Микроспорогенез у сортов Pyrus, Malus, Cerasus, Microcerasus и их гибридов

Автор: Яндовка Л.Ф., Папихин Р.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетические основы и приемы селекции
Статья в выпуске: 1 т.47, 2012 года.
Бесплатный доступ
Изучены особенности микроспорогенеза у видов Pyrus communis, Malus domestica, Cerasus vulgaris, Microcerasus tomentosa, Cerasus avium, а также у их межвидовых и межродовых гибридов. Описаны типичные и нетипичные нарушения микроспорогенеза. Первые были обнаружены у всех изученных растений, вторые - в основном у гибридов. Нетипичные нарушения становились причиной низкой фертильности пыльцевых зерен.
Микроспорогенез, фертильность и жизнеспособность пыльцы
Короткий адрес: https://sciup.org/142133272
IDR: 142133272 | УДК: 634:631.522/.524:631.527.5:576.354.4
Microsporogenesis in the Pyrus, Malus, Cerasus, Microcerasus varieties and their hybrids
The features of microsporogenesis were studied in Pyrus communis, Malus domestica, Cerasus vulgaris, Microcerasus tomentosa, Cerasus avium species and their interspecific and intergeneric hybrids. The typical and non-typical abnormalities in microsporogenesis were described. The typical - were found in all studied plants, the non-typical - in hybrids, as rule. The non-typical abnormalities caused the low fertility of pollen grains.
Текст научной статьи Микроспорогенез у сортов Pyrus, Malus, Cerasus, Microcerasus и их гибридов
Создание новых сортов, которые объединяют в себе хозяйственно ценные признаки культурных растений, относящихся к разным видам и родам, представляет особый интерес для селекционеров. Многим отечественным исследователям удалось преодолеть нескрещиваемость видов и получить плодовитые отдаленные гибриды (1-5). Наибольшую трудность при селекционном использовании межвидовых и межродовых гибридов представляет стерильность их пыльцы. Для разработки эффективных методов преодоления стерильности при отдаленной гибридизации растений необходимо знать особенности формирования генеративных органов, опыления, оплодотворения и развития семезачатка.
Формирование генеративных органов характеризуется наличием критических периодов (6-11). Термин «критические периоды» впервые был введен П.И. Броуновым в 1897 году (12). В развитии пыльника предложено выделить три периода: премейотический, мейотический и пост-мейотический (13). Эта периодизация была использована при разработке общих положений теории критических периодов в онтогенезе. Наиболее важный этап в развитии пыльника — микроспорогенез (мейотический период). Существуют работы по изучению процессов микроспорогенеза и развития пыльцевого зерна у родов Pyrus , Malus , Cerasus и Microcerasus. Типичные нарушения у сортов вишни, груши и яблони описаны многими авторами (14-19). Также имеются данные о фертильности и жизнеспособности пыльцы у представителей этих родов плодовых растений (20, 21).
Целью нашей работы стало изучение особенностей микроспорогенеза у сортов яблони домашней, груши обыкновенной, вишни обыкновенной, вишни войлочной и черешни, а также у их межвидовых и межродовых гибридов в условиях Тамбовской области.
Методика . Исследования проводили в 2003-2008 годах. Объектами служили растения яблони домашней ( Malus domestica Borkh.) — сорт Пепин шафранный [Ренет орлеанский х (Пепин английский х китайка)] селекции И.В. Мичурина; груши обыкновенной ( Pyrus communis ) — сорт Тонковетка народной селекции; яблоне-грушевых гибридов — № 818 (Malus baccata х смесь пыльцы южных сортов груши) селекции Т.А. Горшковой, № 839/67 (Китайка х Деканка зимняя + Кюре) селекции Г.А. Курсакова; груше-яблоневого гибрида — № 01 (Тонковетка х смесь пыльцы яблони) селекции С.Ф. Черненко; вишни обыкновенной ( Cerasus vulgaris ) — сорта Владимирская народной селекции, Интенсивная (Алый Парус х Долорес);
черешни ( Cerasus avium ) — сорт Итальянка (Слава Жукова х Бигарро); вишне-черешневых гибридов — сорт Фея (Коралл х Премьера); вишнечеремуховых гибридов — сорт Харитоновская (Жуковская х Алмаз) селекции ВНИИГиСПР им. И.В. Мичурина; вишни войлочной ( Microcerasus tomentosa ) из насаждений Всероссийского НИИ генетики и селекции плодовых растений им. И.В. Мичурина (ВНИИГиСПР им. И.В. Мичурина, г. Мичуринск, Тамбовская обл.).
