Микробиологический синтез никотинамидадениндинуклеотида и его ингибирование

Автор: Цыренов В.Ж., Санданов А.А., Козин В.А.
Журнал: Вестник Восточно-Сибирского государственного университета технологий и управления @vestnik-esstu
Рубрика: Биологические науки
Статья в выпуске: 5 (50), 2014 года.
Бесплатный доступ
Осуществлен микробиологический синтез никотинамидадениндинуклеотида (НАД) различными коринеподобными бактериями. Исследовано ингибирующее действие НАД и НАДФ. Установлено, что их действие направлено на аденилаткиназу.
Над, salvage биосинтез, ингибирование, аденилаткиназа
Короткий адрес: https://sciup.org/142142931
IDR: 142142931 | УДК: 615.9+577,472
Nad microbiological synthesis and its inhibition
The article describes the microbiological synthesis of NAD by various coryneform bacteria. The inhibitory effect of NAD and NADP is studied. It is established that their action is directed to adenylate kinase.
Текст научной статьи Микробиологический синтез никотинамидадениндинуклеотида и его ингибирование
Никотинамидадениндинуклеотид (НАД) это ценное биологически активное вещество, которое находит применение в медицине в качестве лекарства-иммуномодулятора, а также как реактив и компонент диагностикумов. В настоящее время производство НАД в России отсутствует.
Перспективным методом получения НАД является сэлвидж-синтез, осуществляемый штаммами-продуцентами из группы коринеподобных бактерий.
В статье приведены сравнительные исследования различных штаммов коринеподобных бактерий на предмет возможности их использования в качестве биологических объектов для постановки микробиологического синтеза НАД. Исследованы особенности процессов регуляции фосфорилирования в аденилатной ветви сэлвидж-синтеза НАД, в частности, ингибирование синтеза.
Материалы и методы
Микроорганизмы, культивирование
Объектом исследования явились штаммы коринебактерий: Corynebacterium ammoniagenes ВСТИ 403, Corynebacterium flavum ВСТИ 301, Corynebacterium species ВСТИ 4, коллекция кафедры биотехнологии ВСГУТУ, коллекция ФГУП «ГосНИИгенети-ка». Corynebacterium ammoniagenes АТСС 6872 коллекция Института биохимии им А.Н. Баха, а также типичные представители родов коринеподобных бактерий Arthrobacter citreus 278, Corynebacterium insidiosum 246, Corynebacterium stationis 336, Corynebacterium linens 242 Rhodococcus globerulus 226.
Подготовка питательной среды
Посевная среда [1] содержала: 20 г D-глюкозы моногидрат, 10 г казеинового пептона, 10 г дрожжевого экстракта и 2,5 г NaCL на 1 л воды. Перед стерилизацией pH доводили до 7,23 Н NaOH. Инокулят (10%) готовили на посевной среде. Культуру микроорганизмов объемом по 50 мл выращивали в колбах Эрленмейера объемом 750 мл.
Ферментационная среда в 1 л содержала: 100 г глюкозы моногидрата, дрожжевого экстракта, 10 г KH2PO4, 10 г K2HPO4, 10 г MgSO4∙7H2O; 6 г мочевины, стерилизованной отдель- но, и биотин (30 мкг/л) добавляли после фильтрации, начальный рН 7,4. Для удовлетворения потребности C. ammoniagenes в марганце в эту среду добавляли 2 мкМ MnCL2.
Среду по 50 мл разливали в качалочные колбы на 750 мл, доводили рН 5 н, NaOH до 7,6 и стерилизовали в течение 15 мин при 1 атм. Мочевину стерилизовали отдельно при 0,5 атм в течение 15 мин и добавляли в колбы (0,6%) после стерилизации. Биотин стерилизовали пропусканием через 0,22 мкм фильтр и добавляли в колбы в количестве 30 мкг/л. Фосфаты (К 2 НРО 4 + КН 2 РО 4 ) готовили отдельно и добавляли в колбы к основной части ферментационной среды.
Условия ферментации
Культуру с 2-суточного косяка петлей переносили в 50 мл посевной среды и помещали на качалку (220 об/мин). После суток роста при 30±1°С посевной культурой в количестве 10 об.% инокулировали среду для биосинтеза. Для осуществления сэлвидж-синтеза в среду для биосинтеза добавляли предшественники: аденин никотинамид по 2 мг/мл соответственно на 2-е сут ферментации. Продолжительность ферментации составляла 2-4 сут.
Определение роста
Наблюдения за ростом клеток проводили измерением оптической плотности при 578 нм. Отбирали 1 мл аликвоту культуральной жидкости, разбавляли ее 3 мл дистиллированной воды и добавляли 100 мкл 2 Н HCL с целью растворения осажденных кристаллов. Для предотвращения слипания клеток бактерий добавляли 5 мкл Твин 80 (20% вес/объем). Осадок после низкоскоростного центрифугирования ресуспендировали в 1 мл дистиллированной воды и измеряли мутность. Для вычисления сухого веса пользовались калибровочной кривой.
