Микробные комплексы филлосферы как индикаторы состояния древостоев-эдификаторов

Автор: Сорокин Н.Д., Афанасова Е.Н., Сенашова В.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 9, 2016 года.
Бесплатный доступ
Целью работы являлось исследование формирования микробных комплексов фил-лосферы на ценотическом, популяционном и видовом уровне как индикаторов состояния древостоев-эдификаторов при разных внеш-них воздействиях. При анализе микрофлоры филлосферы древостоев Дороховского (зона интенсивного влияния ГРЭС) и Захаринского (в 70-80 км от ГРЭС) боров Красноярского края в зоне КАТЭКа установлено, что число микробных клеток на 1 г хвои в Дороховском бору в два раза больше, чем в Захаринском. Существенные различия отмечаются в каче-ственном составе микроорганизмов. Эпифи-ты Захаринского бора на 96 % представлены неспорообразующими Гр(-) бактериями, близ-кими по таксономическому положению к роду Pseudomonas. В Дороховском бору 70 % эпи-фитов составляют пигментные формы. Чис-ленность эпифитных микроорганизмов куль-тур сосны и лиственницы в зоне активного техногенеза превосходит таковую на кон-трольном участке в 7-9 раз. Здесь доминиру-ют спороносные бактерии (92 % на сосне и 96 % на лиственнице), а на культурах сосны контрольного участка - неспорообразующие формы (90 %). В исследуемом районе (Нижнее Приангарье) черный пихтовый усач переносит комплекс грибов синевы древесины, представ-ленный главным образом видами Ophiostoma curvicollis, Ophiostoma sp.и Leptographium sp. Общая частота встречаемости этого ком-плекса в ходах вредителя достигла 90-100 % на пробных площадях в дефолиированных шел-копрядом древостоях и не превышала 60 % на контрольном участке в ненарушенном древо- стое. Исследование видового состава и чис-ленности микромицетов коры ели и пихты свидетельствует о зависимости этих пока-зателей от вида и степени дефолиации кроны растения-хозяина и таким образом может яв-ляться адекватным индикатором состояния дерева на ранних стадиях его поражения. Ана-лиз микрофлоры здоровой и пораженной хвои сосны обыкновенной и ели сибирской в лесо-питомниках показывает, что мицелиальные грибы являются постоянными ее обитателя-ми, но представлены в разном соотношении с преобладанием на больной хвое, у которой снижена фитонцидная активность.
Микробные комплексы, филлосфера, древостои, офиостомовые мик-ромицеты, антропогенные (техногенные) воздействия
Короткий адрес: https://sciup.org/14084801
IDR: 14084801 | УДК: 631.41
Microbial phyllosphere complexes as status indicators of tree stands- edificators
The aim of the research was to study the for-mation of microbial phyllosphere complexes in coe-notic, population and species levels as indicators of tree stands- edificator under different external influ-ences. In the analysis of microflora phyllosphere tree stands were Dorokhovsky (zone of intensive effect HES) and Zakharinsky (70- 80 km from HES) pinery found that the number of microbial cells in 1 g of pine needles in the Dorokhovsky forest twice more than in the Zakha-rinsky. Significant differences were observed in the qualitative composition of microorganisms. Epi-phytes of Zakharinsky pinery are represented by 96 % non-spore Gr-bacteria, similar in taxonomic posi-tion of the genus Pseudomonas. In Dorokhovsky pinery epiphytes comprise 70 % pigment forms. The number of epiphytic microorganisms of cul-tures of pine and larch in the area of active techno-genesis exceeds that of the control plot is almost 7-9 times. Here sporeforming bacteria predominate (92 % to 96 % pine and larch), and the control plot of pine cultures - asporogenous form (90 %). In the study the area (Lower Angara) a black fir sawyer beetle carried a complex of blue stain fungi pre-sented species Ophiostoma curvicollis, Ophiostoma sp.and Leptographium sp. The total frequency of occurrence of this complex in the passages of the pest reached 90-100 % in test plots in defoliation by silkworm tree stands and does not exceed 60 % in the control plot in the undisturbed tree stand. The investigation of species composition and abun-dance miсromycetes of bark of spruce and fir shows the dependence of these parameters on the type and degree of defoliation of the crown of the plant host, and thus may be an adequate indicator of the condition of the tree in the early stages of its lesion. The analysis of microflora of healthy and diseased needles of Scots pine and Siberian spruce in forestry indicates that mycelium fungi are its permanent inhabitants but presented in a differ-ent ratio to the prevalence in the diseased needles, which phytoncide activity is reduced.