Процесс мейоза исследовали на временных давленых препаратах с фиксацией в жидкости Карнуа (была приготовлена по сокращенной прописи), окрашенных ацетогематоксилином (22). Качество пыльцы оценивали с помощью окрашивания в ацетокармине, ее жизнеспособность — посредством проращивания на искусственной питательной среде (23). Препараты просматривали под микроскопом Carl Zeiss Jenamed (Германия), для фотодокументирования использовали цифровую камеру DCM-500 с программным обеспечением ScopePhoto.
Статистическую обработку данных проводили в программе Microsoft Excel.
Результаты . Процесс формирования микроспор у всех изученных растений в условиях Тамбовской области обычно начинался во II декаде апреля. В годы с высокими значениями эффективных температур мейоз в микроспороцитах проходил раньше — в первых числах апреля. В целом микроспорогенез у видов протекал сходно. В некоторых случаях наблюдалась асинхронность в делениях микроспороцитов в одном и том же цветке — от профазы I до телофазы II.
Изучение точек роста с помощью давленых препаратов показало, что в соматических клетках растений (2 n ) имеется 34 хромосомы у яблони домашней, груши обыкновенной, яблоне-грушевых и груше-яблоневых гибридов; 32 хромосомы — у вишни обыкновенной, вишне-черешневых и виш-не-черемуховых гибридов; 16 хромосом — у черешни и вишни войлочной.
У исследуемых растений обнаружены нарушения в распределении хроматина в I и II делениях мейоза, а также при цитокинезе в процессе образования диад и тетрад микроспор. У растений с генетически сбалансированными генотипами нарушений было меньше.
Вследствие генетических различий между скрещиваемыми геномами у гибридных растений часть хромосом не конъюгировала в профазе I деления. В результате наряду с бивалентами образовывались униваленты. У яблоне-грушевых гибридов число бивалентов колебалось от 12,0 до 17,0, унивалентов — от 0 до 10,0, тривалентов и тетравалентов не обнаружили. У вишне-черешневого гибрида соотношение бивалентов и унивалентов составляло 14,9:2,1; у вишне-черемухового гибрида — 15,0:2,0. Наибольшей гомологией хромосом у яблоне-грушевых форм характеризовался гибрид № 839/67 — 16,2:1,4. Значительное число образующихся бивалентов свидетельствует о высокой вероятности синтеза между гомологичными хромосомами яблони и груши. Наименьшая конъюгация хромосом наблюдалась у гибрида № 818 (13,8:3,7).
В хромосомных ассоциациях груше-яблоневого гибрида № 01 число бивалентов и унивалентов в среднем составляло соответственно 15,5 и 2,4. В некоторых микроспороцитах этого гибрида встречалось до 10 унивалентов. Наряду с аномальными клетками имелись клетки с правильной конъюгацией хромосом (17 бивалентов). Различить принадлежность хромосом гибридов к тому или иному исходному виду было невозможно из-за большого сходства. Трудно судить о том, какие хромосомы вступали в синапсис. Хромосомные ассоциации гибридов вишни обыкновенной харак- теризовались соотношением 15,6:0,7, у черешни оно составляло 15,8:0,2.
Микроспорогенез у всех исследованных видов и их гибридов в ряде случаев проходил с нарушениями. В зависимости от частоты нарушения разделили на две группы — типичные (часто встречающиеся) и нетипичные (редкие).