Идентификация и количественное определение НАД и нуклеотидов
Идентификацию и количественное определение НАД и нуклеотидов осуществляли после отделения клеток на низкоскоростной центрифуге. Высокопроизводительная жидкостная хроматография (ВЭЖХ) НАД с градиентной элюцией выполнялась по методу [4] с использованием хроматографа Agilent 1100 (производство США), оснащенного ультрафиолетовым детектором переменной частоты, установленным на 254 нм. Сольвент А содержал 20 мл концентрированной фосфорной кислоты в 800 мл деионизованной воды доведенным до 4,0 рН с помощью 25% NaOH, и объемом, доведенным до 1 л. Сольвент В содержал равные объемы ацетонитрила и метанола. Все реактивы пропускали через фильтр (5 мкм). Нуклеотиды элюировали при 37 ºС на колонке С 18 5 µ. Для использования в качестве стандартов в сольвенте А растворяли рибонуклеотиды 5'-АМФ и АТФ (0,1 М стандартный раствор). Скорость элюции во всех опытах устанавливали 0,7 мл/мин.
Результаты и обсуждение
Исследование способности коринеподобных бактерий к сэлвидж-синтезу НАД
В качестве объектов исследования по отбору активного штамма-продуцента НАД использовано 11 культур коринеподобных бактерий. Изучение роста бактерий показало, что все отобранные штаммы-продуценты накапливали биомассу в заданных условиях. Характеры кривых роста выбранных штаммов-продуцентов идентичны. Наибольший рост был показан у С. Аmmoniagenes ВСТИ 404 , а наименьший – у Arthrobcter citreus 278.
Исследование биосинтеза показало, что накопление НАД лучше всего происходит в присутствии детергентов. Наиболее эффективным оказался цетилпиридиний хлорид в концентрациях 1-2 мг/мл (табл.). Результаты ВЭЖХ показали, что в условиях ферментации, когда в культуру микроорганизмов вносятся предшественники – аденин и никотинамид, наблюдается биосинтез НАД (рис. 3).
При использовании в синтезе С. аmmoniagenes (Brevibacterium) АТСС 6872, многократно описанного штамма-продуцента нуклеотидов и НАД, а также продуцента нуклеотидов, С. аmmoniagenes ВСТИ 404 наблюдается значительное накопление НАД. Другие микроорганизмы синтезировали небольшие количества НАД (до 0,5 мг/мл), АТФ, а также следовые количества УФ-поглощающих веществ (рис. 1).
Таблица
Динамика накопления НАД в процессе микробиологического синтеза
|
Микроорганизмы |
Выход НАД, мг/мл |
|||||
|
12 ч |
24 ч |
48 ч |
||||
|
О |
К |
О |
К |
О |
К |
|
|
C. ammoniagenes АТСС 6872 |
1,38 |
0,21 |
2,04 |
0,16 |
2,59 |
0,71 |
|
C.ammoniagenes ВСТИ 404 |
1,9 |
0,2 |
3,16 |
0,64 |
2,68 |
0,67 |
|
C. flavum ВСТИ 301 |
0,35 |
0,31 |
0,52 |
0,26 |
0,43 |
0,37 |
|
C. ammoniagenes ВСТИ 403 |
1,56 |
0,18 |
3,27 |
0,76 |
3,04 |
0,60 |
|
A. citreus 278 |
0,05 |
0,25 |
0,11 |
0,05 |
0,20 |
0,01 |
|
C. insidiosum 246 |
0,09 |
0,25 |
0,19 |
0,05 |
0,09 |
0,07 |
|
C. stationis 336 |
0,35 |
0,17 |
0,38 |
0,37 |
0,31 |
0,4 |
|
C. linens 242 |
0,4 |
0,11 |
0,31 |
0,24 |
0,19 |
0,22 |
|
R. globerulus 226 |
0,17 |
0,15 |
0,29 |
0,34 |
0,26 |
0,31 |
|
Arthrobcter species ВСТИ 5 |
0,1 |
0,12 |
0,25 |
0,2 |
0,11 |
0,21 |
Примечание. О опыт, ферментация в присутствии детергента; К контроль, ферментация в отсутствие детергента. Аденин, никотинамид ЦПХ добавляли по 2 мг/мл
В ходе исследования динамики рН было показано, что в процессе ферментации НАД в первые 12 ч происходит увеличение рН 6,7 до 8,4 у C. ammoniagenes АТСС 6872 и с 7 до 8,5 у C. ammoniagenes ВСТИ 403 . После 12 ч культивирования наблюдали стабилизацию рН до 7 в обоих случаях ферментации: при использовании штаммов-продуцентов C. ammoniagenes АТСС 6872 и C. ammoniagenes ВСТИ 404 .
