Микроэлементы хвои в потомстве культур и естественных популяций ели финской

Автор: Рогозин Михаил Владимирович, Жекина Наталья Валерьевна, Комаров Вячеслав Сергеевич, Кувшинская Людмила Валентиновна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 3, 2014 года.
Бесплатный доступ
Проведен анализ содержания 10 микроэлементов (МЭ) в хвое 107 деревьев в 26-летнем возрасте в опытных культурах на суглинистых почвах в потомстве естественных популяций (Е) и культур (К). Вариация содержания МЭ составила в среднем 46.7% с колебаниями от 12 до 133%, и большинство рядов достоверно отличается от нормального распределения Гаусса-Лапласа. Обнаружено, что потомство К, у родителей которого больше признаков ели европейской, меньше накапливает МЭ, имеет меньшую зольность хвои и снижает ее при увеличении зольности почвы (r = -0.38±0.13). Хвоя потомства Е, у родителей которого больше признаков ели сибирской, наоборот, накапливает МЭ, имеет большую зольность и увеличивает ее с увеличением зольности почвы (r = 0.29±0.14).
Ель финская, популяции, потомство, хвоя, почва, микроэлементы
Короткий адрес: https://sciup.org/147204685
IDR: 147204685 | УДК: 582.47:
Microelements needles in progeny of the culture and natural populations Finnish spruce
The analysis of the content of 10 microelements (ME) in the needles of 107 trees in a 26-year-old cultures in loamy soils in the progeny of natural populations (N) and cultures (C). Variation ME averaged in the progeny N and C, respectively, 48% and 45.4%, and the majority of the series is significantly different from the normal distribution of the Gauss-Laplace. Found that offspring whose parents have more signs of european spruce, less accumulates ME has a lower ash content of needles and reduces it whereas soil increases their concentration (r = -0.38 ± 0.13). Needles of offspring N whose parents are more signs of Siberian spruce, on the contrary, gained ME the more, the more ash increased soil (r = 0,29 ± 0,14).
Текст научной статьи Микроэлементы хвои в потомстве культур и естественных популяций ели финской
Микроэлементы (МЭ) играют важную роль в процессах обмена и фотосинтеза. Изучение их концентраций в хвое часто предпринимают для выяснения отличий между популяциями при загрязнении среды. Обычно речь идет о превышениях концентраций МЭ в несколько раз и доказательство статистических различий не представляется сложным. Однако иногда различия оказываются малы при высокой вариативности данных, и требуется уже много образцов для процедуры такого доказательства.
С другой стороны, имеются работы для сосны обыкновенной, где различия в содержании микро-и макроэлементов характеризуют индивидуальные особенности клонов; предполагается, что эти различия могут быть связаны с быстротой роста и генетическими отличиями [Тараканов и др., 2007], что имеет важное диагностическое значение и позволит опознавать быстрорастущие потомства уже в раннем возрасте, а также отличать одну популяцию от другой.
Объекты и методы
Исследовано 26-летнее потомство ели финской (Picea х fennica (Regel) Кош.) разного происхождения: 52 дерева из 52 потомств, полученных из лесных культур Ф.А. Теплоухова, созданных в 1901-1916 гг. и 55 деревьев из 55 потомств из ес тественных популяций. Потомства выращивались в опытных культурах совместно и представляли собой часть большого опыта [Рогозин, Разин, 2012; Рогозин, 2013], заложенного на площади 11 га по испытанию более 15 тыс. растений, представляющих потомства от 525 деревьев ели. Опыт заложен в 1991 г. в кв. 41 Ильинского лесничества, координаты начала участка N 58°27'20.9" Е 55°50'17.8". Деревья, с которых брали образцы хвои, располагались по площади случайным образом. Изучалось содержание 10 микроэлементов (МЭ): Ni, Мн, Ti, Р, Си, Zn, Ba, Sr, Zr, Pb (в мг/кг), а также зольность хвои после ее сжигания (в %). Содержание хрома и ванадия (Cr, V) оказалось ниже чувствительности метода, и их концентрации не исследовали, хотя следы этих элементов и были зафиксированы. В связи с тем, что предполагалось влияние особенностей почвы на химический состав хвои, в радиусе 1.0 м от ствола дерева в трех местах делали прикопки на глубину 15-18 см и брали объединенный образец почвы. Анализы хвои проводили по методикам атомноабсорбционного анализа [Методы..., 1989], анализы почвы - по методикам почвено-химического [Аринушкина, 1962] и атомио-абсорбционного анализа [Методы..., 1989].