Типичные нарушения в разном сочетании были найдены у всех изученных растений (рис. 1). В метафазе I (M I) обнаруживались униваленты и биваленты, лежащие вне метафазных пластинок. В анафазе I (A I) при расхождении хромосом к полюсам наблюдали отставшие и забежавшие униваленты , асинхронное и численно неравномерное расхождение хромосом к полюсам. Отставшие при расхождении отдельные хромосомы или их группы в телофазе I (T I) формировали в материнских клетках микроспор различающиеся по размерам микроядра (обычно 1-2) либо подтягивались к полюсам. Наиболее частым нарушением в метафазе II (М II) была дезориентация и аномальное расположение хромосом за пределами метафазных пластинок, отмечалась также неравная величина групп хромосом. В анафазе II (А II) наблюдали задержку хромосом в экваториальной области, отставание и забегание вперед, выбросы хроматина за пределы веретена деления, неравномерное распределение хромосом. На стадии телофазы II (Т II) были обнаружены клетки с разным числом ядер (больше или меньше 4), клетки с микроядрами , образовавшимися в результате ошибок при распределении хромосом между дочерними ядрами на предыдущих стадиях мейоза. Достаточно часто вокруг микроядер обособлялся участок цитоплазмы и формировались микроспоры (как правило, меньшего размера). Происходили нарушения на стадии образования тетрад (формирование пентад, гексад, гептад, наличие одного или двух микроядер в одной из микроспор тетрады). В связи с нарушениями в процессе редукционного деления ядерный материал в конце мейоза распределялся неравномерно. Величина и форма микроспор в тетрадах часто различались.
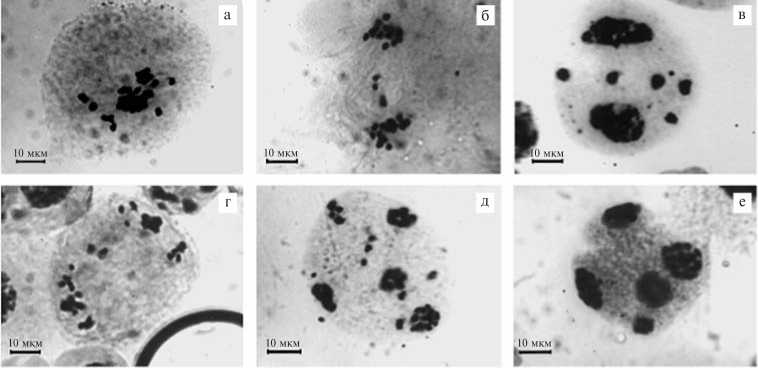
Рис. 1. Типичные нарушения микроспорогенеза у отдаленных гибридов плодовых растений: а — выбросы и отставание хромосом в метафазе I; б — отставание хромосом в анафазе I; в — образование микроядер в телофазе I; г — отставание и выбросы хроматид в анафазе II; д — формирование большего числа ядер в телофазе II; е — формирование пентады (окрашивание ацетогематоксилином, увеличение *1200).
Нетипичные нарушения мейоза наиболее часто обнаруживались у гибридов. Отмечался цитомиксис (перемещение отдельных хромосом или целых ядер из одного микроспороцита в другой или из одной микроспоры в 54
другую). Причина возникновения цитомиксиса, по мнению В .А. Поддуб-ной-Арнольди (19), — нарушение физиологического состояния ядра и цитоплазмы под влиянием гибридизации или неблагоприятных условий окружающей среды; автор указывает на патологическую сущность этого явления. В результате цитомиксиса образуются гаметы с недостаточным или избыточным числом хромосом и их фрагментов. Цитомиксис был обнаружен у гибридов вишни обыкновенной (П I, М II, стадия образования тетрад), черешни (П I, А II, Т II), яблоне-грушевого гибрида № 839/67 (М I), груше-яблоневого гибрида № 01 (A I). Частота клеток с цитомик-сисом у разных по происхождению растений различалась. У генетически сбалансированных генотипов, например у сорта Владимирская, цитомиксис не обнаружили, у гибрида № 839/67 его отмечали в 8,6, у гибридов вишни обыкновенной — в 3,6 % клеток.
