanthomonadaceae (1.8%) — (1.2%) _
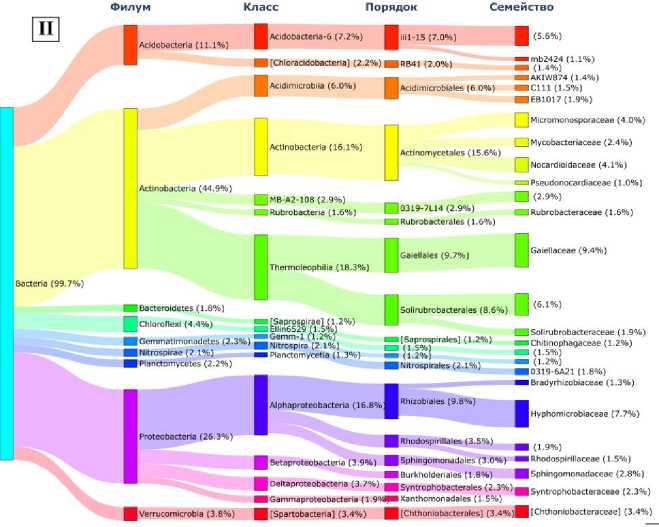
Рис.3. Содержание доминантных (более 1% от общей ДНК) таксонов в микробиоме: I – палеопочва грота Ахцу;
II – современная аллювиальная фоновая почва.
Fig. 3. Content of dominant (more than 1% of total DNA) taxa in the soil microbiome of the Akhtsu Grotto.
Из-за сложностей культивирования, перечень доступных для исследования изолятов планктомицетов остается ограниченным, причем подавляющее большинство ныне охарактеризованных представителей Planctomycetes принадлежат к классу Planctomycetia , который включает порядки Planctomycetales, Gemmatales, Isosphaerales и Pirellulales (Dedysh et al., 2020). Все перечисленные порядки определены в микробиоме палеопочвы грота и фона. Класс Phycisphaerae представлен в меньшей степени и менее изучен. Рядом исследователей была выявлена способность планктомицетов болот и торфяников к деструкции биополимеров, таких как пектин, ксилан и ламинарин (Иванова, Куличевская., 2021; Наумов и др., 2022) а также антимикробная активность пресноводного вида Gemmata palustris (порядок Gemmatales ) (Белова и др., 2020), что может предполагать наличие среди данной группы микроорганизмов видов с высоким биотехнологическим потенциалом.
Среди минорных таксонов представляли интерес цианобактерии или сине-зеленые водоросли, содержание которых от тотальной ДНК палеопочвы составляло 0.6%, а в фоновых почвах на порядок ниже – 0.05%. Среди трех классов цианобактерий в составе микробиома палеопочвы в большем количестве содержались представители класса Chloroplast порядков Streptophyta и Strame-nopiles . Цианобактерии – это морские, пресноводные и почвенные виды, участники симбиозов (например, в лишайнике), они составляют значительную долю океанического фитопланктона, способны к формированию толстых бактериальных матов, а также являются главными участниками цветения воды, которое вызывает массовые заморы рыбы и отравления животных и людей. Однако существуют цианобактерии, населяющие аридные территории, это в основном род Nostoc (Горовцов, Герасименко, 2014) , который отсутствовал в микробиоме грота.
Большое значение для науки и экологического благополучия всего человечества могут иметь представители нитчатого рода Leptolingbya, присутствующие в самых разных средах обитания, включая пресноводные, морские и наземные объекты, а также в экстремальных местах, таких как термальные источники. Данный род был определен и в палеопочве грота. В исследовании 2022 г.
впервые изучалось образование поли-3-гидроксибутирата (ПГБ) у Leptolingbya sp. NIVA-CYA 255, выделенного из египетской почвы. ПГБ – исходное вещество для производства биоразлагаемого пластика. Склонность к биоразложению при воздействии определенных активных биологических сред делает пластик на основе ПГБ потенциальной заменой полимерам на нефтехимической основе, таким как вездесущий полиэтилен высокой плотности (ПЭВП) (Kettner et al., 2022).
ВЫВОДЫ
В результате исследования группового и видового состава прокариотического сообщества палеопочвы грота Ахцу, сформированной в гидроморфных условиях притеррасной части реки Мзымта в раннеголоценовый период, было определено низкое содержание нитрификаторов и аэробных целлюлозоразлагающих микроорганизмов по сравнению с современной аллювиальной фоновой почвой. По результатам секвенирования ампликонов гена 16S - pPHK установлено, что вид домена Archaea , филума Euryarchaeota , класса Thermoplasmata , семейства Methanomas-siliicoccaceae , способный к метаногенезу, может являться индикатором переувлажнения, которое создаtт анаэробные условия. Доминирование в составе бактериоценоза филума Planctomycetes (1.3% от общей ДНК) также характеризует условия формирования палеопочвы грота как гидроморфные. Использованный комплекс микробиологических методов показал хорошую эффективность при изучении истории и условий формирования древнего палеоо-бьекта. Данные анализа подтвердили гипотезу о гидроморфном генезисе изученной палеоаллювиальной почвы в момент ее дневного формирования в начале голоцена и объяснили историю формирования геоморфологии речной долины.