Текст научной статьи Микробные комплексы филлосферы как индикаторы состояния древостоев-эдификаторов
Введение . Микробные реакции на воздействие различных нарушающих факторов проявляются быстро и достаточно отчетливо, что позволяет в короткие сроки выявить наиболее нарушенные экологические зоны, экосистемы и отдельные компоненты, прогнозировать их состояние при сохранении или устранении антропогенного фактора [2, 3, 5, 7, 11, 12].
Из приведенных в литературе материалов очевидно, что наиболее полно микробиологическая индикация и диагностика исследована для почвенных и ризосферных биогоризонтов лесных экосистем и связана в большей степени с антропогенными (техногенными) нарушениями корнеобитаемой зоны фитоценоза. Однако верхняя часть фитоценозов филлосфера также подвергается мощному антропогенному влиянию поллютантов промышленных предприятий, радиоактивному загрязнению и дополнительному воздействию насекомых-вредителей и микроорганизмов фитопатогенов. Поэтому ассоциации эпифитных микроорганизмов, обитающих на поверхности листовых пластинок и корней, принимают на себя первый «удар» техногенеза, выполняют функцию защитников и индикаторов состояния дерева. При этом индикаторная роль микроорганизмов филлосферы древесных видов растений изучена слабо.
Цель исследования : изучение формирования микробных комплексов филлосферы на ценоти-ческом, популяционном и видовом уровне как индикаторов состояния древостоев-эдификаторов при разного рода внешних воздействиях .
Объекты и методы исследования. Объектом исследования в зоне КАТЭКа Красноярского края были микробные сообщества филлосферы сосновых насаждений Дороховского, Пионерского, Захаринского боров и Ададымских березовых насаждений. Эпифитные микроорганизмы изучались также на культурах сосны и лиственницы. Все указанные участки расположены в 5–7 км от факела техногенных выбросов Назаровской ГРЭС; вне сферы влияния поллютантов находятся Ададымские березовые насаждения и культуры сосны южного склона хребта Арга. Сбор материала для микробиологического анализа проводился в период вегетации взрослых деревьев и культур в 2012–2014 гг. Образцы хвои отбирались на тридцати модельных деревьях. Всего проанализировано 90 образцов.
В Хакасии отбирали образцы для микологического анализа офиостомовых грибов фил-лосферы в зоне лесостепных лиственничников, отличающихся повышенной теплообеспеченно-стью, оптимальной для развития короеда Ips cembrae – основного переносчика микромице-тов. Образцы пораженной древесины собирали также на гарях в сосновых и сосноволиственничных древостоях Нижнего Приангарья.
Объектом исследования в лесопитомниках Маганского и Уярского лесничеств Красноярского края являлась эпифитная микрофлора здоровой и больной хвои двух древесных пород: сосны обыкновенной ( Pinus sylvestris ) и ели сибирской ( Picea obovata ).
При анализе эпифитной микрофлоры брали навеску 1 г хвои и делали смыв стерильной водой на качалке типа АВУ-6с в течение 10 мин. Для выявления грибов смыв с хвои (0,1 мл) высевали методом Коха на плотную среду Чапека. Бактерии определяли на рыбо-пептонном агаре (РПА), актиномицеты – на крахмало-аммиачном агаре. Культивирование проводили при температуре 25 ° С. Микробиологический посев проводился в 3-кратной повторности. Достоверность результатов посева обеспечивают 270 определений. Данные обрабатывались при помощи Microsoft Excel 97 и Statistica 5. Все данные достоверны при уровне значимости p = 0,05.