Установлено, что наибольшей способностью к синтезу НАД обладают штаммы C. ammoniagenes ВСТИ 404 и C. ammoniagenes ВСТИ 403 . Их положительным свойством является то, что их способность синтезировать нуклеотиды не лимитируется содержанием Mn++ в среде культивирования.
Накопление НАД в условиях сверхсинтеза не достигало 100% выхода от количества внесенных предшественников. Это позволило предположить, что при сверхсинтезе НАД действуют регуляторные механизмы, одним из которых, возможно, является ретроингибирование.
Внесение в культуральную среду экзогенных НАД, НАДФ и НАДН показало, что эти соединения ингибировали сверхсинтез НАД. Ингибирующий эффект НАД проявляется при концентрации его 4 мг/мл. НАДФ оказывал более сильный эффект (рис. 1).
Известно, что ферменты сэлвидж-синтеза НАД находятся под контролем АТР, потому представлялось интересным выяснить, направлено ли обнаруженное нами ретроингибирующее действие НАД и НАДФ на образование АТР, являющегося промежуточным метаболитом синтеза НАД.
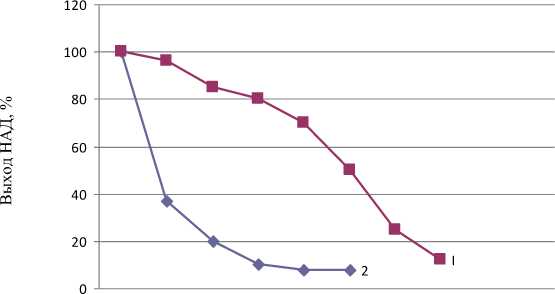
Рис. 1. Влияние НАД (I) и НАДФ (2) на сверхсинтез НАД (время синтеза 45 ч)
В отсутствие пиридинового предшественника и в присутствии АМФ наблюдался сверхсинтез АТР. При изучении влияния НАД и НАДФ на образование АТР из АМР было обнаружено, что уровень накопления АТР из АМР снижался при внесении в среду НАД (3 мкмоль/мл) и НАДФ (2 мкмоль/мл). НАД ингибировал синтез АТР на 56%, НАДФ на 17%. Поскольку в качестве предшественника использовался АМР, заключили, что одной из точек приложения ингибирующего действия НАД и НАДФ на сверхсинтез НАД является синтез АТР и АМР.
Для подтверждения этих результатов исследована активность фосфорилирования АМР бесклеточными экстрактами в качестве источника фосфорилирующих ферментов с использованием радиоактивного [С14] – АМР.
Максимальное угнетение реакции фосфорилирования АМР наблюдалось при одной и той же концентрации обоих эффектов – 5 мМ (рис. 2).
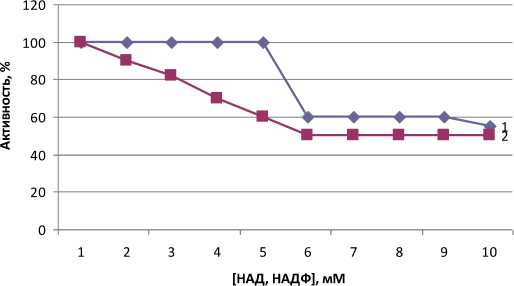
Рис. 2. Влияние НАД (I) и НАДФ (2) на синтез АТФ бесклеточными C. ammoniagenes
Для того чтобы отделить аденилаткиназу от нуклеозиддифосфосиназы, при очистке использовали кислотную обработку бесклеточного экстракта С. ammoniagenes АТСС 6872 I NHCL до рН 3 с последующим дробным высаливанием сернокислым аммонием от 35 до 55 % насыщения.
Активность аденилаткиназы ингибировалась на 50% под действием НАД и НАДФ при концентрации эффекторов 3 и 3,5 мМ соответственно (рис. 3).
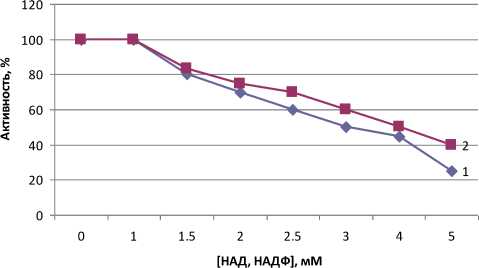
Рис. 3. Ингибирование NАD (I) и NADР (2) частично очищенной аденилаткиназы
Выводы
-
1. Для осуществления микробиологического синтеза никотинамидадениндинуклеотида (НАД) методом ферментации отобраны штаммы-продуценты Corynebacterium ammoniagenes ВСТИ 404 и Corynebacterium ammoniagenes ВСТИ 310.
-
2. Установлено, что ингибирующее влияние микробиологического синтеза НАД осуществляется конечными продуктами синтеза.