Список литературы Микробиологические индикаторы раннеголоценовой притеррасной почвы долины реки Мзымта черноморского побережья Кавказа
- Белова С.Э., Салтыкова В.А., Дедыш С.Н. Антимикробная активность нового пресноводного планктомицета Lacipirellula parvula PX69T // Микробиология. 2020. Т. 89. № 5. С. 503-510. https://doi.org/10.31857/S0026365620050043.
- Водяницкий Ю.Н., Плеханова И.О. Биогеохимия тяжелых металлов в загрязненных переувлажненных почвах (аналитический обзор) // Почвоведение. 2014. № 3. С. 273-282.
- Горовцов А.В., Герасименко А.А. Родовой состав и численность цианобактерий в антропогенно-преобразованных почвах г. Волгодонска // Научно-методический электронный журнал “Концепт”. 2014. № 26. С. 326-330. URL: http://e-koncept.ru/2014/64366.htm.
- Дёмкин В.А., Ельцов М.В., Дёмкина Т.С., Хомутова Т.Э. Палеопочвы археологических памятников степной зоны как индикаторы развития природной среды в голоцене // Вестник Вестник Тамбовского университета. Серия: Естественные и технические науки. 2013. Т. 18. Вып. 3. С. 966-970.
- Дёмкина Т.С. Пространственно-временная динамика состояния микробных сообществ почв степей Волго-Донского междуречья // Аридные экосистемы. 2020. Т. 26. № 1(82). С. 76-83.
- Добровольский Г.В., Балабко П.Н., Стасюк Н.В., Быкова Е.П. Аллювиальные почвы речных пойм и дельт и их зональные отличия // Аридные экосистемы. 2011. Т. 17. № 3(48). С. 5-13.
- Иванова А.А., Куличевская И.С. Gemmata palustris sp. nov. Новый планктомицет, выделенный из низинного болота севера России // Микробиология. 2021. Т. 90. № 5. С. 564-573.
- Кулаков С.А., Кизилов А.С., Дятлов А.С. Открытие нового памятника мезолита в Сочинском Причерноморье // Археологические открытия. 2017. Т. 2015. С. 233-234.
- Наумов Д.Г., Куличевская И.С., Дедыш С.Н. Генетические детерминанты утилизации ксилана у планктомицета класса Phycisphaerae, Humisphaera borealisM1803 T // Микробиология. 2022. Т. 91. № 3. С. 300-311. https://doi.org/10.31857/S0026365622300048.
- Практикум по агрохимии / под ред. В.Г. Минеева. М.: Изд-во МГУ, 2001. 687 c.
- Практикум по микробиологии / под ред. М.А. Егорова, Л.М. Захарчук, А.И. Нетрусова. М.: Изд-во Академия, 2005. 608 c.
- Плеханова Л.Н. Антропогенная деградация почв речных террас Волго-Уральского региона в эпоху бронзы и ее влияние на современный почвенно-растительный покров // Аридная экосистема. 2019. Т. 25. № 3 (80). С. 187-192. https://doi.org/10.24411/1993-3916-2019-10065.
- Чернов Т.И., Железова А.Д. Динамика микробных сообществ почвы в различных диапазонах времени (обзор) // Почвоведение. 2020. № 5. C. 590-600. https://doi.org/10.31857/S0032180X20050044.
- Чевердин Ю.И., Титова Т.В., Беспалов В.А., Гармашова Л.В. Мониторинг микробиологических параметров сезонно переувлажненных почв Каменной Степи // Научный альманах. 2015. № 4. C. 259-265. https://doi.org/10.17117/na.2015.04.
- Чирак Е.Л., Першина Е.В., Дольник А.С., Крутовая О.В., Василенко Е.С., Когут Б.М., Мерзлякова Я.В., Андронов Е.Е. Таксономическая структура микробных сообществ в почвах различных типов по данным высокопроизводительного секвенирования библиотек гена 16S-рРНК // Сельскохозяйственная биология. 2013. № 3. С. 100-109. https://doi.org/10.15389/agrobiology.2013.3.100rus.
- Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация почв России. М.: Изд-во Почвенный институт им. В.В. Докучаева РАСХН, 2008. 343 с.
- Borrel G.O., Toole P.W., Harris H.M., Peyret P., Brugere J.F., Gribaldo S. Phylogenomic data support a seventh order of Methylotrophic methanogens and provide insights into the evolution of Methanogenesis // Genome Biology and Evolution. 2013. Vol. 5. No. 10. Р. 1769-1780. https://doi.org/10.1093/gbe/evt128.
- Chernov T.I., Zhelezova A.D., Kutovaya O.V., Tkhakakhova A.K., Bgazhba N.A., Kurbanova F.G., Makeev A.O., Puzanova T.A., Rusakov A.V., Khokhlova O.S. Comparative analysis of the structure of buried and surface soils by analysis of microbial DNA // Microbiology. 2018. Vol. 87. No. 6. P. 833-841. https://doi.org/10.1134/S0026261718060073.
