Микробные сообщества подзолистых почв на вырубках среднетаежных еловых лесов

Автор: Виноградова Юлия Алексеевна, Лаптева Елена Морисовна, Перминова Евгения Максимовна, Анисимов Сергей Сергеевич, Новаковский Александр Борисович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология и география почв
Статья в выпуске: 5 т.16, 2014 года.
Бесплатный доступ
В данной статье рассмотрено влияние сплошнолесосечных рубок на изменение состояния и структуры комплекса почвенных микроорганизмов. Показано, что на первых стадиях восстановления растительного покрова на вырубках, в связи с ухудшением экологического состояния подзолистых почв, происходит снижение численности и биомассы микроорганизмов, численности эколого-трофических групп микроорганизмов, функциональной активности микробиоты.
Подзолистые почвы, вырубки, микробное сообщество, мультисубстратное тестирование
Короткий адрес: https://sciup.org/148205695
IDR: 148205695 | УДК: 631.461:631.445.2:630*187:582.475(1-924.82)
Microbe communities in podzolic soils at logged spruce forest areas of the middle taiga zone
This paper is devoted to the influence of clear-cut forest areas on the health status and structure of soil microorganisms. The initial vegetation cover restoration stages at clear cuttings are characterized with a declining number and biomass of microorganisms, fewer ecologic-trophic groups of microorganisms, and a less functional activity of microbiota, due to bad ecological status of podzolic soils.
Текст научной статьи Микробные сообщества подзолистых почв на вырубках среднетаежных еловых лесов
Почвенное микробное сообщество – наиболее мобильный блок почвенной биоты, активно реагирующий на любые изменения экологических условий как природного, так и техногенного характера. Основные компоненты комплекса почвенной микробиоты (водоросли, микроскопические грибы, актиномицеты, бактерии) успешно используются в качестве тест-систем для оценки экологического состояния как непосредственно почв, так и в целом наземных экосистем. В последние годы для этих целей широкое применение получил метод мультисубстратного тестирования (МТС) [3]. Изначально он был предложен для идентификации патогенных микроорганизмов [17], но в настоящее время успешно используется для диагностики микробных сообществ природных экосистем [19, 20, 18]. Его основным преимуществом, по сравнению с классическими микробиологическими методами (определение численности и соотношения эколого-трофических групп микроорганизмов, численности и биомассы бактерий, дрожжей и микроскопических грибов), является возможность оценки функционального состояния (функциональной активности, или функционального разнообразия) микробного сообщества, населяющего различные горизонты почвы [16, 2, 4].
В Республике Коми одним из значимых антропогенных факторов, приводящих к масштабным нарушениям природных наземных экосистем, следует отнести промышленные рубки. В результате смены растительности на вырубках
происходят существенные изменения растительности, меняются микроклиматические параметры формирующихся биотопов, гидрологический режим почв и их физико-химические свойства. Все это приводит к определенным структурным перестройкам комплекса почвенных беспозвоночных животных, а также почвенных микробных сообществ [7].
Цель данной работы заключалась в выявлении основных закономерностей изменения структуры и функционального состояния микробных сообществ подзолистых почв на первых этапах естественного лесовозобновления после сплошнолесосечных рубок среднетаежных еловых лесов.
ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили на территории Республики Коми, на стационарных участках, заложенных в Усть-Куломском р-не для проведения комплексных исследований на разновозрастных вырубках среднетаежных еловых лесов [10]. Район исследования расположен в Южно-Тиманском округе Вычегодской провинции среднетаежной подзоны типичных подзолистых почв [1]. В качестве объектов исследования выбраны подзолистые суглинистые почвы коренного ельника чернично-зеленомошного (участок ПП1) и лиственно-хвойного насаждения, сформировавшегося после сплошнолесосечной рубки в 2001/2002 гг. (участок ПП2).