Выросшие колонии микроорганизмов микро-скопировали при увеличении в 1350 раз. Проводили количественный учет колоний споровых и неспоровых бактериальных форм, дрожжей, мицелиальных грибов, актиномицетов. С помощью метода Грегерсона (Gregersen) устанавливали способность бактериальных форм окрашиваться по Граму.
Идентификацию выделенных культур эпифитных микроорганизмов осуществляли по морфологическим, культуральным и физиоло- гическим признакам с использованием определителей [4, 6, 8, 10].
Виды офиостомовых грибов идентифицировали на основании морфологических признаков анаморф и телеоморф [1].
Результаты исследования и их обсуждение. При анализе микрофлоры филлосферы древостоев Дороховского (зона интенсивного влияния ГРЭС) и Захаринского (в 70–80 км от ГРЭС) боров установлено, что число микробных клеток на 1 г хвои в Дороховском бору в два раза больше, чем в Захаринском (табл. 1). Существенные различия отмечаются в качественном составе микроорганизмов. Эпифиты Захарин-ского бора на 96 % представлены неспорообразующими Гр(-) бактериями, близкими по таксономическому положению к роду Pseudomonas . В Дороховском бору 70 % эпифитов составляют пигментные формы. Эпифитная микрофлора Ададымских березовых насаждений в численном отношении значительно превосходит микрофлору Дороховского и Захаринского боров и достигает 52 840 тыс. КОЕ на 1 г хвои. Объясняется это различие разным фитонцидным действием на микрофлору лиственных и хвойных пород. В составе микроорганизмов на листьях березовых насаждений возрастает до 30 % количество спорообразующих бактерий преимущественно рода Bacillus .
Численность эпифитных микроорганизмов культур сосны и лиственницы в зоне активного техногенеза превосходит таковую на контрольном участке в 7–9 раз . Здесь доминируют спороносные бактерии (92 % на сосне и 96 % на лиственнице), а на культурах сосны контрольного участка – неспорообразующие формы (90 %). Такая существенная разница в составе микро-боценозов филлосферы объясняется влиянием частиц техногенных выбросов. За счет увеличения в микробных комплексах спорообразующих форм микроорганизмов повышается их устойчивость к высоким концентрациям техногенных загрязнителей, в составе которых преобладают соединения кальция, железа, магния, марганца, стронция, бария и кадмия.
При учете биомассы эпифитных микроорганизмов и исследовании физиологической активности доминантных культур отмечено, что по мере приближения насаждений к источнику загрязнения (Назаровская ГРЭС) биомасса микроорганизмов возрастает, а физиологическая активность по отношению к азоту снижается. Снижение ферментативной активности микро- организмов следует расценивать как вреадап-тивную реакцию на те компоненты, которые попадают на поверхность древостоев в результате осаждения поллютантов.
При исследовании филлосферы древесных насаждений в лесных экосистемах Сибири вместе с эпифитными бактериями в качестве индикаторных микроорганизмов для оценки состояния дерева использовали офиостомовые грибы, ассоциированные в природе с насекомыми-ксилофагами [1, 9]. Установлено, что грибы синевы древесины являются обязательным компонентом грибных сообществ в ходах короеда-типографа и черного пихтового усача, доминируя среди других мицелиальных форм на первых этапах сукцессии грибов в поврежденных растительных тканях ели и пихты. В дефолии-рованных сибирским шелкопрядом древостоях ели и пихты сибирской грибы синевы древесины являются обязательным компонентом и основой микобиоты черного пихтового усача и короеда-типографа. Зарегистрированная частота встречаемости грибов свидетельствует об эпидемическом уровне их распространения в популяции вредителя на поврежденных участках (табл. 2). В исследуемом районе (Нижнее Приангарье) черный пихтовый усач переносит комплекс грибов синевы древесины, представленный главным образом видами Ophiostoma curvicollis, Ophiostoma sp. и Leptographium sp. Общая частота встречаемости этого комплекса в ходах вредителя достигла 90–100 % на пробных пло- щадях в дефолиированных шелкопрядом древостоях и не превышала 60 % на контрольном участке в ненарушенном древостое. Исследование видового состава и численности микро-мицетов коры ели и пихты позволяет говорить о зависимости этих показателей от вида и степени дефолиации кроны растения-хозяина и таким образом может являться адекватным индикатором состояния дерева на ранних стадиях его поражения.