- Dedysh S.N., Kulichevskaya I.S., Beletsky A.V., Ivanova A.A., Rijpstra W.I.C., Sinninghe-Damsté J.S., Mardanov A.V., Ravin N.V. Lacipirellula parvula gen. nov., sp. nov., representing a lineage of planctomycetes widespread in low-oxygen habitats, description of the family Lacipirellulaceae fam. nov. and proposal of the orders Pirellulales ord. nov., Gemmatales ord. nov. and Isosphaerales ord. nov // Systematic and Applied Microbiology. 2020. Vol. 43. No. 1. P. 126050. https://doi.org/10.1016/j.syapm.2019.126050.
- Elshahed M.S., Youssef N.H., Luo Q., Najar F.Z., Roe B.A., Sisk T.M., Bühring S.I., Hinrichs K., Krumholz L.R. Phylogenetic and metabolic diversity of Planctomycetes from anaerobic, sulfide- and sulfur-rich Zodletone Spring, Oklahoma // Applied and environmental microbiology. 2007. Vol. 73. No. 15. Р. 4707-4716. https://doi.org/10.1128/AEM.00591-07.
- Frindte K., Lehndorff E., Vlaminck S., Werner K., Kehl M., Khormali F., Knief C. Evidence for signatures of ancient microbial life in paleosols // Scientific reports. 2020. Vol. 10. No. 1. https://doi.org/10.1038/s41598-020-73938-9.
- Hsu C-C., Tsai H., Huang W-S., Huang S-T. Carbon Storage along with Soil Profile: An Example of Soil Chronosequence from the Fluvial Terraces on the Pakua Ta-bleland, Taiwan // Land. 2021. Vol. 10. No. 5. https://doi.org/10.3390/land10050447.
- Kettner A., Noll M., Griehl C. Leptolyngbya sp. NIVA-CYA 255, a promising candidate for poly (3-hydroxybutyrate) production under mixotrophic deficiency conditions // Biomolecules. 2022. Vol. 12. No. 4. https://doi.org/10.3390/biom12040504.
- Kovaleva N.O., Stolpnikova E.M. Volcanic Soil Series of the Lesser Caucasus as an Archive of Early Pleistocene Paleoecological Information // Paleontological Journal. 2020. Vol. 8. No. 54. Р. 872-881. https://doi.org/10.1134/S0031030120080092.
- Kulichevskaia I.S., Pankratov T.A., Dedysh S.N. Detection of representatives of the Planctomycetes in sphagnum peat bogs by molecular and cultivation methods // Microbiology. 2006. Vol. 75. No. 3. Р. 389-396. https://doi.org/10.1134/S0026261706030155.
- Kuznetsova A.I., Ivanova E.A., Samylina O.S., Pimenov N.V., Ivanova E.A., Kurbanova F.G., Gruzdev D.S. Prokaryotic communities in saline soils of the Lake Elton Area in a Soil Catena along the Khara River // Microbiology. 2020. Vol. 89. No. 6. Р. 670-684. https://doi.org/10.1134/S0026261720060119.
- Mack G.H., James W.C., Moger H.C. Classification of paleosols // Geological society of America bulletin. 1993. Vol. 105. No. 2. Р. 129-136. https://doi.org/10.1130/0016-7606(1993)105<1635:COPDAR-2.3.CO;2.
- Manucharova N.A., Cheptsov V.S., Belov A.A., Vorobyeva E.A., Zenova G.M., Stepanov A.L. Soil as a natural bank of microbial diversity: new approaches and relevant aspects // Bulletin of the Russian foundation for fundamental research. 2020. Vol. 106. No. 2. Р. 88-100. https://doi.org/10.22204/2410-4639-2020-106-02-88-100.
- Sedov S., Sheinkman V., Bezrukova E., Zazovskaya E., Yurtaev A. Sartanian (MIS 2) ice wedge pseudomorphs with hydromorphic pedosediments in the north of West Siberia as an indicator for paleoenvironmental reconstruction and stratigraphic correlation // Quaternary International. 2022. Vol. 632. Р. 192-205. https://doi.org/10.1016/j.quaint.2022.05.002.
- Sollinger A., Schwab C., Weinmaier T., Loy A., Tveit А.T., Schleper C., Urich T. Phylogenetic and genomic analysis of Methanomassiliicoccales in wetlands and animal intestinal tracts reveals clade-specific habitat preferences // FEMS Microbiology Ecology. 2016. Vol. 92. No. 1. https://doi.org/10.1093/femsec/fiv149.
- World Reference Base for Soil Resources International soil classification system for naming soils and creating legends for soil maps / edited by: P. Schad, C. Huysteen, E. Michéli. World Soil Resources Reports. Rome. 2014. 203 р.
- Zakharikhina L.V., Rogozhina E.V., Kizilov A.S. Paleosols оf archaeological sites in the Sochi Black Sea coast // Natural volatiles & essential oils journal. 2021. Vol. 8. No. 4. Р. 8004-8036. URL: https://www.nveo.org/index.php/journal/article/view/1668/1457.