Детальная характеристика растительного покрова и почв ключевых участков представлена в работах [10, 5, 6]. Здесь отметим следующее. Участок ПП1 заложен в ельнике черничнозеленомошным. Состав древостоя 6Е4Пх+Б, возраст 60-230 лет. Отмечен подрост ели и пихты. Травяно-кустарничковый ярус из Vaccinium myrtillus, Maianthemum bifolium, Linnaea borealis, Dryopteris (sp.) и др. В напочвенном покрове Pleurozium shreberi, Hylocomium splendens, редко Polytrichum commune, Sphagnum girgensohnii, проективное покрытие 80-85%. В почвенном покрове доминируют подзолистые текстурнодифференцированные почвы.
Участок ПП2 характеризует вырубку, где в зимний период 2001/2002 гг. были проведены сплошнолесосечные рубки главного пользования. В настоящее время на участке идет активное возобновление древесной растительности. На вырубке выражен интенсивный рост березы и рябины. В формирующемся фитоценозе значительную роль выполняет подрост хвойных растений, сохранившийся во время лесозаготовки. Следует отметить, что при проведении рубки напочвенный покров не был нарушен. Однако в его структуре в настоящее время происходят определенные изменения - почти исчезли Vaccinium myrtillus, Trientalis europaea, Maianthemum bifolium, Linnaea borealis и появились V. vitis-idaea и Carex sp. [10]. В напочвенном покрове доминируют политрихо-вые мхи. Основным компонентом в структуре почвенного покрова являются подзолистые текстурно-дифференцированные с микропрофилем подзола почвы.
Для оценки состояния микробных сообществ подзолистых почв отбор проб проводили в августе 2013 г. из слоя живых мхов (СЖМ), подстилочно-торфяного (подгоризонты О1, О2, О3) и подзолистого горизонта (EL(hg)). Для анализа использовали свежие образцы. Численность основных физиологических групп микроорганизмов почвы оценивали методом посева почвенных суспензий на твердые питательные среды [11]. Общее количество гетеротрофных бактерий учитывали на мясо-пептонном агаре (МПА), олигонитрофилов - на среде Эшби, олиготрофов - на голодном агаре, микроорганизмов, использующих минеральные формы азота, - на крахмало-аммиачном агаре (КАА). Численность бактерий и спор грибов, длину мицелия микроскопических грибов и их биомассу определяли с использованием метода люминесцентной микроскопии [9].
Мультисубстратное тестирование выполняли в соответствии со стандартной методикой [8]. При проведении анализа оценивали спектры потребления субстратов (СПС) микробными сообществами. В работе использовали стандартные 96-луночные планшеты с 47 источниками органического углерода (сахара, спирты, соли органических кислот, аминокислоты, амины, амиды, нуклеозиды). Планшеты инкубировали в термостате в течение 3-х суток при температуре 28°С до появления визуально регистрируемой окраски ячеек. Изменение окраски субстратов вызвано развитием микроорганизмов, способных потреблять тест-субстрат, содержащийся в ячейке планшета, и восстанавливать бесцветный трифенил-тетразолий в бордово-красный формазан. После окончания инкубации осуществляли фотометрическое считывание оптической плотности ячеек при длине волны 510 нм на фотометре «Уни- план». На основании совокупности полученных данных рассчитывали коэффициенты биоразнообразия (индекс Шеннона, индекс выравненности Пиелу), рангового распределения потребления субстратов, стабильности сообщества (d), а также удельную метаболическую работу (W) и интегральный параметр (G) общего благополучия системы [4]. Для сравнения спектров потребляемых субстратов применяли дискриминантный и кластерный анализы (кластеризация - по Варду, мера расстояния - Манхеттеновское расстояние) [15].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Как показали проведенные исследования, для рассмотренных почв характерны общие закономерности профильного изменения численности эколого-трофических групп микроорганизмов (ЭТГМ). В основном микроорганизмы сосредоточены в слое живых мхов и подстилочно-торфяном горизонте (рис. 1). Максимальной численностью отличается верхняя часть лесной подстилки -подгоризонт О1. В слое живых мхов и в нижележащих подгоризонтах подстилочно-торфяного горизонта численность микроорганизмов на 2-4 порядка меньше. При переходе от органогенных горизонтов к минеральным наблюдается резкое снижение общего количества гетеротрофных бактерий, учитываемых на твердых питательных средах. Такое распределение микроорганизмов типично для почв таежных экосистем [12]. Оно характерно и для почвы целинного, ненарушенного ельника чернично-зеленомошного (ПП1), и для почвы хвойно-лиственного насаждения, сформировавшегося на вырубке 2001/2002 гг. (ПП2).