Из других нетипичных нарушений следует отметить появление на стадии А I выбросов бивалентов за пределы веретена деления, образова ние мостов между не разошедшимися к полюсам хромосомами. У гибридных растений эти нарушения встречались чаще. На стадии Т I было выявлено наличие трех и более микроядер разной величины у вишни обыкновенной и яблоне-грушевого гибрида № 839/67. В анафазе II хро мосомные мосты и неравные по числу группы хромосом обнаруживали у черешни, вишни обыкновенной, яблоне-грушевых и груше-яблоневых гибридов. Большое число микроядер (до 8-9) на стадии Т II отмечали у многих растений. На стадии образования тетрад могло формироваться иное число микроспор: диады, триады (яблоне-грушевые и груше-яблоневые гибриды, вишня войлочная, вишня обыкновенная, черешня) и октады (виш-не-черешневые гибриды).
so. Частота нарушений на раз-
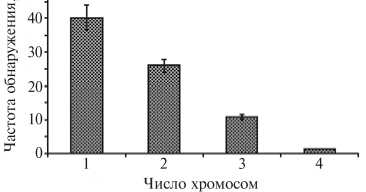
Рис. 2. Частота обнаружения хромосом вне экваториальной пластинки в метафазе I у яблоне-грушевого гибрида № 818.
ных стадиях мейоза у изученных видов и их гибридов оказалась неодинаковой. Наиболее высокой она была у гибридных растений (табл.). Характерная особенность метафазы первого деления мейоза — наличие одной-четырех хромосом, отстающих в прометафазном движении или выброшенных в цитоплазму за пределы метафазной пластинки. У вишни обыкновенной генетически сбаланси рованного сорта Владимирская частота выброшенных и отстающих хромосом составляла 17,7, частота нарушений в М I у вишне-черешневого гибрида Фея равнялась 41,4, у сорта Тонковетка груши обыкновенной — 6,4, у отдаленных межродовых гибридов яблони и груши — 39,6-89,5 %. Число хромосом вне экваториальной пластинки могло доходить до четырех (рис. 2). У гибрида № 818 число микроспороцитов с одной хромосомой вне экваториальной пластинки составляло 40,1, с двумя хромосомами — 26,0, с тремя — 10,7, с четырьмя — 1,2 %. У груше-яблоневого гибрида № 01 в М I наблюдали также разделение метафазной пластинки (16,9 %) на несколько групп (чаще на две). Подобное нарушение в процесе мейоза может служить предпосылкой образования несбалансированных гамет. У генетически сбалансированных растений подобного явления не обнаружено.
В А I, как и на предыдущих стадиях мейоза, частота клеток с нарушениями у гибридов была выше. Так, у вишни сорта Владимирская в АI
Частота хромосомных нарушений на разных стадиях микроспорогенеза у Pyrus communis , Malus domestica , Cerasus vulgaris, Microcerasus to-mentosa, Cerasus avium и их гибридов
При просмотре анафазных клеток первого деления у груше-яблоневого и яблоне-грушевого гибридов выяснилось, что большинство клеток (от 15,3 до 34,3 %) были аномальными из-за преждевременно разошедшихся хро мосом. Отставание хромосом в А I у гибридов также встречалось довольно часто (60-70 % от общего числа аномальных клеток). Обычно отставали 25 хромосом. Значительно реже наблюдали хромосомы, выброшенные в цитоплазму (10,8 %).
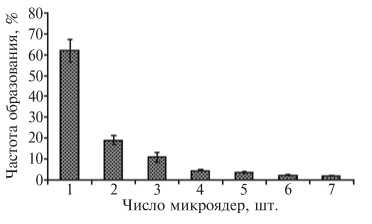
Рис. 3. Частота образования разного числа микроядр в телофазе I у яблоне-грушевого гибрида № 839/67.