Результаты и их обсуждение
Ряды распределения частот зольности хвои и в том, и в другом потомствах были проверены на от-
линия от нормального распределения частот Гаусса-Лапласа по критерию «хи-квадрат» (у2). Данные были разбиты на 8 классов, и полученные оценки (у2 = 0.29-1.27) оказались значительно меньше критических у2 а95 = 6.25. что позволяет считать их распределение близким к нормальному. Критерий отношения дисперсий в сравниваемых выборках оказался равен F = 1.03 < Гу^ = 1.58. что позволило статистически корректно сравнивать средние значения с оценкой достоверности различий между ними по критерию 1.
Средняя зольность хвои в потомстве культур, называемых здесь сокращенно «потомство К», составила 5.31 ± 0.11%. а в потомстве естественных популяций, называемых «потомство Е» - 6.03 ± 0.10%. Различие составило 0.72% при 1 = 4.9 > t o.os = 1.98. Вариация значений зольности оказалась низкой и составила 12.4 и 14.5% соответственно, для потомства естественных популяций и потомства культур.
Следует отметить, что материнские насаждения имели существенные различия в морфологии по строению шишек и коры [Рогозин. 2013]: культуры имели больше признаков ели европейской, а естественные популяции - больше признаков ели сибирской. Повышенная зольность хвои у последних полностью согласуется со сведениями о том. что фенотипы с преобладанием признаков ели сибирской имеют более плотную древесину, чем фенотипы с преобладанием признаков ели европейской [Коренев. 2008]. Вероятно, это должно быть обусловлено физиологически, и повышенное накопление «зольных» элементов в хвое влияет на повышение плотности древесины.
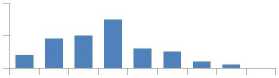
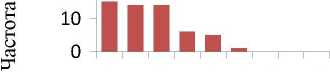
Общая картина соотношений между долями изученных микроэлементов в хвое оказалась одинакова в обоих потомствах. с полным совпадением рангов концентрации наиболее весомых МЭ: наибольший вклад вносят фосфор (Р) и марганец (Мп), далее идут Ba. Sr и Zn (рис. 1).
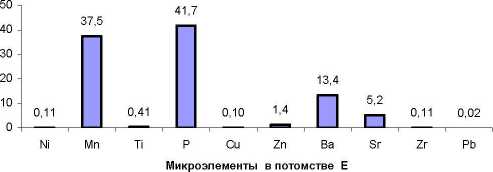
а
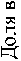
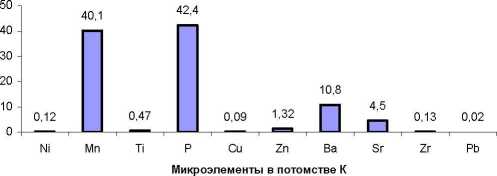
Рис. 1. Содержание 10 микроэлементов в хвое потомства естественных популяций (а) и в потомстве лесных культур (б) ели финской
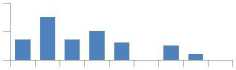
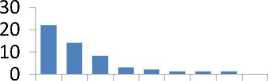
Для общего представления о различиях в накапливании МЭ в хвое потомств разного происхождения мы использовали вначале самое простое сравнение, а именно, разность средних значений МЭ в потомствах Е и К. принимая за 100% их со держание в потомстве Е: (Е - К) / Е 8 100. В результате оказалось, что в потомстве естественных популяций содержание семи МЭ оказалось выше, а трех - ниже, чем в потомстве К (рис. 2).
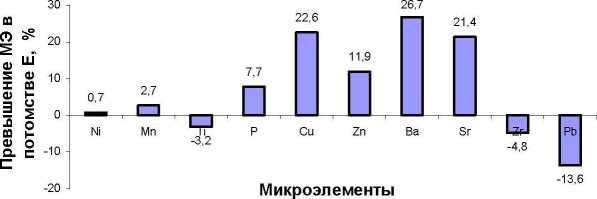
Рис. 2. Превышения в содержании микроэлементов в потомстве из естественных популяций (потомство Е) в сравнении с потомством из культур
Далее необходимо было определить статистическую достоверность полученных превышений, и здесь возникли определенные сложности, связанные с правомерностью использования стандартных процедур. Обычно используют следующие расчеты: определяют стандартное отклонение, ошибку среднего значения, критерий различия между средними (1) и сравнение его со стандартным (1 0.99; 10.95 или 1 о go)- Однако сама эта процедура допускается в строго определенном случае, а именно, при наличии двух условий.
Первое условие состоит в том, что дисперсии в сравниваемых выборках должны быть равны и отсутствие достоверных различий между ними доказывают с помощью критерия Фишера F в виде отношения дисперсий [Айвазян, Мхитарян, 1998].
Второе условие заключается в распределении частот, достоверно не отличающемся от нормального. Если распределение иное и имеет, например, асимметрию, то требуется преобразование исходных данных в иную шкалу, в которой распределение уже не будет отличаться от нормального. Отличие от нормального распределения оценивают с помощью критерия «хи-квадрат» (%2). Если же приблизить распределение к нормальному не получается, то проводят сравнение не просто средних значений, а сравнивают частоты у сравниваемых распределений по классам, причем по тому же критерию %2 Косвенно пригодность выборок для сравнения без преобразований часто определяют по значениям коэффициентов вариации (CW), и если они менее 30%, то распределения обычно мало отличаются от нормального и тогда обе выборки пригодны для сравнения без преобразования данных в иную шкалу.
В нашем случае вариация превысила уровень в 30% в потомстве Е в девяти, а в потомстве К - в восьми случаях из 10, причем у свинца вариация достигла 89-133%, что объясняется большой частотой данных этого элемента вблизи нулевых значений. В среднем вариация содержания МЭ в потомстве Е составила 48%, а в потомстве К - 45.4% (таблица).
По первому условию сравнения выборок (равенство дисперсий) в исходных выборках Е и К дисперсии оказались равны (достоверные отличия отсутствовали), но в преобразованных через логарифмы данных в одном случае (по меди) различия в дисперсиях были и отношения дисперсий превысили критическое значение (F = 1.77 > F0.95 = 1.58).
По второму условию исходные данные имели распределения частот, значимо отличающиеся от нормального, в 7 рядах из 10 в каждом потомстве (таблица). Поэтому мы применили преобразование исходных данных по всем МЭ через натуральный логарифм (In), после чего провели их разбиение на 6-8 классов. Кроме обычно используемого для оценки различия средних критерия 1, мы применили вышеупомянутый более тщательный метод сравнения выборок - по частотам в каждом из 6-8 классов по кри терию ^. Вначале мы провели сравнение фактических и теоретических частот этих классов для нормальных распределений в рядах с исходными данными ^ исх), а затем в рядах с логарифмированными данными (%2 In). Полученные изменения в ^оказались разнонаправленными - критерий как снижался, так и повышался, т.е. характер рядов в плане их приближения к нормальному распределению не был однозначен. В потомстве Е в 5 случаях критерий снизился и в 5 - повысился, а в потомстве К его повышение произошло только в двух случаях. В среднем значения критерия в потомстве Е снизились от ^ исх = 11.7 до %2 In =10.4, а в потомстве К - от ^ исх = 23.0 до %2 In =13.6. Положительный результат от логарифмирования данных в целом есть, и если исходные ряды распределения отличались от нормального в семи случаях, то логарифмированные - уже только в пяти по обоим потомствам. Особенно важным такое преобразование данных оказалось для свинца, и для него удалось доказать различия в распределении частот в 8 классах, хотя критерий различия средних 1 = 0.52 был недостоверен (таблица).
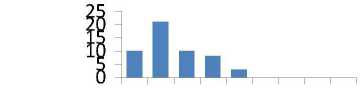
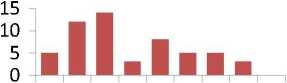
Общая картина распределений частот весьма разнообразна и наблюдается повышенная частота в самых разных классах; различия между средними значениями и частотами в классах распределений из естественных популяций и культур имеют марганец, титан, фосфор, медь, барий, стронций, свинец, т.е. семь элементов из десяти (рис. 3).
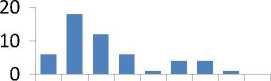
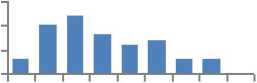
Различия в средних значениях и в распределении частот МЭ по классам между выборками из естественных популяций и культур по трем элементам из десяти - никелю, цинку и цирконию доказать не удалось (таблица), хотя внешне ряды и отличаются (рис. 4).
Кроме отличий в содержании МЭ в потомствах разного происхождения, важно было выяснить характер их накопления в хвое в зависимости от их содержания в почве. Разумеется, мы ожидали, что чем больше МЭ в почве, тем больше их будет накапливать и хвоя. Суммарная концентрация десяти МЭ в хвое оказалась тесно связана с зольностью хвои как в потомстве из культур (г = 0.64±0.09), так и в потомстве из естественных насаждений (г = 0.82 ± 0.04). Поэтому мы попытались далее определить, существует ли еще и связь между зольностью почвы и зольностью хвои.
Однако здесь все оказалось не так просто. В потомстве естественных ценозов эта связь получилась, как и ожидалось, положительной (г = 0.29 ± 0.14), но в потомстве из культур она сменилась на отрицательную (г = -0.38 ± 0.13), Обе связи достоверны и превышают уровень 1 о,9о = 1-7. Отсюда следует, что потомства явно и существенно отличаются физиологическими реакциями: при повышении зольности почвы потомства из культур снижают, а потомства из естественных популяций, наоборот, повышают зольность хвои (рис. 5).
Содержание микроэлементов в хвое ели в потомстве естественных популяций (Е) и в потомстве культур (К) с оценкой рядов распределения в исходной шкале данных (%2 исх) и в шкале, преобразованной через логарифмы значений (/2 In) с оценкой различий между ними по выборкам из 55 (потомство Е) и 52 значений (потомство К)
|
Показатели и статистики |
Микроэлементы |
В среднем |
Критические значения для ^ = 0.95 |
||||||||||
|
Ni |
Мн |
Ti |
Р |
Си |
Zn |
Ва |
Sr |
Zr |
Pb |
||||
|
Потомство Е, |
X, , мг/кг |
1.47 |
487.9 |
5.39 |
543.4 |
1.33 |
17.7 |
174.5 |
67.4 |
1.46 |
0.22 |
- |
|
|
cv,% |
40.0 |
39.9 |
48.4 |
14.1 |
36.8 |
48.8 |
37.1 |
37.1 |
44.2 |
133.2 |
48.0 |
- |
|
|
6 классов |
%2 исх |
12.5* |
16.7* |
12.2* |
5.6 |
13.8* |
1.65 |
5.95 |
13.4* |
11.3* |
24.0* |
11.7 |
7.81 |
|
частот |
Х21п |
6.26 |
17.7* |
19.9* |
12.2* |
17.8* |
4.61 |
5.29 |
4.58 |
2.89 |
12.5* |
10.4 |
7.81 |
|
Потомство К, |
Х2, мг/кг |
1.46 |
474.8 |
5.56 |
501.5 |
1.03 |
15.6 |
127.9 |
53.0 |
1.53 |
0.25 |
- |
- |
|
cv,% |
41.0 |
27.8 |
32.7 |
14.9 |
61.8 |
70.5 |
37.8 |
40.4 |
37.3 |
89.5 |
45.4 |
- |
|
|
6 классов |
%2 исх |
12.6* |
15.7* |
45.3* |
2.24 |
2.17 |
49.0* |
11.9* |
19.4* |
7.09 |
64.8* |
23.0 |
7.81 |
|
частот |
Х21п |
10.3* |
40.9* |
14.3* |
0.95 |
12.3* |
0.48 |
2.50 |
3.17 |
4.54 |
46.7* |
13.6 |
7.81 |
|
средними, % |
0.7 |
2.7 |
-3.2 |
7.7 |
22.6 |
11.9 |
26.7 |
21.4 |
-4.8 |
-13.6 |
7.21 |
||
|
Различия |
средними (1) |
0.08 |
0.40 |
0.39 |
2.86* |
2.75* |
1.11 |
4.20* |
3.20* |
0.59 |
0.52 |
1.61 |
1.98 |
|
дисперсиями (F) |
1.02 |
1.47 |
1.43 |
1.02 |
1.30 |
1.27 |
1.34 |
1.17 |
1.13 |
1.33 |
1.25 |
1.58 |
|
|
ИСХОДНЫМИ |
рядов по их частотам в 7 классах (%2) |
0.91 |
15.3* |
15.4* |
14.7* |
25.6* |
7.90 |
15.7* |
16.2* |
1.91 |
3.59 |
11.7 |
12.59 |
|
данными |
11.4 |
15.5* |
18.0* |
14.5* |
20.0* |
7.17 |
14.7* |
15.9* |
4.15 |
5.46 |
12.7 |
14.07 |
|
|
рядов по их частотам в 8 классах (%2) |
|||||||||||||
|
средними (1) |
0.17 |
0.38 |
1.16 |
2.80* |
3.69* |
1.74 |
4.45* |
3.41* |
0.93 |
1.71 |
2.04 |
||
|
Различия |
дисперсиями (F) |
1.05 |
1.43 |
1.46 |
1.01 |
1.77* |
1.16 |
1.04 |
1.03 |
1.20 |
1.15 |
1.23 |
1.58 |
|
между логарифмами |
рядов по их частотам в 7 классах (%2) |
4.63 |
18.2* |
15.7* |
12.7* |
19.1* |
7.09 |
22.7* |
9.92 |
3.30 |
9.49 |
12.2 |
12.59 |
|
данных |
рядов по их частотам в 8 классах (%2) |
1.83 |
17.7* |
18.6* |
15.3* |
23.7* |
8.02 |
20.2* |
16.2* |
4.25 |
17.5* |
14.3 |
14.07 |
|
* значение критерия превышает его стандартное критическое значение для р = 0.95 | |
|||||||||||||
Такие же направления корреляций, но несколько ослабленные, обнаружены при выяснении влияния зольности почвы на сумму всех 10 МЭ в хвое: в потомстве К она оказалась отрицательной (г = -0.20 ± 0.14), а в потомстве Е, наоборот, положительной (г = 0.23 ±0.13).
о
О
О
О
О
О
О
|
123456789 |
|
|
123456789 |
|
|
123456789 |
|
|
123456789 |
|
|
123456789 |
|
|
123456789 |
|
о
|
Мп* |
|
|
123456789 |
О
О
О
Ti*
р**
Си**
Ва**
О

Sr**
О

Pbh*
Классы ряда распределения
Рис. 3. Ряды распределения частот значений содержания микроэлементов в хвое в потомстве естественных популяций (слева) и в потомстве из культур (справа):
* достоверные различия в частотах в классах рядов распределения; ** достоверные различия в средних и в частотах в классах рядов распределения
Таким образом, морфологические отличия це-нопопуляций из культур Теплоухова. имевших больше признаков от ели европейской, в сравнении с таковыми в естественных популяциях, где деревья имели больше признаков ели сибирской [Рогозин. Разин. 2012; Рогозин. 2013]. имеют свое
«химическое» продолжение. Хвоя потомства из культур имеет меньшую зольность и достоверные отличия по частотам в классах рядов распределения значений элементов Мп. Ti. РЬ. она имеет достоверно меньшую зольность хвои и она меньше накапливает Р. Си. Ba. Sr в сравнении с хвоей в потомстве из естественных популяций.
20 ■
0 - ■ ■ ■ ----
Ni
20 -
Zn
О
Классы ряда распределения
Рис. 4. Ряды распределения частот значений содержания микроэлементов в хвое в потомстве естественных популяций (слева) и в потомстве из культур (справа), не имеющие достоверных различий в средних значениях и в классах рядов распределения
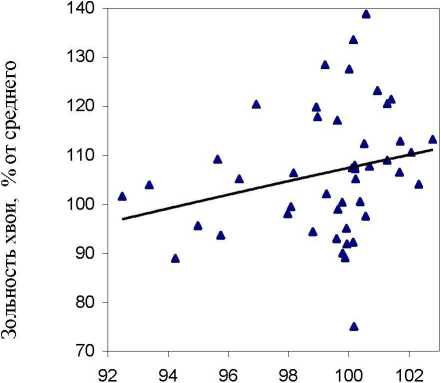
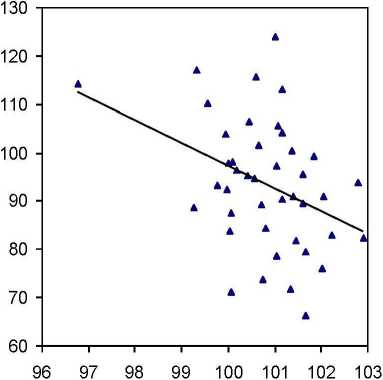
Зольность почвы вблизи растения. % от среднего
Рис. 5. Влияние зольности почвы вблизи растения на зольность его хвои в потомстве ели из естественных насаждений (слева) и в потомстве из культур (справа)
Заключение
Обнаружены достоверные отличия в химическом составе хвои по множеству микроэлементов и разнонаправленная реакция растений на увеличение или уменьшение их содержания в почве, свидетельствующие о существенных отличиях в физиологии потомства ели из культур Теплоухова. у которых больше признаков ели европейской, в отличие от потомства из естественных популяций, у которых больше признаков ели сибирской. Эти от личия подтверждают различия между популяциями. выявленные ранее на морфологическом уровне.
Работа выполнена за счет финансовой поддержки со стороны Министерства образования и науки России в рамках базовой части.
Список литературы Микроэлементы хвои в потомстве культур и естественных популяций ели финской
- Айвазян С.А., Мхитарян В.С. Прикладная статистика и основы эконометрики. М.: ЮНИТИ, 1998. 1024 с
- Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1962. 491 с
- Коренев И.А. Продуктивность ели в связи с морфологической изменчивостью вида в подзоне южной тайги: дис.... канд. с.х. наук. Кострома, 2008. 125 с
- Методы анализа минерального вещества, применяющиеся в центральной лаборатории ПО «Уралгеология». Свердловск, 1989. 81 с
- Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. Пермь, 2012. 210 с
- Рогозин М.В. Изменение параметров ценопопуляций Ртш sylvestris L. и Ргевй х fennica (Regel) Кот. в онтогенезе при искусственном и естественном отборе: автореф. дис.... д-ра с.х. наук. Пермь, 2013. 47 с
- Тараканов В.В. и др. Элементный состав хвои в разных клонах сосны обыкновенной//Лесоведение. 2007. № 1. С. 28-35