В структуре биомассы рассмотренных микробных сообществ основную роль играет мицелий микроскопических грибов (табл. 1). На его долю в почве участка ПП1 приходится 68,7-85,6% общей биомассы микроорганизмов. В направлении от органогенного горизонта к минеральному доля грибного мицелия в общей микробной биомассе снижается, но при этом происходит возрастание доли биомассы спор грибов - от 11,1 % в гор. О до 31,2% в гор. EL. Вклад в микробную биомассу прокариот незначителен. Максимальное ее количество (3,1% от общей микробной биомассы) приурочено к горизонту лесной подстилки.
Ранее проведенными исследованиями показано, что в биоклиматических условиях средней тайги на вырубках происходит временное переувлажнение почв, сопровождающееся активизацией процессов глееобразования, возрастанием кислотности верхней части почв [10], усилением агрессивности и миграционной способности гумусовых веществ [6], мобилизацией и сегрегацией соединений железа [21]. Максимально эти изменения выражены в почвах молодых вырубок (5-
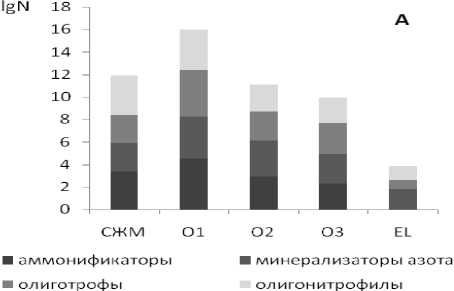
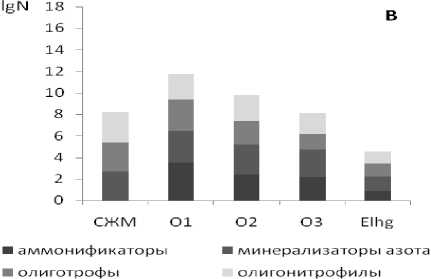
Рис. 1. Численность (lgN, где N – число жизнеспособных клеток в 1 г абсолютно-сухой почвы) и соотношение трофических групп микроорганизмов в почве коренного ельника чернично-зеленомошного (А) и хвойно-лиственного насаждения, сформировавшегося на вырубке 2001/2002 гг.
10 лет). Преобразование морфологических и физико-химических свойств подзолистых почв на вырубках нашло свое отражение в изменении параметров, характеризующих состояние почвенного микробного сообщества. Ухудшение экологических условий функционирования микроорганизмов на начальных этапах восстановления растительного покрова на вырубках в первую очередь проявилось в снижении численности и биомассы основных компонентов почвенного микробного комплекса (табл. 1). В лесной подстилке почвы участка ПП2, по сравнению с аналогичным горизонтом почвы коренного ельника (ПП2), в 6,5 раз снизилась численность и биомасса прокариот, в 1,8-1,9 – численность и биомасса спор микроскопических грибов, длина и биомасса грибного мицелия. В минеральном подзолистом горизонте количественные параметры структурной перестройки микробного сообщества выражены более резко. В гор. ELhg почвы вырубки, имеющем морфохроматические признаки оглеения в виде сизовато-серых пятен, численность и биомасса бактерий снижены практически в 12-12,5 раз, спор грибов – в 4,6-5 раза. В тоже время длина и биомасса мицелия микроскопических грибов осталась практически на том же уровне, что может быть обусловлено естественными причинами – изначально слабым распространением грибного мицелия в минеральную толщу подзолистых почв и преимущественным его концентрированием в органогенных горизонтах почв таежных экосистем. Данные люминесцентной микроскопии (табл. 1) хорошо согласуются с результатами оценки численности эколого-трофических групп микроорганизмов методом посева на твердые питательные среды (рис. 1). В почвах вырубки (ПП2) во всех исследованных образцах суммарная численность эколого-трофических групп микроорганизмов на 2-4 порядка ниже по сравнению с аналогичными горизонтами коренного ельника чернично-зеленомошного (ПП1).