Предпосылкой образования микроядер в Т I служили частые отставания и выбросы хромосом в цитоплазму на предшествующих стадиях мейоза. Микроядра в Т I у гибридов встречались чаще, чем у сбалансированных генотипов, их число могло достигать семи (рис. 3). Неодинаковую величину ядер зафиксировали только у груше-яб-лоневого гибрида № 01 и сортов вишни обыкновенной Фея и Ха-ритоновская; у черешни в двух клетках также была обнаружена неравная величина ядер.
Второе деление мейоза в ряде случаев тоже протекало с аномалиями. Материнские клетки микроспор в М II с группами, неравноценными по числу составляющих их хромосом, наиболее часто встречались у гибридных растений. В каждой группе число хроматид составляло 12-27. Выбросы хроматид в цитоплазму у гибридов обнаруживались в 21,3 % случаев. Клетки с хромосомными мостами встречались с частотой 0,6 % от общего числа нарушений.
Нарушения в анафазе II приводили к возникновению в телофазе II большого числа материнских клеток с микроядрами, образовавшимися из выброшенных или отставших хромосом. В Т II у генетически сбалансированных форм в аномальных мейоцитах обнаруживали не более двух микроядер, в то время как у гибридов — до 7-8. Выброшенные на начальных стадиях мейоза хроматиды в телофазе II в некоторых случаях обособлялись в группы и формировали дополнительные ядра. У видовых форм дополнительных ядер не наблюдали. У гибридных генотипов, например яблоне-грушевого гибрида № 818 и вишне-черешневого гибрида Фея, обнаружили до 8 ядер. У формы № 01, помимо четырехъядерных, образовывались трехъядерные (6,2 %) и двухъядерные (2,0 %) мейоциты. Такое соотношение клеток у гибрида № 01 указывает на генотипическую склонность этой формы к образованию нередуцированных гамет, что считается весьма ценным свойством для дальнейшего использования в селекции. Средний процент нарушений по всем стадиям мейоза у генетически сбалансированных растений был значительно меньше, чем у гибридных форм (см. табл.).
Следствием нарушений на разных стадиях мейоза у исследованных растений стало аномальное формирование и развитие тетрад. Размеры микроспор и ядер в клетках варьировали, нередко обнаруживались микронуклеусы и добавочные ядра, формирующиеся в телофазе второго деления. Частота нарушений на стадии образования тетрад у гибридов была выше, чем у сортов (см. табл.).
Показателем морфологической полноценности микроспор служит фертильность пыльцы. На морфологическую сформированность и жизне- способность пыльцевых зерен влияют разные факторы (24). Основной из них — происхождение растений. В наших опытах наибольшую фертильность пыльцы, определяемую ацетокарминовым методом, имели генетически сбалансированные генотипы. У вишни сорта Владимирская этот показатель составлял 74,5, у вишни войлочной — 89,3, у яблони сорта Пепин шафранный — 87,9, у груши сорта Тонковетка — 72,2 %. Доля морфологически сформированных пыльцевых зерен у гибридов была ниже: у сорта Харитоновская — 66,4, у сорта Фея — 65,2 %. У яблоне-грушевых гибридов № 839/67 и № 818 доля окрашенной пыльцы была ниже — 2,1 %, фертильность груше-яблоневого гибрида № 01 равнялась 16,1 %.
Жизнеспособность пыльцы, определяемая ее проращиванием на искусственной питательной среде, составляла у вишни сорта Владимирская 43,7, у вишни войлочной — 42,9, у яблони сорта Пепин шафранный — 87,9, у груши сорта Тонковетка — 72,2, у сорта Харитоновская — 9,9, у сорта Фея — 18,9 %. Пыльца межродовых гибридов семечковых растений прорастала плохо. Так, у формы № 01 ее жизнеспособность составляла 8,3, у № 839/67 — 6,9, пыльца гибрида № 818 не прорастала.