В ситуации, когда есть визуальные признаки поражения деревьев фитопатогенами (инфицированная и здоровая хвоя) комплексы эпифитных микроорганизмов существенно различаются по качественному и количественному составу. Анализ микрофлоры здоровой и пораженной хвои сосны обыкновенной ( Pinus sylvestris ) в лесопитомниках свидетельствует, что мицелиальные грибы являются постоянными ее обитателями, но представлены в разном соотношении с преобладанием на больной хвое. Максимальная численность эпифитных микроорганизмов на здоровой (283 · 10³ КОЕ) и пораженной (127 · 10³ КОЕ) хвое приходится на первую декаду июня и затем плавно понижается (рис.). Новый скачок численности, но меньший по уровню, у здоровой хвои регистрируется в сентябре – 33 · 10³ КОЕ, а у больной хвои – в августе (115 · 10³ КОЕ). У здоровой хвои в первой декаде июня и в третьей декаде сентября дрожжи являются доминирующей культурой: их доля 97,5 и 47,5 % соответственно.
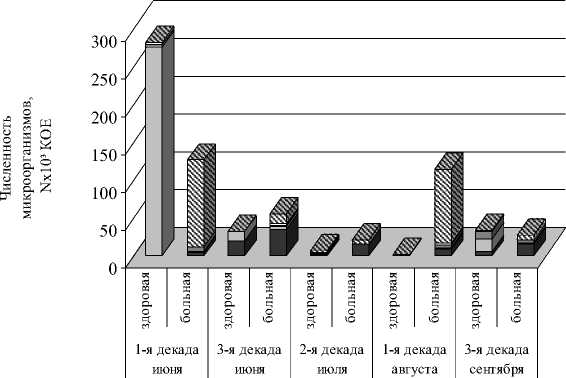
Мицелиальные грибы Дрожжевые грибы Актиномицеты Споровые бактерии Неспоровые бактерии
Динамика численности эпифитной микрофлоры сосны обыкновенной
Таблица 1
|
Район |
Число клеток на 1 г хвои или листьев |
Биомасса, мг/г хвои или листьев |
Преобладающая форма |
Положительная реакция, % |
||||
|
на органический азот |
на неорганический азот |
на крахмал |
на желатину |
на каталазу |
||||
|
Ададымс-кие насаждения |
52840 |
0,21 |
Спорообразующие (30 %) |
84 |
82 |
67 |
36 |
73 |
|
Захаринский бор |
1400 |
0,012 |
Неспоровые (96 %) |
57 |
64 |
72 |
36 |
63 |
|
Дороховский бор |
2900 |
0,013 |
Пигментные (70 %) |
37 |
46 |
6 |
13 |
7 |
|
Хребет Арга |
2100 |
0,005 |
Неспоровые (86 %) |
44 |
42 |
22 |
11 |
10 |
|
Культуры сосны |
14500 |
0,05 |
Спорообразующие (92 %) |
10 |
25 |
17 |
14 |
12 |
|
Культуры лиственницы |
18400 |
0,06 |
Спорообразующие (96 %) |
40 |
30 |
26 |
21 |
18 |
Численность, биомасса и физиологическая активность эпифитных микроорганизмов
Частота встречаемости грибов синевы древесины в тканях пихты сибирской, поврежденной черным пихтовым усачом
Таблица 2
|
Тип леса |
Период дефолиации |
Встречаемость грибов, % |
|||
|
общая |
Leptographium sp . |
Ophiostoma sp. |
O. curvicollis |
||
|
Контроль |
0 |
60 |
60 |
0 |
0 |
|
ПОР |
1994–1995 гг. |
96 |
68 |
28 |
60 |
|
ПОР |
1995–1996 гг. |
100 |
88 |
52 |
16 |
|
ПМЗ |
1995 г. |
100 |
100 |
63 |
29 |
|
ПРО |
1994–1995 гг. |
84 |
68 |
52 |
48 |
Примечание: ПОР – пихтарник осочково-разнотравный; ПРО – пихтарник разнотравно-осочковый; ПМЗ – пихтарник мелкотравно-зеленомошный.
Вестник КрасГАУ. 2016. №9
Заключение. Результаты анализа микробных комплексов филлосферы древесных пород в зоне активного антропогенного (техногенного) воздействия показывают, что микробные комплексы служат инструментом индикации и мониторинга состояния фитоценоза на ранних стадиях его поражения.
Численность эпифитных микроорганизмов, соотношение спороносных и неспоровых форм, наличие пигментированных бактерий являются критериями степени техногенного воздействия Назаровской ГРЭС на древостои.
Негативное состояние насаждений в результате нарушающих воздействий (дефолиация сибирским шелкопрядом) диагностируется высокой частотой встречаемости комплекса офиостомовых грибов, ассоциированных с насекомыми-ксилофагами.
Структурные изменения в количественном соотношении различных групп эпифитных микроорганизмов на здоровой и пораженной инфекцией хвое связаны не только с фенофазой роста и развития деревьев, но и с их фитонцидной активностью.
Список литературы Микробные комплексы филлосферы как индикаторы состояния древостоев-эдификаторов
- Афанасова Е.Н., Пашенова Н.В. Взаимоот-ношения офиостомовых грибов, переноси-мых насекомыми-ксилофагами, между собой и другими микромицетами хвойных пород Сибири//Микология и фитопатология. -2005. -Т. 39. -№ 2. -С. 62-65.
- Горленко М.В. Функциональное биоразно-образие почвенных микроорганизмов: под-ходы к оценке//Перспективы развития почвенной биологии. -М.: Наука, 2001. -С. 228-234.
- Гузев В.С. Экологическая оценка антропо-генных воздействий на микробную систему почвы: автореф. дис.. д-ра биол. наук. -М.: Изд-во МГУ, 1988. -38 с.
- Добровольская Т.Г. Структура бактериаль-ных сообществ почв. -М.: Академкнига, 2002. -282 с.
- Добровольская Т.Г., Скворцова И.Н., Лысак Л.В. Методы выделения и идентификации поч-венных бактерий. -М.: Изд-во МГУ, 1989. -71 с.
- Красильников Н.А. Определитель бактерий и актиномицетов. -М.; Л.: Изд-во АН СССР, 1949. -829 с.
- Никитина З.И. Микробиологический мони-торинг наземных экосистем. -Новосибирск: Наука, 1991. -219 с.
- Определитель бактерий Берджи/под ред. Дж. Хоулта, Н. Крига, П. Снита и др. -М.: Мир, 1997. -Т. 1, 2. -800 с.
- Пашенова Н.В., Ветрова В.П., Матренина Р.М. и др. Офиостомовые грибы в ходах лиственничного короеда//Лесоведение. -1995. -№ 6. -С. 61-67.
- Скворцова И.Н. Идентификация почвенных бактерий. -М.: Изд-во МГУ, 1983. -63 с.
- Сорокин Н.Д. Микробиологический монито-ринг лесных экосистем Сибири при различ-ных антропогенных воздействиях//Успехи современной биологии. -1993. -Т. 113, вып. 4. -С. 137-169.
- Сорокин Н.Д. Микробиологическая диагно-стика лесорастительного состояния почв Средней Сибири. -Новосибирск: Изд-во СОРАН, 2009. -221 с.