Использование метода МСТ для характеристики функциональной активности почвенной микробиоты показало, что микробные сообщества рассмотренных почв близки по количеству потребляемых субстратов. В органогенных горизонтах из 47 субстратов микробиотой использовано 13-30 (ПП1) и 23-33 (ПП2) источника органического углерода. Микробное сообщество подзолистых горизонтов ассимилирует всего 19 субстратов. Широкий спектр потребляемых субстратов, охватывающий различные классы соединений, свидетельствует о достаточно высоком функциональном потенциале бактериальных сообществ, составляющих основу микробных комплексов в лесных подстилках ненарушенной подзолистой почвы (ПП1) и почвы «молодой» вырубки (ПП2).
Таблица 1. Численность и биомасса микроорганизмов в почвах ельника чернично-зеленомошного (ПП1) и хвойно-лиственного насаждения, сформировавшегося на вырубке 2001/2002 гг. (ПП2)
|
Участок |
Горизонт |
Численность |
Биомасса (мг/г почвы) |
||||
|
бактерии, млн кл./г |
споры грибов, млн кл./г |
мицелий грибов, м/г |
бактерии |
мицелий грибов |
споры грибов |
||
|
ПП1 |
О |
3680 |
27.7 |
556.4 |
0.078 |
2.17 |
0.28 |
|
EL |
5.0 |
5.0 |
28.78 |
0.0001 |
0.11 |
0.05 |
|
|
ПП2 |
O |
570 |
15.1 |
288.23 |
0.012 |
1.12 |
0.16 |
|
ELhg |
0.4 |
1.1 |
25.6 |
8.3∙10-6 |
0.10 |
0.01 |
|
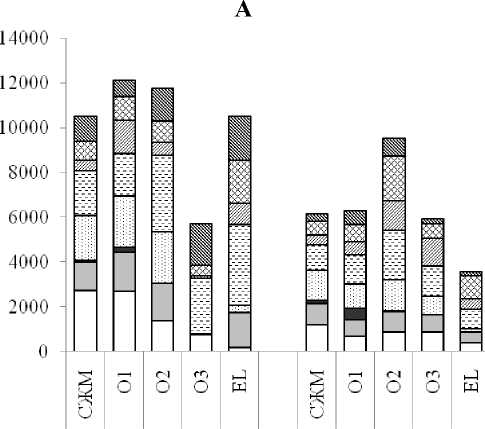
Почвы целинного леса и вырубки отличаются по интенсивности потребления субстратов (рис. 2А). На участке ПП1 микробные сообщества лес- ной подстилки и подзолистого горизонтов близки по их суммарному потреблению. Несколько выбивается в этом ряду подгоризонт О3 (подгори- зонт гумификации), где суммарная интенсивность ассимиляции источников органического углерода снижена практически в 2 раза по сравнению с верхними подгоризонтами подстилки и горизонтом EL. В пределах стратифицированной на подгоризонты лесной подстилки отмечается достаточно четкая тенденция снижения доли потребления пентоз, олигосахаридов и низкомолекулярных органических кислот (НМОК) по мере повышения степени разложения растительных остатков (СЖМ → О1→ О2 → О3) и повышения – активности использования аминокислот и азотсодержащих органических соединений (мочевина, креатинин, тимидин). Микробное сообщество и активизацией использования НМОК и полимерных (крахмал, декстран, твин-80) соединений.