Таким образом, у растений видов Pyrus communis , Malus domestica , Cerasus vulgaris , Microcerasus tomentosa , Cerasus avium и их гибридов выявлены типичные и нетипичные нарушения микроспорогенеза. Типичные нарушения, которые имеются у всех изученных растений, связаны с распределением хроматина (неспособность большинства хромосом конъюгировать друг с другом, формирование унивалентов, отставания, забегания, выбросы хромосом за пределы веретена деления и неравномерное расхождение хромосом к полюсам, приводящее к образованию микроядер и нарушениям при образовании тетрад микроспор). Нетипичные нарушения мейоза наиболее часты у гибридов. Результатом таких нарушений становится образование гамет с числом хромосом, не соответствующим гаплоидному, и формирование пыльцы с пониженной фертильностью. Выявлены гибридные генотипы, способные формировать некоторое количество фертильных пыльцевых зерен. Они могут быть использованы в селекционной работе с целью объединения в потомстве хозяйственно ценных признаков растений, относящихся к разным видам и родам. Особенности протекания микроспорогенеза могут стать дополнительным признаком к общепринятым характеристикам представителей видов и их гибридов.
Л И Т Е Р А Т У Р А
-
1. Г о р ш к о в а Т.А. Яблоне-грушевые гибриды. Бюлл. научно-технической информации ЦГЛ им. И.В. Мичурина. Мичуринск, 1956, 1: 13-18.
-
2. П а п и х и н Р.В., М у р а т о в а С.А. Получение межродовых гибридов плодовых семечковых растений с применением метода культуры тканей и полиплоидизации in vitro. С.-х. биол., 2011, 5: 63-68.
-
3. К у р с а к о в Г.А. Отдаленная гибридизация плодовых растений. М., 1986.
-
4. М и ч у р и н И.В. Сочинения. Т. I-IV. М., 1948.
-
5. Р у д е н к о И.С. Отдаленная гибридизация и полиплоидия у плодовых растений. Кишинев, 1978.
-
6. Ж у к о в О.С., Х а р и т о н о в а Е.Н. Селекция вишни. М., 1988.
-
7. Б е р е з е н к о Н.П., Т р е т я к К.Д., 3 а в г о р о д н я я В.Г. Эмбриогенез и осыпание цветков и завязей у некоторых сортов вишни. Мат. Всес. совещания эмбриологов растений. Кишинев, 1986: 180.
-
8. К о з л о в а Н.А. Некоторые морфологические особенности развития генеративных органов и семян у вишни латвийской низкой ( Cerasus fruticosa ). II. Ботанический журнал, 1959, 45(1): 84-89.
-
9. С в е т л о в П.Г. Теория критических периодов и ее значение для понимания принципов действия среды на онтогенез. М.-Л., 1960: 263-285.
-
10. С к а з к и н Ф.Д. Критический период у растений по отношению к недостатку воды в почве. Л., 1971.
-
11. С п и ц ы н И.П. Генетика, цитология, эмбриология вишни. Экология. Тамбов, 1994.
-
12. Б р о у н о в П.И. Значение сельскохозяйственно-меорологических наблюдений и краткое руководство для проведения их. СПб, 1897.
-
13. К а м е л и н а О.П. Сравнительная эмбриология цветковых растений. Пыльник. Л., 1981, т. 1: 18-21.
-
14. Д е б и с к а е в а С.Ю. Фертильность пыльцы сортов черешни. В сб. науч. тр. СевероКавказского НИИ горного и предгорног о садоводства: Интенсификация садоводства. Нальчик, 1995, вып. 5: 90-93.
-
15. Н а з а р о в а М.Н., С и м о н о в а О.А. Мейоз при микроспорогенезе и качество пыльцы у вишни войлочной. I. В сб. науч. тр.: Отдаленная гибридизация и полиплоидия в селекции растений. Воронеж, 1989.