По сравнению с почвой участка ПП1, на участке ПП2 практически во всех горизонтах снижается интенсивность потребления субстратов, за исключением подгоризонта О3 (рис. 2А). При этом, в отличие от почвы целинного леса, в нижней части лесной подстилки почвы вырубки (подгоризонты О2 и О3, где осуществляются процессы микробной трансформации растительного материала), наблюдается более активное потребление микробными сообществами НМОК при снижении использования полимерных соединений (рис. 2В). Активизация потребления НМОК обу- подзолистого горизонта характеризуется резким снижением потребления пентоз и олигосахаридов
□ I DTI ИТТТ DTV BY BYT 0VTT 0VTTT
словлена возрастанием их продуцирования в условиях повышения увлажнения почв [14].
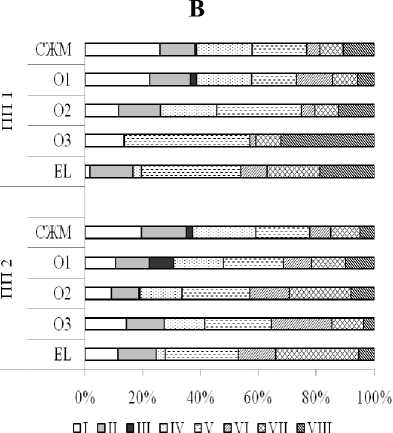
Рис. 2. Абсолютная (А; по вертикали: оптическая плотность; по горизонтали: горизонт почвы, участок) и относительная (В; по вертикали: горизонт почвы, участок; по горизонтали: процент) интенсивность потребления микробными сообществами различных групп субстратов: I – пентозы; II – гексозы; III – олигосахариды; IV – спирты; V – аминоксилоты; VI – низкомолекулярные органические кислоты; VII – полимеры; VIII – азотсодержащие органические соединения
Анализ спектров потребления субстратов почвенными микробными сообществами и их оценка на основе такого расчетного параметра как удельная метаболическая работа W, выявили четкие различия в рассматриваемом ряду почв (табл. 2). Максимальными показателями метаболической работы (1753-2581) характеризуются микробные сообщества почвы коренного ельника (ПП1). На вырубке это показатель снижен, в зависимости от генетического горизонта (подгоризонта), в 1,3-2,2 раза. Наиболее существенно выражено уменьшение величины удельной метаболической работы W в минеральном подзолистом горизонте вырубки: в почве целинного леса этот показатель составляет 2581, в почве вырубки – 1163.
Следует отметить, что микробные сообщества горизонтов лесных подстилок почв участков ПП1
и ПП2, особенно их верхних подгоризонтов О1 и О2, включая слой живого мха, незначительно отличаются по функциональному разнообразию. Значения индекса биоразнообразия Шеннона варьируют в узком пределе 4,3-4,7. Основные различия прослеживаются при сопоставлении нижней части лесных подстилок (гор. О3) и подзолистого горизонта. В почве целинного леса индекс Шеннона составил – 3,3-3,9, в почве вырубки – 4,0-4,3. Возрастание функционального разнообразия микробных сообществ в почвах вырубки обусловлено, скорее всего, расширением спектра поступающего органического материала в почву и на ее поверхность в результате смены пород – на вырубке формируется молодое лиственное насаждение из березы, осины, рябины с участием в подросте хвойных. В качественном отношении растительный опад, ежегодно включающийся в биологический круговорот, на вырубках имеет определенные отличия от опада коренного елового леса [5]. Опад лиственных насаждений характеризуется более высоким содержанием азота, кальция, магния, по сравнению с химическим составом опада в ельниках черничнозеленомошных, что может обусловливать расширение разнообразия почвенных микроорганизмов, несмотря на переувлажнение почвы вырубки.
Использование метода мультисубстратного тестирования позволяет оценить не только функциональную активность микробного сообщества, но и его экологическое состояние на основе расчета коэффициента рангового распределения потребления субстратов d как меры дестабилизации или возмущенности системы и интегрального параметра общего благополучия системы G как отношения биоразнообразия к нестабильности [3, 4]. В табл. 3 приведена разработанная М.В. Горленко и П.А. Кожевиным [3] шкала для характеристики состояния микробных сообществ по данным коэффициента рангового потребления субстратов d.
Таблица 2. Результаты оценки функционального состояния микробных сообществ по данным мультисубстратного тестирования
|
Площадка, горизонт |
Количество потребленных субстратов, N |
Коэффициент |
Интегральный параметр общего благополучия системы, G=H/d |
Удельная метаболическая работа, W |
|||
|
рангового распределения субстратов, d |
выравнен-ности, Е |
Шеннона, H |
|||||
|
ПП1 |
СЖМ |
30 |
0.7 |
0.9 |
4.6 |
6.2 |
1892 |
|
ПП1 |
О1 |
27 |
0.7 |
1.0 |
4.5 |
6.3 |
2044 |
|
ПП1 |
О2 |
25 |
0.5 |
0.9 |
4.3 |
9.0 |
2303 |
|
ПП1 |
О3 |
13 |
1.0 |
0.9 |
3.3 |
3.3 |
1753 |
|
ПП 1 |
EL |
19 |
0.4 |
0.9 |
3.9 |
9.3 |
2581 |
|
ПП2 |
СЖМ |
28 |
1.2 |
0.9 |
4.5 |
3.8 |
1197 |
|
ПП2 |
O1 |
33 |
1.2 |
0.9 |
4.7 |
3.9 |
1196 |
|
ПП2 |
O2 |
25 |
0.6 |
0.9 |
4.3 |
6.8 |
1803 |
|
ПП2 |
O3 |
23 |
0.8 |
1.0 |
4.3 |
5.1 |
1218 |
|
ПП2 |
ELhg |
19 |
2.6 |
0.9 |
4.0 |
1.6 |
1163 |
Таблица 3. Зависимость характера системы от значения d по [3]
|
Значение d |
Характеристика благополучия системы |
|
0,01-0,1 |
Благополучная избыточная система с максимальным запасом прочности |
|
0,1-0,4 |
Устойчивая стабильная система |
|
0,4-0,8 |
Система с истощенными ресурсами или система, находящаяся под обратимым воздействием какого-либо нарушающего фактора |
|
0,8-1,0 |
Кризисная дестабилизированная система |
|
Более 1,0 |
Необратимо нарушенная система |
Как показали проведенные расчеты (табл. 2), микробные сообщества практически всех горизонтов подзолистой почвы целинного леса, за исключением нижней части лесной подстилки (О3), отвечают системе с истощенными ресурсами или находящимися под обратимым воздействием какого-либо нарушающего фактора (d = 0,4-0,7). Подгоризонт гумификации О3 по показателям коэффициента рангового потребления субстратов d попадает в категорию кризисной дестабилизированной системы (d = 1,0). Почва целинного леса не имеет антропогенных нарушений, ее микробиота функционирует в режиме, соответствующем естественной природной динамике. Возможно, относительное неблагополучие в состоянии микробного сообщества почвы участка ПП1 обусловлено спецификой погодных условий года наблюдений. По данным ближайшей к стационарным участкам метеостанции «Помоздино», лет- ний период 2013 г. характеризовался аномально жаркими погодными условиями с недобором осадков. Иссушение лесных подстилок под пологом темнохвойного елового леса могло повлиять на ухудшение условий функционирования почвенной биоты. Однако в сравнении с почвой вырубки микробные сообщества почвы целинного леса оказались наиболее благополучными с точки зрения качества или «здоровья» почвы (табл. 2). Для почвы вырубки получены более высокие значения коэффициента рангового распределения d и наименьшие – интегрального параметра общего благополучия системы G (табл. 2). Расчетные данные позволяют рассматривать микробное сообщество почвы участка ПП2 в качестве необратимо нарушенной системы (слой живых мхов, подгоризонт О1, горизонт ELhg) и кризисной дестабилизированной системы (горизонт О3). Только лишь комплекс микроорганизмов средней части лесной подстилки – подгоризонта ферментации – отвечает системе с нарушенными ресурсами. Таким образом, ухудшение экологических условий на ранних стадиях послерубочной сукцессии за счет временного переувлажнения почв, развития в них процессов оглеения и повышения кислотности [10], нашло свое отражение в дестабилизации функционирования комплекса микроорганизмов в почвах вырубки.
Статистическая обработка спектров поглощения субстратов с использованием кластерного анализа выявила определенную специфичность микробных сообществ рассмотренных почв (рис. 3). Объединились в один кластер и оказались наиболее близки по функциональным характеристикам микробные сообщества органогенных горизонтов участков ПП1 и ПП2 и слоя живых мхов на участке ПП1. Что вполне объяснимо, т.к. на первых этапах восстановительной сукцессии растительного покрова на вырубках еловых лесов в подзоне средней тайги еще сохраняется сходство в строении горизонтов лесных подстилок почвы вырубки и целинного леса. Минеральные горизонты и слой живых мхов почвы вырубки образуют отдельный кластер.
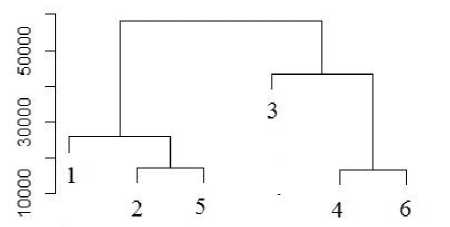
Рис. 3. Дендрограмма сходства в потреблении субстратов микробными сообществами коренного ельника (ПП1) и «молодой» вырубки (ПП2) (кластеризация – по Варду, мера расстояния – Манхеттенов-ское расстояние). Условные обозначения: слой живого мха – 1 (ПП1), 4 (ПП2); лесная подстилка – 2 (ПП1), 5 (ПП2); подзолистый горизонт – 3 (ПП1), 6 (ПП2).
ВЫВОДЫ
-
1) Установлены параметры, характеризующие микробные сообщества подзолистых почв северотаежных еловых лесов;
-
2) Показано, что в горизонтах лесных подстилок подзолистых почв вырубок происходит снижение численности, биомассы и функциональной активности микроорганизмов. В меньшей степени эти изменения затрагивают показатели длины мицелия и его биомассы в минеральных подзолистых горизонтах;
-
3) На ранних стадиях сукцессии после рубки еловых лесов происходит дестабилизация экологического состояния почвенных микробных комплексов.
Работа выполнена при финансовой поддержке проекта программы Президиума РАН №12-П-4-1065 «Взаимосвязь структурно-функциональной и пространственно-временной организации почвенной биоты с динамическими аспектами изменения подзолистых почв и почвенного органического вещества в процессе естественного восстановления таежных экосистем Европейского Северо-Востока после рубок главного пользования».
Список литературы Микробные сообщества подзолистых почв на вырубках среднетаежных еловых лесов
- Атлас почв Республики Коми/Под ред. Г.В. Добровольского, А.И. Таскаева, И.В. Забоевой. Сыктывкар, 2010. 356 с.
- Беленева И.А. Применение метода мультитсубстратного тестирования для характеристики морских сообществ обрастания металлов и сплавов/И.А. Беленева, У.В. Харченко, Ю.Л. Ковальчук//Биология моря. 2010. Т. 36. № 2. С. 145-150.
- Горленко М.В. Мультисубстратное тестирование природных микробных сообществ/М.В. Горленко, П.А. Кожевин. М.: МАКС Пресс, 2005. 88 c.
- Добровольская Т.Г. Реакция бактериальных сообществ лесной подстилки и почвы на внесение легкодоступных источников углерода и азота/Т.Г. Добровольская, М.В. Горленко, Н.В. Костина, А.Л. Степанов, С.А. Нестеров, А.В. Тиунов//Проблемы агрохимии и экологии. 2012. № 2. С. 36-41.
- Дымов А.А. Растительный опад в коренном ельнике и лиственно-хвойных насаждениях/А.А. Дымов, К.С. Бобкова, В.В. Тужилкина, Д.А. Ракина//Лесной журнал. 2012. № 3. С. 7-18.
- Дымов А.А. Изменение органического вещества таежных почв в процессе естественного лесовозобновления растительности после рубок (средняя тайга Республики Коми)/А.А. Дымов, Е.Ю. Милановский//Почвоведение. 2014. № 1. С. 1-9.
- Лаптева Е.М. Влияние рубок главного пользования на биотический комплекс среднетаежных подзолистых почв/Е.М. Лаптева, А.А. Колесникова, А.А. Таскаева, Т.Н. Конакова, А.А. Кудрин, Ю.А. Виноградова, Е.М. Перминова//Разнообразие лесных почв и биоразнообразие лесов: Матер. докл. V-ой Всеросс. науч. конф. по лесному почвоведению. Пущино, 2013. С. 175-177.
- Методика выполнения измерений интенсивности потребления тест-субстратов микробными сообществами почв и почвоподобных объектов фотометрическим методом: ФР.1.37.2010.08619., ПНД Ф Т 16.1.17-10. М., 2010.
- Методы почвенной микробиологии и биохимии/Под. ред. Д.Г. Звягинцева. М.: Изд-во МГУ, 1991. 304 с.
- Путеводитель научной почвенной экскурсии. Подзолистые суглинистые почвы разновозрастных вырубок (подзона средней тайги). Сыктывкар, 2007. 84 с.
- Теппер Е.З. Практикум по микробиологии/Теппер Е.З., Шильникова В.К., Переверзева Г.И. М.: Дрофа, 2004. 256 с.
- Регуляторная роль почвы в функционировании таежных экосистем/Отв. ред. Г.В. Добровольский. М.: Изд-во Наука, 2002. 364 с.
- Стольникова Е.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов Европейской территории России/Е.В. Стольникова, Н.Д. Ананьева, О.В. Чернова//Почвоведение. 2001. № 4. С. 479-494.
- Шамрикова Е.В. Кислотность почв таежной и тундровой зон Европейского Северо-Востока России. СПб.: Наука, 2013. 160 с
- Шипунов А.Б. Наглядная статистика. Используем R!/А.Б. Шипунов, E.М. Балдин, П.А. Волкова, А.И. Коробейников, С.А. Назарова, С.В. Петров, В.Г. Суфиянов. М.: ДМК Пресс, 2012. 298 с.
- Якушев А.В. Микробиологическая характеристика вермикомпостирования методом мультисубстратного тестирования/А.В. Якушев, Б.А. Бызов//Почвоведение. 2008. № 11. С. 1381-1387.
- Garland J.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization/J.L. Garland, A.L. Mills//Appl. Environ. Microbiol. 1991. Vol. 57. No 8. P. 2351-2359.
- Gömöryova E. Soil microbial community response to variation in vegetation and abitic environment in a temperate old-growth forest/E. Gömöryova, K. Ujhazy, M. Martinak., D. Gömöry//Applied Soil Ecology. 2013. Vol. 68. P. 10-19.
- Preston-Mafham, J. Analysis of microbial community functional diversity using sole-carbon-source utilization profiles -a critige/J. Preston-Mafha., L. Boddy, P. Randerson//Microbiology Ecology. 2002. Vol. 42. P. 1-14.
- Tian Y. Soil microbial communities associates with the rhizosphere of cucumber under different summer cover crops and residue management: А 4-year field experiment/Y. Tian, X. Zhang, J. Wang, L. Gao//Scientia Horticulturae. 2013. Vol. 150. P. 100-109.
- Lapteva E.M. Behavior of Iron Compounds in Soils of a Chronosiquence of Clear-Cuts/E.M. Lapteva, A.A. Dymov//Soil and Water Conservation under Changing Land Use. Lleida, Sspain, 2006. P. 231-234.