-
16. П а х о м о в а Н.П. Микроспорогенез межродового гибрида Malus baccata Borkh. х Pyrus communis L. Тр. ЦГЛ им. И.В. Мичурина (Мичуринск), 1971, т. 12: 210-221.
-
17. X а р и т о н о в а Е.Н. Цитологические исследования косточковых. Тр. ЦГЛ им. Мичурина. Мичуринск, 1971, т. 12: 188-204.
-
18. F u r u c a w a Y., B u c o v a c M.J. Embryo sac development in sour cherry during the pollination period as related to fruit set. Hort. Sci., 1989, 24(6): 1005-1008.
-
19. С е д о в Е.Н., С е д ы ш е в а Г.А., С е р о в а З.Н., Г о р б а ч е в а Н.Г. Доноры для создания триплоидных и иммунных к парше сортов яблони. Вест. Саратовского гос-агроуниверситета им. И.В. Вавилова, 2010, 7: 35-38.
-
20. М а ш к и н С.И. Цитогенетический анализ спонтанных и искусственных гибридов и аллополиплоидов Cerasus . II. Генетика, 1994, 30(прил.): 98.
-
21. Р о м а н о в а Н.П., Ш е л а б о т и н Г.П., Л е о н ч е н к о В.Г., X а н и н а Н.П., Н и к о л ь с к и й Б.В., Р а с т о р г у е в С.Л., Т у р о в с к и й И.И. Методические рекомендации по применению цитологических методов в плодоводстве. М., 1988.
-
22. Т о п и л ь с к а я Л.А., Л у ч н и к о в а С.В., Ч у в а ш и н а Н.П. Изучение соматических и мейотических хромосом смородины на ацетогематоксилиновых давленых препаратах. Бюл. научно-технической информации ЦГЛ им. И.В. Мичурина (Мичуринск), 1975, 22: 58-61.
-
23. Я н д о в к а Л.Ф., Ш а м р о в И.И. Фертильность пыльцы Cerasus vulgaris и Cerasus tomentosa ( Rosaceae ). Ботанический журнал, 2006, 91(2): 206-218.
-
24. T a b a c a r u C.D., S t e f u r e a c T.J. L'embryogenese chez certaines sortes de griottiers ( Prunus cer. L.). Rev. Roum. Biol. Ser. Biol. Veget., 1986, 31(1): 19-22.
-
1 ФГБОУ ВПО Тамбовский государственный университет Поступила в редакцию
им. Г.Р. Державина, Медицинский институт, 27 января 2009 года
392000 г. Тамбов, ул. Интернациональная, 33, e-mail: yandovkaTGU@mail.ru ;
-
2ФГБОУ ВПО Мичуринский государственный
аграрный университет,
393770 Тамбовская обл., г. Мичуринск, ул. Интернациональная, 101,
MICROSPOROGENESIS IN THE Pyrus , Malus , Cerasus , Microcerasus VARIETIES AND THEIR HYBRIDS
L.F. Yandovka1, R.V. Papikhin2
S u m m a r y
The features of microsporogenesis were studied in Pyrus communis , Malus domestica , Cerasus vulgaris , Microcerasus tomentosa , Cerasus avium species and their interspecific and intergeneric hybrids. The typical and non-typical abnormalities in microsporogenesis were described. The typical — were found in all studied plants, the non-typical — in hybrids, as rule. The non-typical abnormalities caused the low fertility of pollen grains.
Научные собрания

МЕЖДУНАРОДНЫЙ КОНГРЕСС ПО КАРТОФЕЛЕВОДСТВУ (WPC)
(27-30 мая 2012 года, г. Эдинбург, Шотландия)
Лозунг Конгресса — «Думать глобально.
Побеждать локально». Событие сфокуси ровано на ключевых фундаментальных проблемах производства, переработки и потребления картофеля и их практическом решении.
Информация:
