Минералого-геохимические особенности пещерной фоссилизации ископаемых костей на примере пещеры Иманай (Южный Урал)
")
Автор: Силаев В.И., Паршукова М.Н., Гимранов Д.О., Филиппов В.Н., Киселва Д.В., Смолева И.В., Тропников Е.М., Хазов А.Ф.
Журнал: Вестник Пермского университета. Геология @geology-vestnik-psu
Рубрика: Минералогия, кристаллография
Статья в выпуске: 4 т.19, 2020 года.
Бесплатный доступ
Впервые получены результаты комплексных минералого-геохимических исследований ископаемых костей плейстоценовых млекопитающих из пещеры Иманай (Южный Урал) с использованием оптической и сканирующей электронной микроскопии, статистического объемометрического метода определения нанопористости, термического, рентгеноструктурного и рентгено-флюоресцентного анализов, газовой хроматографии, ИК, КР и РЛ спектроскопии, ИСП МС, изотопной масс-спектрометрии. Цель исследований - получение экспериментальных данных о характере и степени фоссилизации костных остатков в условиях пещер, оценка возможностей палеоэкологических реконструкций по пещерным ископаемым костям. Обсуждаются данные о микростроении костных останков, нанопористости, химическом составе и концентрации микроэлементов, рентгеноструктурных и термических свойствах, изотопном составе минеральной и органической компонент в костях. Согласно полученным данным, фоссилизация костных остатков в условиях пещер принципиально отличается на всех уровнях строения и состава ископаемых костей от таковой во внепещерных обстановках. В частности, пещерные кости отличаются интенсивной эпигенетической кальцитизацией, в 3-5 раз большим содержанием СО2 и аномально высокими значениями атомного отношения Ca/P, но многократно уступают по содержанию иллювиированных минеральных загрязнений, обогащению микроэлементами вмещающей геологической среды, содержанию Сорг и коллагена. По изотопному составу углерода и кислорода в биоапатите пещерный костный детрит является аномально тяжелым, сближаясь с изотопным составом углерода и кислорода в морских известняках. Приведены некоторые палеоэкологические реконструкции, включая диету пещерных животных, рассмотрены доказательства обитания в пещере Иманай поздних неандертальцев.
Неоплейстоцен, южный урал, пещера иманай, пещерные львы и медведи, минералого-геохимические исследования, особенности пещерной фоссилизации ископаемых костей, палеоэкологические реконструкции, поздние неандертальцы
Короткий адрес: https://sciup.org/147246195
IDR: 147246195 | УДК: 56+(470.55/.58) | DOI: 10.17072/psu.geol.19.4.323
Mineralogical and geochemical features of cave fossilization of bone detritus on the example of Imanay cave (South Urals)
First time, the results of complex mineralogical and geochemical studies of fossil bone detritus of Pleistocene mammals from the Imanay Cave (South Urals) were obtained using optical and scanning electron microscopy, a statistical volumetric method for nanoporosity determination, thermal, X-ray structural and X-ray fluorescence analyzes, gas chromatography, IR, Raman and x-ray luminescent spectroscopy, ICP MS, and isotope mass spectrometry. The aim of the research was to obtain experimental data on the nature and degree of fossilization of bone remains in caves, and to assess the possibilities of paleoecological reconstructions from cave fossil bones. The data on the microstructure of bone remains, nanoporosity, chemical composition and concentration of trace elements, X-ray structural and thermal properties, isotopic composition of the mineral, and organic components in bones are discussed. According to the data obtained, fossilization of bone remains in caves is fundamentally different at all levels of structure and composition of fossil bones from those outside caves environments. In particular, cave bones are distinguished by intense epigenetic calcitization, 3-5 times higher CO2 content and abnormally high Ca/P atomic ratios, but they are many times inferior in terms of the content of illuminated mineral impurities, enrichment in microelements of the host geological environment, content of Corg and collagen. According to the isotopic composition of carbon and oxygen in bioapatite, cave bone detritus is abnormally heavy, approaching the isotopic composition of carbon and oxygen in marine limestones. Some paleoecological reconstructions are presented and evidence of the habitation of the late Neanderthals in the Imanai cave is considered.
Текст научной статьи Минералого-геохимические особенности пещерной фоссилизации ископаемых костей на примере пещеры Иманай (Южный Урал)
В современной отечественной палеонтологии при всей ее тотальной аналитической запущенности (Силаев и др., 2016) есть все- таки область, выделяющаяся даже на таком фоне. И эта область – пещерные ориктоценозы, уровень исследований которых практически не изменился с 18–19 вв., т. е. со вре-
мени первых собирателей ископаемых костей в пещерах (Чикишев, 1973; Горбунова, 1988; Дублинский, Козлова, 1994; Оводов, 2009), на территории, например, Западной Сибири (Ф.И. Страленберг1, И.Г. Гмелин, И. Пестов, И.С. Боголюбский), Алтае-Саянской области (А.Г. Влангели2, И.Ф. Брандт, П.С. Паллас, Ф.Б. Геблер, А.И. Кулибин3), Хакасии (П. С. Паллас), Грузии (Ф. Дюбуа де Монпере4), Крыма (К. С. Мережковский5). Лишь открытие феноменальной костеносно-сти элювиальных грунтов в пещере Иманай на Южном Урале (Яковлев и др., 2016; Гим-ранов и др., 2016; Паршукова, 2016; Гимра-нов и др., 2017; Гимранов и др., 2018; Gim-ranov et al., 2018) потребовало кардинального пересмотра палеонтолого-археологической традиции поверхностного описания и зоологической систематизации пещерного костного детрита в пользу экспериментального естествознания. Тем более что для этого уже существует вполне адекватная научная база (Смирнов и др., 2009; Вотяков и др., 2009; Силаев и др., 2015).
Следует подчеркнуть, что в настоящее время реализация программы экспериментальных научных исследований костных остатков пещерных животных весьма актуализирована открытием в 2018 г. в Крыму уникальной пещеры Таврида с громадным комплексом костного детрита, датирующегося в диапазоне от миоцена до неоплейстоцена. Очевидно, что без надежных знаний об особенностях пещерной фоссилизации ископаемых костей полноценное изучение ориктоценоза такой пещеры просто невозможно.
Общие сведения
Карстовая пещера Иманай была обнаружена в 2010 г. госинспектором Национального парка Башкирия Р. Р. Альмухаметовым в борту одноименного лога в 7 км к северо-востоку от пос. Нугуш (рис. 1, а). Пещера длиной до 100 м с гротом размером 5 × 6 м располагается в сероцветных доломитизиро-ванных известняках кунгурского яруса нижней перми. У входа в пещеру и внутри нее обнаружен редкий по сложности ориктоценоз плейстоценовых млекопитающих (табл. 1), центральное место в котором занимают костные остатки пещерных львов и медведей. Датирование этих костей было осуществлено в ЦКП «Лаборатория радиоуглеродного датирования и электронной микроскопии» Института географии РАН.
Таблица 1. Иманай-пещерный ориктоценоз плейстоценового возраста (Gimranov, Kosintsev, 2020)
|
Виды ископаемой фауны |
Число идентифицирован-ных образцов |
|
Canis sp. |
5 |
|
Marmota sp. |
13 |
|
Castor fiber (Linnaeus, 1758) |
2 |
|
Pantera ex. gr. Fossilis-spelaea |
536 |
|
Cuon sp. |
2 |
|
Canis lupus (Linnaeus, 1758) |
42 |
|
Canis sp. |
1 |
|
Vulpes lagopus (Linnaeus, 1758) |
2 |
|
V. vulpes (Linnaeus, 1758) |
3 |
|
Vulpes sp. |
17 |
|
Meles sp. |
2 |
|
Gulo gulo (Linnaeus, 1758) |
24 |
|
Mustela ermine (Linnaeus, 1758) |
1 |
|
M. ex.gr. putorius-eversmanii |
6 |
|
Mustelidae gen. indet. |
1 |
|
Ursus «savini» (Andrews, 1922) |
9414 |
|
Ursus arctos (Linnaeus, 1758) |
16 |
|
Ursus thibetanus (Cuvier, 1823) |
3 |
|
Mammunthus primigenius (Blumenbach, 1799) |
2 |
|
Eguus ferus (Boddaert, 1785) |
4 |
|
Rhinocerotidae gen. indet |
2 |
|
Saiga tatarica (Linnaeus, 1766) |
22 |
|
Ovis ammon (Linnaeus, 1758) |
17 |
|
Итого |
10137 |
Калиброванные значения возраста составили для львов – 34940 ± 140, а для медведей – 42410 ± 260 лет назад. Полученные датировки приходятся примерно на средину кар-гинского (средневалдайского) интерстадиала, а точнее, на разные периоды относительного похолодания в рамках этого времени — более ранний (44–41 тыс. лет назад) для пе- щерных львов и более поздний (35–30 тыс. лет назад) для медведей.
Кроме костного детрита в пещерных отложениях археологами найдено множество каменных орудий мустьерской эпохи – среднего палеолита (Гимранов и др., 2016), приписываемых поздним неандертальцам, а также обнаружены очажные пятна, свидетельствующие о непосредственном обитании древних людей в пещере.
Изученный профиль костеносных элювиальных отложений в пещере Иманай имеет мощность около 1 м. Он почти нацело сложен гранулометрически весьма неоднородными грунтами серого цвета с буроватым или розоватым оттенками (рис. 1, б). В направлении сверху вниз профиль был подразделен на 12 горизонтов или (условно) слоев (рис. 1, в), гранулометрический состав которых определяется пятью фракциями: гравийной, песчаной грубозернистой, песчаной крупно-среднезернистой, песчаной мелко-тонкозернистой и алевропелитовой (глинистой). Обобщая полученные нами данные (Силаев и др., 2018), можно прийти к следующей литологической номенклатуре пещерного костеносного элювия.
Слой 1 : гравий песчаный, отвечающий по литологическому составу доломитисто-известковому мергелю.
Слой 2 : песок гравийный доломитисто-известковый.
Слой 3 : песок алевро-гравийный доломитисто-известковый.
Слой 4 : песок алевро-гравийный известководоломитовый.
Слой 5 : песок алевро-гравийный, отвечающий по составу известковисто-доломито-вому глинистому мергелю.
Слой 6 : песок алевро-гравийный доломитовоизвестковый.
Слой 7 : песок алевро-гравийный некарбонатный. Слой 8 : песок алевро-гравийный известковый. Слой 9 : песок алевро-гравийный известководоломитовый.
Слой 10 : песок алевро-гравийный доломитисто-известковый
Слой 11 : песок гравийный доломитисто-известковый.
Слой12 : песок гравийный, отвечающий по составу доломитисто-известковому мергелю.
На треугольнике гранулометрического состава костеносных грунтов (рис. 2, а) пробы почти всех слоев элювиальных отложений в пещере Иманай попали в поле гравийных песков, за исключением образцов песчаного гравия в слое 1 и глинисто-гравийных песков в слое 9. Очевидно также, что основную массу в исследуемых грунтах составляют пески, широко варьирующиеся по гранулометрии от грубо- до мелко-тонкозернистых. Судя по композиции песчаных фракций, иманайские грунты являются микститовыми (рис. 2, б), т. е. совершенно несортированными, что указывает на их автохтонное (непереотложен-ное) происхождение.
Таким образом, костеносные грунты в пещере Иманай по гранулометрическому составу являются песчано-гравийными, гравийно-песчаными и глинисто-гравийно-песчаными. При этом в направлении сверху вниз по профилю отложений в интервале до слоя 10 включительно выявляется тенденция к переходу от более крупнообломочных (песчано-гравийных и гравийно-песчаных) осадков к несколько менее крупнообломочным (глинисто-гравийно-песчаным). В самых нижних слоях 11 и 12 пещерные отложения опять становятся гравийнопесчаными. Отсутствие признаков флюи-дальности, гранулометрической сортировки и механической обработки обломков свидетельствует об их именно элювиальном происхождении. При этом признаков существенных гидролитических изменений (химического выветривания) грунтов тоже не выявлено, что можно рассматривать как благоприятное обстоятельство для перспективных палеонтологических и палеозоологических поисков.
По химическому и минеральному составу элювиальные отложения в пещере являются фосфатно-карбонатно-силикатными. В силикатную компоненту входят кварц, альбит, диоктаэдрические гидрослюды и хлориты. Карбонатная компонента варьирует от известковой до известковисто-доломитовой, что комплементарно составу подвергнувшихся карстованию нижнепермских карбонатных пород. Присутствие фосфатного вещества в грунтах обусловлено, очевидно, дезинтеграцией костных остатков, их частичным растворением и переотложением растворенного фосфатного вещества.

Рис. 1. Географическое положение пещеры Иманай (а), общий вид (б) и строение (в) выявленного в ней профиля костеносных элювиальных грунтов: 1 – стенки карбонатной пещеры; 2–4 – грунты соответственно сероцветные, светло-бурые и серовато-коричневые; 5 – натеки карбонатного туфа; 6 – каменные артефакты, отнесенные к мустьерской эпохе (среднему палеолиту); Л и М – наличие костного детрита соответственно пещерных львов и медведей в слоях элювиальных грунтов
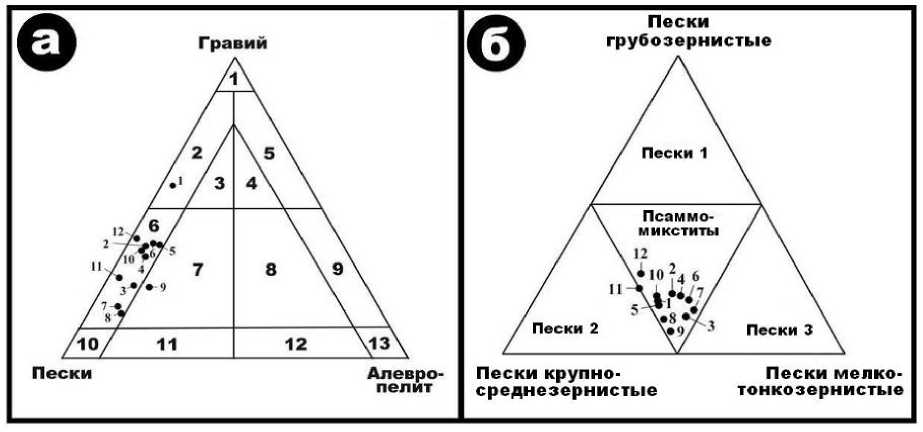
Рис. 2. Общая гранулометрическая характеристика костеносных элювиальных грунтов в пещере Иманай (а) и оценка гранулометрического состава их песчаной компоненты (б). а : 1 – гравий; 2–5 – гравий соответственно песчаный, глинисто-песчаный, песчано-глинистый и глинистый; 6, 7 – пески соответственно гравийные и глинисто-гравийные; 8, 9 – глины соответственно гравийно-песчаные и гравийные;10 – пески; 11 – пески глинистые; 12 – глины песчаные; 13 – глины. б : пески преимущественно грубозернистые (1), крупно-среднезернистые (2), мелко-тонкозернистые (3); псаммомикс-титы – несортированная гранулометрическая смесь песков. Точками показаны составы исследованных образцов пещерных грунтов из слоев 1–12
Присутствие в грунтах тонкодисперсного костного материала подтверждается не только повышенным (до 0.33 мас. %) в них содержанием Сорг, но и результатами термического анализа. Таким образом, почти весь интервал костеносных элювиальных грунтов в пещере Иманай (выше 12 горизонта) образовался уже после максимума последнего оледенения (LGM), т. е. в период после 15 тыс. лет назад (Кольфсхотен и др., 2008).
Объекты и методы исследований
Важнейшее место в иманайском пещерном ориктоценозе занимают костные остатки пещерных медведя и льва. В обоих случаях мы имеем дело с особенными феноменами – малым пещерным медведем, обитавшим большей частью, как считается, вне пещер, и так называемым гигантским пещерным львом (Сотникова, Гимранов, 2017). Последний факт доказывается анализом размеров зубов. Зубы иманайских львов – и самцов, и самок (рис. 3, а) – значительно превышают таковые у обычных пещерных львов (Ba-rucka, 2008) и тем более у современных львов (рис. 3, б). При этом обнаруживается, что у современных и ископаемых пещерных львов половой диморфизм в интегральных характеристиках зубов выражается совершенно по-разному. Если у современных львов самцы характеризуются более крупными (примерно на 20–25 %) и при этом более удлиненными (острыми) зубами, то у пещерных львов самцы отличались от самок, хотя и более крупными, но более «тупыми» зубами (рис. 3, в).
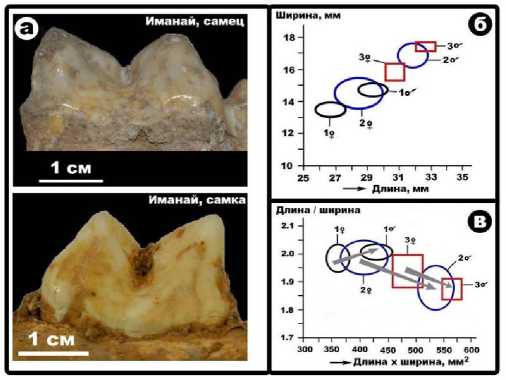
Рис. 3 . Форма и размеры нижних хищных (car-nassial) зубов иманайских пещерных львов (а) и графики полового диморфизма по зубам (б, в) для современных львов (1), для львов из пещеры Вие-шховска Гурна (2) и пещеры Иманай (3)
Таким образом, получается, что иманай-ские львы, судя по зубам, действительно отличались более крупными размерами, чем обычные Panthera (Leo) spelaea. Как известно, в период перехода от среднего к поздне- му плейстоцену в Европе появился Panthera (Leo) spelaea intermedius, промежуточный по размерам между архаичным Pantera (Leo) antrox и относительно мелким Panthera (Leo) spelaea (Argani et al., 2007; Сотникова, Кириллова, 2017). Не исключено, что именно эта промежуточная форма ископаемого пещерного льва, обозначенная в табл. 1 как Pantera ex. gr. Fossilis-spelaea, и обнаружена в пещере Иманай (рис. 4).
Непосредственными объектами наших исследований послужили две коллекции остеологически (анатомически) разнообразных костных остатков пещерных львов и медведей (табл. 2). Этот костный детрит не несет признаков механической обработки и варьирует по размерам в львиной и медвежьей группах образцов в сравнительно узких пре- делах.

Рис. 4 . Размеры пещерных и современных львов (а) на фоне иллюстрации Велизара Семионовски6 «Недолгий триумф» вымышленного факта плейстоценовой истории пещеры Иманай (б)
Наибольшим остеологическим разнообразием характеризуются костные остатки пещерного медведя, будучи представленные фрагментами черепа, нижней челюсти, ат-
6 Очень талантливый болгаро-американский зоолог, специализирующийся на научной и художественной иллюстрации вымерших млекопитающих и палеоландшафтов.
ланта, бедренной кости, костей метаподия, фаланг конечностей, зубов и клыков (рис. 5), размеры которых составляют соответственно (см): (11.4 ± 3.7) × (5.3 ± 2.6); 17.7 × 6; (4 ± 1.9) × (2.3 ± 1.7); (10.2 ± 7.7); (3.35 ± 0.7) × (1.5 ± 0.2); (1.7 ± 0.8) × (1.2 ± 0.5); (4.8 ± 3) × (1.8 ± 0.9). Общая статистика размеров исследованных медвежьих костей определяется как (5.1 ± 4.2) × (2.5 ± 2) см.
Таблица 2. Исследованный костный детрит львов (1–15) и медведей (16–30)
|
№п/п |
№ обр. |
Остеология |
Горизонт |
|
1 |
622 |
Позвонок |
2 |
|
2 |
823 |
« |
4 |
|
3 |
2847 |
« |
5 |
|
4 |
492 |
« |
6 |
|
5 |
2869 |
« |
« |
|
6 |
2803 |
Ребро |
1 |
|
7 |
2411 |
« |
2 |
|
8 |
2870 |
« |
6 |
|
9 |
2871 |
« |
« |
|
10 |
3111 |
Сесамовидная кость |
1 |
|
11 |
3112 |
« |
« |
|
12 |
2987 |
« |
2 |
|
13 |
3046 |
« |
4 |
|
14 |
2974 |
Фаланга № 2 |
3 |
|
15 |
2975 |
Клык |
« |
|
16 |
1/1 |
Фаланга |
1 |
|
17 |
1/2 |
Череп |
« |
|
18 |
2 |
Метаподий |
2 |
|
19 |
3/1 |
Атлант |
3 |
|
20 |
3/2 |
Ребро |
4 |
|
21 |
4/1а |
Зубная эмаль |
« |
|
22 |
4/1б |
Дентин |
« |
|
23 |
4/2 |
Моляр нижний |
« |
|
24 |
7 |
Клык в грунте |
7 |
|
25 |
8 |
Бедро |
8 |
|
26 |
9 |
Фаланга |
9 |
|
27 |
(10+11)/1 |
Челюсть |
10 + 11 |
|
28 |
(10+11)/2 |
Премоляр верхний |
« |
|
29 |
12м |
Метаподий |
12 |
|
30 |
12ф |
Фаланга |
« |
К костным остаткам иманайского пещерного льва относятся позвонки, ребра, сесамовидная кость и клыки (рис. 6), размеры которых заметно превышают размеры костей пещерного медведя (см): (10.3 ± 6.2) × (7.2 ± 3.3); (9.2 ± 3.7) × (2.1 ± 0.7); (30 ± 13.4) × (13.5 ± 4.6); 13.7 × 10. Общая статистическая оценка размеров костного детрита пещерного льва – (14.9 ± 10.4) ± (6.8 ± 5.5) см. В сечениях костей наблюдается достаточно хорошо сохранившееся внутреннее строение с контрастно различающимися компактным и губчатоячеистым типами костной ткани (рис. 7).

Рис. 5 . Исследованные костные остатки има-найского пещерного медведя

Рис. 6. Исследованные костные остатки има-найского пещерного льва

Рис. 7 . Типичные сечения костей иманайского пещерного льва: 1, 2 – соответственно компактный и губчато-ячеистый типы костной ткани
Таким образом, проведенные измерения и расчеты приводят к выводу о том, что костные остатки иманайских львов и медведей по размерам статистически сильно различаются: обломки скелетов львов в среднем почти в три раза крупнее, чем обломки скелетов медведей. Интересно, что примерно такое же различие обнаруживается и при отдельном сопоставлении клыков. Из всего этого следует, что причиной выявленных различий являются не только особенности фрагментиза-ции скелетов, но, вероятно, и изначальные расхождения в массивности и, возможно, прочности костей.
Раскопки и палеозоологические исследования костного детрита финансировались за счет гранта Российского научного фонда (проект № 20-74-00041). В ходе исследований иманайских пещерных костей применялся широкий комплекс современных аналитических методов: гранулометрический анализ грунтов ситовым методом (в диапазоне от +2 до –0.06 мм); термический анализ (DTG-60А/60 АН, Shimazu); аналитическая химия (О. В. Кокшарова); определение содержания Сорг методом кулонометрического титрования (С. А. Забоева); рентгеновская дифрактометрия (Ю. С. Симакова, XRD-6000); рентгенофлюоресцентный анализ (С. Т. Неверов, XRD-1800, Shimazu); рамановская спектроскопия (Е.А. Васильев, Lab Ram800 Horiba, He-Ne лазер, 632.8 нм); рентгенолюминесцентная спектроскопия (Ю. В. Глухов, установка, состоящая из монохроматора AAS-1, рентгеновского аппарата УРС-1.0, блока электронной регистрации ФЭУ106 и ртутной лампы ДРК-120 (λвозб = 365 нм); оптическая микроскопия (компьютеризированный комплекс OLYMPUS BX51); аналитическая растровая электронная микроскопия (JSM-6400 Jeol); определение нанопористости по кинетике адсорб-ции/десорбции азота (Nova 1200e, Quantachrome Instruments); определение содержания микроэлементов методом ИСП-МС (ELAN 9000 Perkin Elmer, Германия); элементный анализ костного коллагена методом газовой хроматографии (Е. А. Туманова; ЕА 1110 (СНNS-O), CE Instruments, Италия); инфракрасная спектроскопия (М. Ф. Самотолкова; ФТ-2 Инфралюм); анализ изотопного состава С, О в биоапатите и С, N в костном коллагене (проточный масс-спектрометр Delta V. Avantage и аналитический комплекс Thermo Fisher, Германия).
ИСП-МС анализы выполнены в ЦКП УрО РАН «Геоаналитик» при поддержке грантом РНФ № 16-17-10283, КР спектры получены в Санкт-Петербургском горном университете. Все остальные аналитические работы осуществлены в Институте геологии Коми НЦ УрО РАН.
Микростроение и ультрапористость органоминерального костного композита
Внутреннее строение костных остатков иманайских львов и медведей исследовалось методом оптической поляризационной микроскопии. Полученные результаты свидетельствуют в целом о значительной степени деградации биоапатита костей, но различной для пещерных медведей и львов. Исследованы фрагменты атланта, костей метаподия, черепа и фаланги пещерных медведей (рис. 8).
Атлант (шейный позвонок) — проанализирована плотная костная ткань, характеризующаяся минимальной степенью коллофа-низации (аморфизации) биоапатита и умеренной сохранностью его волокнистости и остеон-структуры.
Размеры сечений остеонов варьируют в пределах (269 ± 69) × (204 ± 41) мкм, коэффициент их изометричности оценивается в (0.78 ± 0.15). Остеоциты почти не наблюдаются.
Метаподий (кости конечностей) – плотная костная ткань с умеренной степенью коллофанизации биоапатита и сохранности волокнистости и остеон-структуры. Остео-циты не наблюдаются. Размеры сечений остеонов колеблются в пределах (476 ± 152) × (224 ± 81) мкм, коэффициент изометрично-сти составляет (0.49 ± 0.18).
Череп – плотная ткань со значительной степенью коллофанизации биоапатита и лишь фрагментарно сохранившейся остеон-структурой. Размер сечений остеонов изменяется в пределах (448 ± 103) × (261 ± 106) мкм, коэффициент изометричности сечений – (0.58 ± 0.14). Многие каналы заполнены микрокристаллическим карбонатом.
Фаланги (пальцы конечностей) – наблюдается плотный и губчатый типы костной ткани с сильно коллофанизированным биоапатитом. В плотной ткани остеон-структура сильно затушевана и практически не наблюдается. В губчатой ткани трабекулы нарушены, размер ячей варьируется в пределах (722 ± 218) × (389 ± 77) мкм, коэффициент изометричности оценивается в (0.59 ± 0.25).
Оптикомикроскопическим методом были исследованы фрагменты ребер, позвонки, сесамовидные кости, фаланги и клыки пещерных львов (рис. 9).
Ребра – исследована плотная ткань, сложенная практически нацело коллофанизиро-ванным и потому оптически непрозрачным биоапатитом. Наблюдается теневая волокнистая текстура сильно измененной кости.
Позвонки — плотная и губчатая ткани. За редким исключением ткани сложены тотально коллофанизированным оптически непрозрачным биоапатитом. Тем не менее в плотной ткани улавливается теневая остеон-структура. Видимые размеры сечений остеонов колеблются в пределах (389 ± 154) × (243 ± 102), коэффициент изометричности составляет (0.66 ± 0.17). В редких случаях в костной ткани позвонков наблюдаются участки, не затронутые коллофанизацией с хорошо сохранившейся первичной структурой. Губчатая ткань – плохо сохранившаяся, с сильно деградированными трабекулами, в ячеях наблюдается переотложенный карбонат.
Сесамовидная кость – плотная и губчатая костные ткани. Большинство образцов ткани сложены тотально коллофанизированным оптически непрозрачным биоапатитом. Остеон-структура едва различается, видимые размеры сечений остеонов оцениваются в (291 ± 50) × (226 ± 63) мкм, коэффициент изометричности – (0.78 ± 0.19). Кроме того, в плотной ткани наблюдаются каверны выщелачивания и микротрещины, инкрустированные микрокристаллическим карбонатом.
Размер каверн составляет (291 ± 176) × (143 ± 90) мкм, коэффициент изометрично-сти — (0.54 ± 0.23). Ячеи в губчатой ткани также минерализованы переотложенным карбонатом. В единичных случаях обнаружены фрагменты сесамовидной кости с хо- рошей сохранностью губчатой ткани, сложенной биоапатитом с незначительной степенью коллофанизации. В трабекулах наблюдается волокнистое строение, их толщина варьируется в пределах (276 ± 118) мкм. Ячеи – неминерализованные с размером (468 ± 157) × (294 ± 80) мкм, коэффициент изометричности составляет (0.66 ± 0.2).
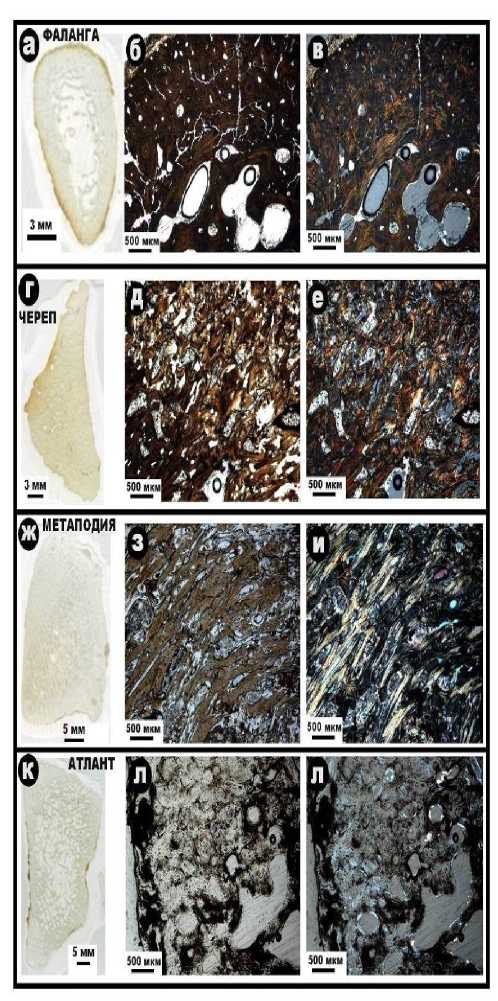
Рис. 8 . Микростроение костного детрита иманайских пещерных медведей под оптическим микроскопом: а, г, ж, к – сканы шлифов; б, д, з, л – изображения в режиме параллельных николей; в, е, и, м – изображения в режиме скрещенных николей
Фаланги – плотная и губчатая костные ткани с оптически непрозрачным нацело коллофанизированным биоапатитом. Остеон-структура проступает очень нечетко, видимый размер сечений остеонов колеблется в пределах (341 ± 172) × (235 ± 121) мкм, коэффициент изометричности – (0.71 ± 0.15). Трабекулы в губчатой ткани сильно деградированы, размер ячей составляет (657 ± 313) × (333 ± 146) мкм, коэффициент изометрично-сти – (0.55 ± 0.2).
Клык – сложен нацело коллофанизиро-ванным биоапатитом, разбитым сетью тонких трещин. Разделение на дентин и эмаль не наблюдается.
Обобщение результатов оптикомикроскопических исследований приводит к следующему выводу. Иманайский костный детрит в целом обнаруживает степень вторичных изменений, значительно превышающую степень изменения близких по возрасту ископаемых костей животных, захороненных во внепещерных условиях, например, плейстоценовых лошадей на открытой П. Ю. Павловым западно-уральской стоянке Заозерье (Силаев и др., 2019).
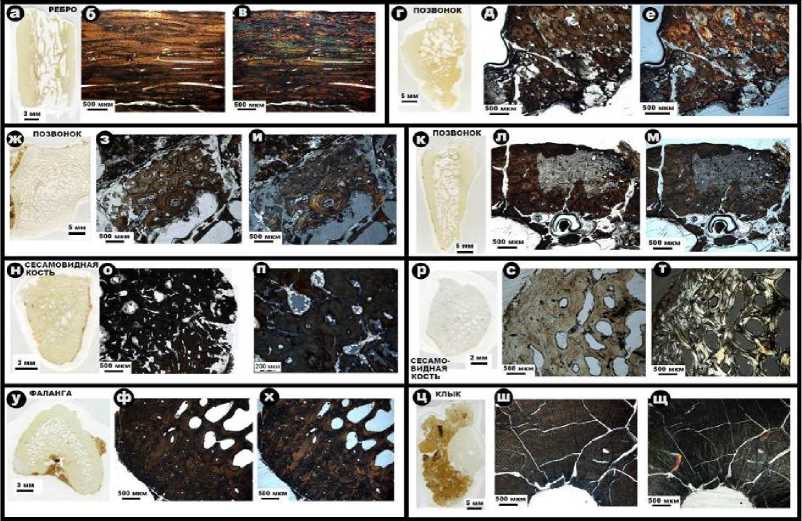
Рис. 9 . Микростроение костного детрита иманайских пещерных львов под оптическим микроскопом: а, г, ж, к, н, р, у, ц – сканы шлифов; б, д, з, л, о, с, ф, ш – изображения в режиме параллельных николей; в, е, и, м, п, т, х, щ – изображения в режиме скрещенных николей
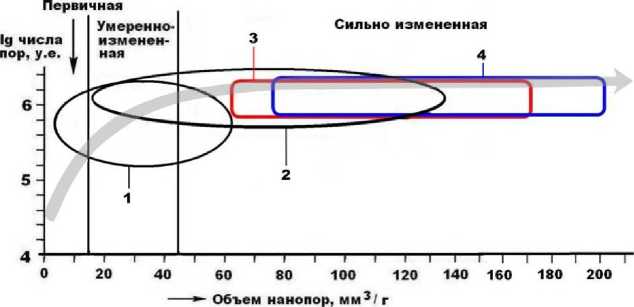
Рис. 10 . Нанопористость ископаемых костей плейстоценовых мегамлекопитающих: 1, 2 – мамонтовая фауна соответственно с Печорского Приуралья (Silaev et al., 2017) и Омского Прииртышья (коллекция А. А. Бондарева); 3, 4 – иманайские соответственно пещерные львы и медведи
Таблица 3. Характеристика нанопористости в костных остатках иманайских пещерных львов (1– 12) и медведей (13–26)
|
№ п/п |
№ обр. |
Объект |
S o , м2/г |
V o , мм3/г |
R o , нм |
V o i, нм3 |
N o |
lgN o |
|
1 |
492 |
Позвонок |
88.97 |
168 |
2.402 |
58.07 |
2893.06 |
6.46 |
|
2 |
622 |
« |
75.61 |
181.7 |
4.81 |
465.41 |
390.41 |
5.59 |
|
3 |
823 |
« |
79.63 |
94.4 |
2.372 |
55.92 |
1688.13 |
6.23 |
|
4 |
2847 |
« |
101.3 |
165.6 |
3.271 |
146.64 |
1129.3 |
6.05 |
|
5 |
2411 |
Ребро |
77.87 |
94.6 |
2.429 |
60.05 |
1575.35 |
6.23 |
|
6 |
2803 |
« |
62.57 |
83.2 |
2.661 |
78.95 |
1053.83 |
6.02 |
|
7 |
2871 |
« |
45.92 |
51.6 |
2.246 |
47.47 |
1087.0 |
6.04 |
|
8 |
2987 |
Сесамовидная кость |
75.98 |
157.5 |
4.146 |
298.61 |
527.75 |
5.72 |
|
9 |
3046 |
« |
55.31 |
66.1 |
2.39 |
57.2 |
1155.59 |
6.06 |
|
10 |
3111 |
« |
46.19 |
53.3 |
2.307 |
51.45 |
1035.96 |
6.01 |
|
11 |
2974 |
Фаланга |
82.18 |
102.7 |
2.499 |
65.39 |
1570.58 |
6.2 |
|
12 |
2975 |
Клык |
122.7 |
220.7 |
3.597 |
195 |
1131.79 |
6.05 |
|
Среднее |
75.5 |
116.77 |
2.88 |
125.66 |
1285.27 |
6.06 |
||
|
СКО |
21.45 |
55.01 |
0.82 |
126.38 |
615.17 |
0.22 |
||
|
Коэффициент вариации, % |
28.4 |
41.1 |
28.5 |
3.6 |
47.9 |
3.6 |
||
|
13 |
1/2 |
Череп |
82.02 |
152.4 |
3.717 |
215.17 |
708.28 |
5.85 |
|
14 |
(10+11)/1 |
Челюсть |
99.35 |
120.7 |
2.43 |
60.12 |
2024.28 |
6.3 |
|
15 |
3/1 |
Атлант |
121 |
269.2 |
4.45 |
369.23 |
729.08 |
5.86 |
|
16 |
3/2 |
Ребро |
67.8 |
84.3 |
2.448 |
64.32 |
1310.63 |
6.12 |
|
17 |
8 |
Бедро |
61.34 |
63.6 |
2.074 |
37.38 |
1701.44 |
6.23 |
|
18 |
2 |
Кость метаподия |
89.95 |
165.9 |
3.689 |
210.35 |
788.68 |
5.9 |
|
19 |
12м |
« |
98.7 |
129.3 |
2.62 |
75.36 |
1715.76 |
6.23 |
|
20 |
1/1 |
Фаланга |
82.02 |
152.4 |
3.717 |
215.17 |
708.28 |
5.85 |
|
21 |
9 |
« |
111.3 |
201.1 |
3.631 |
200.6 |
1002.49 |
6.0 |
|
22 |
12ф |
« |
102.3 |
115.7 |
2.262 |
48.49 |
2447.93 |
6.38 |
|
23 |
4/1б |
Премоляр |
51.97 |
83.5 |
3.215 |
139.24 |
599.68 |
5.78 |
|
24 |
4/2 |
Моляр |
34.39 |
39 |
2.268 |
48.88 |
797.87 |
5.9 |
|
25 |
7 |
Клык нижний |
111.3 |
171.2 |
3.075 |
121.83 |
1405.24 |
6.15 |
|
26 |
(10+11)/2 |
Премоляр нижний |
130.4 |
305.4 |
3.15 |
131.34 |
2325.26 |
6.19 |
|
Среднее |
88.36 |
140.01 |
3.09 |
145.03 |
1304.64 |
6.04 |
||
|
СКО |
27.61 |
62.25 |
0.76 |
103.34 |
641.94 |
0.21 |
||
|
Коэффициент вариации, % |
31.2 |
44.5 |
24.6 |
71.2 |
49.2 |
3.5 |
||
Примечание. S o , м2/г – удельная поверхность нанометровых пор; V o , мм3/г – совокупный объем пор; R o , нм – средний размер пор; V o i, нм3 – объем единичной поры; N o – условное количество пор на удельный объем.
Степень фоссилизации костей в изученной коллекции различна. Кости пещерных львов демонстрируют большую изменен-ность, чем кости медведей, что проявляется в степени коллофанизации костного биоапатита.
Важнейшей структурной характеристикой костного композита является ультрапористость в нанометровом диапазоне, которая сильно деградирует в ходе фоссилизации, характеризуя степень вторичного изменения и относительный возраст ископаемых костей. Пещерные кости отличаются более раз- рушенной наноструктурой (табл. 3), чем того же возраста кости млекопитающих, захороненных во внепещерных обстановках (рис. 10). Костный детрит пещерных медведей статистически демонстрирует большую степень деградации наноструктуры, чем кости львов. Выявляется однотипная для львов и медведей последовательность увеличения степени изменения анатомических разностей костей в направлении от зубов к ребрам, бедренным и сесамовидным костям и далее к черепным и челюстным фрагментам, костям метаподия, атланту и фалангам.
Химический и нормативно-минеральный составы
На валовый химический состав анализировались смывы грунтовых загрязнений с костей и сами кости пещерных животных. Полученные результаты пересчитывались на нормативно-минеральный состав.
По химическому составу смывы с костей (табл. 4, 5) оказались довольно близкими к составу пещерных грунтов, что свидетельствует об автохтонности исследуемого костного детрита по отношению к костеносному пещерному элювию. В нормативно-минеральный состав смывов входят кварц, полевые шпаты, филлосиликаты (слюды и хлориты), а в львиных костях еще и пирит. Тем не менее в составе смывов от 50 до 65 % приходится на апатит и карбонаты. Появление этих минералов в смывах отражает, очевидно, результат гидролитического разложения костного биоапатита в условиях карстовых пещер. Принципиальное уравнение соответствующей химической реакции может быть представлено в следующем виде: Ca 10 [(P 6– x C x O 24 ](OH) 2–x + 10HCO 3 – + xCa2+ + (2–x)H+= (10+x)Ca[CO 3 ] + (6–x)HPO 3 + (8–x)OH.
Костные остатки пещерных львов и медведей по валовому химическому составу (табл. 6) в целом близки, за исключением некоторых компонентов. Так, в костях медведей обнаруживается в два раза больше SiO 2 , в три раза больше Fe 2 O 3 и раз в 30 больше MnO. Пересчет этих данных на нормативноминеральный состав показывает, что содержание иллювиированных в кость минеральных загрязнений колеблется в пределах 1–7 мол. % у львов и 0.8–17 мол. % у медведей (табл. 7). Этот результат разительно отличается от данных по костям, фоссилизирован-ным во внепещерных обстановках. В последних содержание иллювиированной минеральной примеси может достигать 50 мол. % и даже более (Silaev et al., 2917; Силаев и др., 2019).
По содержанию нормативных минералов в иллювиированных загрязнениях исследованные кости несколько различаются: в медвежьих костях больше кварца и слюды, раза в 4 больше Fe–Mn оксигидроксидов, но в 3.5–4 раза меньше хлоритов и почти нет пирита.
Таблица 4 . Химический (мас. %) и нормативно-минеральный (мол. %) составы смыва грунтов с костей иманайских пещерных львов
|
Компоненты |
492 |
622 |
823 |
2411 |
2847 |
2974 |
2975 |
Статистика |
|
SiO 2 |
35.7 |
28.78 |
31.77 |
31.19 |
33.56 |
36.07 |
28.83 |
32.27 ± 2.98 |
|
TiO 2 |
0.35 |
0.56 |
0.67 |
0.41 |
0.34 |
0.35 |
0.33 |
0.43 ± 0.13 |
|
Al 2 O 3 |
7.75 |
6.21 |
6.87 |
9.43 |
7.6 |
8.13 |
7.27 |
7.51 ± 1.02 |
|
Fe 2 O 3 |
2.23 |
4.99 |
5.29 |
3.21 |
2.5 |
2.42 |
2.24 |
3.27 ± 1.32 |
|
Cr 2 O 3 |
Н. о. |
0.09 |
Н. о. |
Н. о. |
0.03 |
0.03 |
Н. о. |
0.02 ± 0.03 |
|
NiO |
« |
0.05 |
« |
0.03 |
0.02 |
0.02 |
0.02 |
0.02 ± 0.02 |
|
ZnO |
0.08 |
0.25 |
0.24 |
0.12 |
0.17 |
0.09 |
0.11 |
0.15 ± 0.07 |
|
MnO |
0.08 |
0.16 |
0.15 |
0.14 |
0.09 |
0.09 |
0.1 |
0.12 ± 0.03 |
|
MgO |
1.94 |
1.29 |
1.22 |
3.1 |
1.94 |
2.09 |
1.81 |
1.91 ± 0.62 |
|
CaO |
36.2 |
43.83 |
39.18 |
37.27 |
34.79 |
32.68 |
41.18 |
37.88 ± 3.82 |
|
SrO |
Н. о. |
0.05 |
0.03 |
Н. о. |
0.02 |
0.02 |
0.01 |
0.02 ± 0.02 |
|
Na 2 O |
0.2 |
0.18 |
Н. о. |
0.16 |
0.21 |
0.25 |
0.18 |
0.17 ± 0.08 |
|
K 2 O |
0.98 |
1.16 |
1.2 |
1.37 |
0.99 |
1.05 |
0.95 |
1.1 ± 0.15 |
|
P 2 O 5 |
14.34 |
12.21 |
13.25 |
7.77 |
14.42 |
13.59 |
12.72 |
12.61 ± 2.28 |
|
SO 3 |
0.15 |
0.19 |
0.13 |
5.8 |
3.32 |
3.12 |
4.25 |
2.42 ± 2.29 |
|
Ca/P ат |
3.2 |
4.55 |
3.75 |
6.15 |
3.32 |
3.12 |
4.25 |
4.05 ± 1.07 |
|
Апатит |
35.19 |
29.91 |
35.41 |
18.13 |
36.54 |
34.56 |
26.12 |
30.84 ± 6.72 |
|
Кальцит |
14.72 |
34.53 |
19.18 |
40.44 |
30.25 |
16.8 |
39.58 |
27.93 ± 10.92 |
|
Кварц |
25.39 |
16.93 |
22.53 |
14.1 |
23.51 |
28.68 |
15.05 |
20.88 ± 5.57 |
|
Полевые шпаты |
1.71 |
1.43 |
Не опр. |
1.17 |
1.78 |
2.18 |
1.37 |
1.38 ± 0.69 |
|
Слюда |
7.64 |
8.54 |
11.0 |
9.38 |
7.73 |
8.78 |
6.67 |
8.53 ± 1.4 |
|
Хлориты |
15.2 |
8.42 |
11.73 |
16.69 |
0.08 |
8.83 |
11.1 |
10.29 ± 5.44 |
|
Пирит |
0.15 |
0.24 |
0.15 |
0.09 |
0.11 |
0.17 |
0.11 |
0.15 ± 0.05 |
Таблица 5 . Химический (мас. %) и нормативно-минеральный (мол. %) состав смыва грунтов с костей иманайских пещерных медведей
|
Компоненты |
1/1 |
2 |
3/1 |
4/1 |
7 |
8 |
9 |
(10+11)/1 |
12 |
Статистика |
|
SiO 2 |
37.08 |
32.0 |
33.9 |
34.52 |
35.37 |
37.83 |
39.83 |
42.03 |
50.97 |
38.17 ± 7.7 |
|
TiO 2 |
0.38 |
0.32 |
0.37 |
0.38 |
0.43 |
0.46 |
0.47 |
0.59 |
0.77 |
0.46 ± 0.14 |
|
ZrO2 |
Н. о. |
Н. о. |
0.02 |
Н. о. |
Н. о. |
0.02 |
0.02 |
0.02 |
Н. о. |
0.01 ± 0.01 |
|
Al 2 O 3 |
8.9 |
7.17 |
8.24 |
8.1 |
8.18 |
8.83 |
9.43 |
10.43 |
13.9 |
9.24 ± 1.97 |
|
Fe 2 O 3 |
2.66 |
1.92 |
2.49 |
2.54 |
2.51 |
2.74 |
2.85 |
3.34 |
4.13 |
2.8 ± 0.62 |
|
Cr 2 O 3 |
0.04 |
Н. о. |
0.03 |
0.03 |
0.03 |
0.03 |
0.03 |
0.03 |
0.03 ± 0.01 |
|
|
NiO |
0.02 |
« |
0.01 |
0.02 |
0.02 |
0.02 |
0.02 |
0.02 |
0.03 |
0.02 ± 0.01 |
|
CuO |
0.01 |
« |
0.01 |
Н. о. |
Н. о. |
Н. о. |
Н. о. |
Н. о. |
Н. о. |
0.002 ± 0.004 |
|
ZnO |
0.08 |
0.09 |
0.08 |
0.06 |
0.07 |
0/05 |
0.09 |
0.05 |
0.05 |
0.07 ± 0.02 |
|
MnO |
0.08 |
0.06 |
0.09 |
0.06 |
0.09 |
0.18 |
0.13 |
0.12 |
0.15 |
0.11 ± 0.04 |
|
MgO |
2.48 |
2.12 |
1.51 |
2.29 |
1.97 |
1.59 |
2.11 |
2.0 |
2.24 |
2.03 ± 0.32 |
|
CaO |
32.73 |
39.66 |
36.57 |
38.72 |
38.51 |
32.63 |
30.08 |
31.4 |
20.62 |
33.44 ± 5.95 |
|
SrO |
0.02 |
Н. о.. |
0.02 |
0.02 |
0.01 |
0.02 |
0.02 |
0.02 |
Н. о. |
0.01 ± 0.01 |
|
Na 2 O |
0.21 |
0.29 |
0.2 |
0.17 |
0.16 |
0.2 |
0.19 |
0.14 |
0.1 |
0.18 ± 0.05 |
|
K 2 O |
1.15 |
0.99 |
0.95 |
0.99 |
0.97 |
1.0 |
1.14 |
1.17 |
1.53 |
1.1 ± 0.18 |
|
P 2 O 5 |
13.99 |
15.24 |
15.4 |
12.01 |
11.6 |
14.27 |
13.49 |
8.58 |
5.44 |
12.22 ± 3.31 |
|
SO 3 |
0.17 |
0.14 |
0.11 |
0.09 |
0.08 |
0.13 |
0.1 |
0.06 |
0.07 |
0.11 ± 0.04 |
|
Ca/P ат |
2.97 |
3.3 |
3.01 |
4.09 |
4.21 |
2.9 |
2.83 |
4.64 |
4.81 |
3.64 ± 0.79 |
|
Апатит |
34.72 |
37.76 |
38.42 |
29.67 |
28.68 |
35.72 |
33.60 |
21.2 |
13.49 |
30.36 ± 8.29 |
|
Кальцит |
11.21 |
16.37 |
12.95 |
20.63 |
21.12 |
10.72 |
9.25 |
18.67 |
12.64 |
14.84 ± 4.47 |
|
Кварц |
22.5 |
19.93 |
20.26 |
21.06 |
21.78 |
23.31 |
24.39 |
24.81 |
29.33 |
23.04 ± 2.91 |
|
Полевые шпаты |
2.0 |
2.76 |
1.92 |
1.61 |
1.52 |
1.89 |
1.8 |
1.32 |
0.94 |
1.75 ± 0.51 |
|
Слюда |
10.07 |
8.66 |
8.35 |
8.63 |
8.44 |
8.83 |
10.03 |
10.2 |
13.32 |
9.61 ± 1.58 |
|
Хлориты |
19.5 |
14.52 |
18.1 |
18.4 |
18.46 |
19.53 |
20.93 |
23.8 |
30.28 |
20.39 ± 4.45 |
Примечание. Н.о. – не обнаружен.
Следовательно, получается, что кости пещерных львов и медведей фоссилизирова-лись в нетождественных Eh-условиях – кости львов в какой-то период пребывали в более восстановительных условиях (образование пирита), а кости медведей, напротив, в более окислительных (оксигидроксидное омарганцевание).
Важнейшей особенностью фоссилизации исследуемых костей является их кальцитиза-ция, т. е. замещение исходного биоапатита карбонатами. Это коррелируется с отмеченной выше фосфатизацией пещерноэлювиальных грунтов, обусловленной при-вносом в них замещенной в костях части фосфатного вещества. Такая картина вполне закономерна, поскольку костный детрит иманайских плейстоценовых млекопитающих был захоронен в карстогенной пещере. Прямое определение карбонатности костей осуществлялось химико-аналитическим методом. Анализ показал, что в костном детрите пещерных львов содержание СО2 варьируется в пределах 6.5–17.5 мас. %, возрастая в последовательности: (зубы, клыки) < сеса- мовидные кости < позвонки < (рёбра, фаланги). В среднем это составляет 12.29 ± 3.41 мас. %. В костном детрите пещерных медведей содержание СО2 колеблется в пределах 6—17.5 мас. %, увеличиваясь в направлении от зубов и клыков к фрагментам черепа, челюсти, атланта, далее к бедренной кости, фалангам и костям метаподия. Средние данные составляют (10.66 ± 4.24) мас. %. Согласно расчетам, содержания СО2 в пещерных костях прямо и очень сильно (r = 0.82—0.88) коррелируются с атомным отношением Ca/P, из чего следует, что кальцитизация пещерных костей имеет наложенный, эпигенетический характер.
Из результатов сравнительного анализа видно, что иманайские пещерные кости по содержанию СО 2 значительно преобладают над примерно одновозрастными костями плейстоценовых млекопитающих, фоссили-зированными во внепещерных обстановках (рис. 11), что, очевидно, обусловлено нетронутостью последних эпигенетической каль-цитизацией.
Таблица 6. Химический состав костей пещерных львов (1–14) и медведей (15–29), мас. %
|
№ п/п |
№ обр |
SiO 2 |
TiO 2 |
Al2O 3 |
Fe 2 O 3 |
MnO |
MgO |
CaO |
SrO |
ZnO |
NiO |
Na 2 O |
K 2 O |
P 2 O 5 |
SO 3 |
Ca/P ат |
|
1 |
622 |
1.42 |
н. о. |
0.44 |
0.22 |
н. о. |
0.79 |
64.12 |
0.04 |
0.07 |
н. о. |
0.49 |
0.07 |
32.26 |
0.08 |
2.52 |
|
2 |
823 |
0.90 |
« |
0.44 |
0.13 |
« |
0.76 |
60.20 |
0.04 |
0.07 |
« |
0.58 |
0.05 |
36.70 |
0.13 |
2.08 |
|
3 |
2847 |
0.56 |
« |
0.17 |
0.11 |
« |
0.75 |
63.10 |
0.04 |
0.08 |
« |
0.55 |
н. о. |
34.52 |
0.12 |
2.32 |
|
4 |
492 |
1.01 |
« |
0.28 |
0.13 |
« |
0.72 |
62.27 |
0.03 |
0.04 |
« |
0.59 |
0.03 |
34.78 |
0.12 |
2.27 |
|
5 |
2869 |
1.73 |
« |
0.59 |
0.20 |
« |
0.80 |
62.25 |
0.05 |
0.10 |
« |
0.35 |
0.06 |
33.75 |
0.12 |
2.33 |
|
6 |
2803 |
1.87 |
« |
0.57 |
0.19 |
« |
0.81 |
60.69 |
0.05 |
0.06 |
« |
0.30 |
0.06 |
35.28 |
0.12 |
2.18 |
|
7 |
2411 |
0.60 |
« |
0.28 |
0.14 |
« |
0.83 |
63.57 |
0.03 |
0.06 |
« |
0.53 |
0.04 |
33.82 |
0.10 |
2.38 |
|
8 |
2871 |
2.43 |
« |
0.68 |
0.36 |
0.04 |
0.80 |
62.97 |
0.04 |
0.10 |
« |
0.49 |
0.08 |
31.93 |
0.08 |
2.50 |
|
9 |
3111 |
1.16 |
« |
0.47 |
0.17 |
н. о. |
0.93 |
63.55 |
0.05 |
0.09 |
« |
0.57 |
0.05 |
32.78 |
0.18 |
2.45 |
|
10 |
2987 |
1.33 |
« |
0.49 |
0.18 |
« |
0.69 |
58.93 |
0.04 |
0.05 |
« |
0.45 |
0.07 |
37.57 |
0.20 |
1.99 |
|
11 |
3046 |
0.45 |
« |
0.18 |
0.11 |
н. о. |
0.78 |
61.82 |
0.03 |
0.09 |
« |
0.51 |
н. о. |
35.87 |
0.16 |
2.18 |
|
12 |
2974 |
3.50 |
0.07 |
1.05 |
0.42 |
0.02 |
0.94 |
61.48 |
0.05 |
0.10 |
« |
0.54 |
0.13 |
31.57 |
0.13 |
2.47 |
|
13 |
2975 |
0.33 |
н. о. |
0.12 |
0.06 |
н. о. |
0.81 |
59.94 |
0.05 |
0.08 |
« |
0.34 |
0.02 |
38.13 |
0.12 |
1.99 |
|
14 |
3038 |
0.54 |
« |
0.20 |
0.08 |
« |
0.75 |
61.13 |
0.05 |
0.13 |
« |
0.55 |
0.02 |
36.41 |
0.13 |
2.13 |
|
Среднее |
1.27 |
0.01 |
0.43 |
0.18 |
0.004 |
0.8 |
61.86 |
0.04 |
0.08 |
0 |
0.49 |
0.05 |
34.67 |
0.13 |
2.27 |
|
|
СКО |
0.88 |
0.02 |
0.25 |
0.1 |
0.01 |
0.07 |
1.55 |
0.01 |
0.02 |
0 |
0.09 |
0.03 |
2.10 |
0.03 |
0.18 |
|
|
Коэф.вариации, % |
69 |
200 |
58 |
56 |
250 |
9 |
2.5 |
25 |
25 |
0 |
18 |
60 |
6 |
23 |
8 |
|
|
15 |
ИМ-1/1 |
1.76 |
н. о. |
0.53 |
1.95 |
0.77 |
1.06 |
57.03 |
н. о. |
н. о. |
н. о. |
0.12 |
0.08 |
36.32 |
0.38 |
1.99 |
|
16 |
ИМ-1/2 |
4.2 |
0.06 |
1.20 |
0.45 |
0.04 |
0.98 |
57.26 |
0.05 |
0.08 |
« |
0.39 |
0.15 |
35.03 |
0.11 |
2.33 |
|
17 |
ИМ-2 |
1.26 |
н.о. |
0.38 |
0.16 |
0.01 |
0.92 |
63.33 |
0.03 |
0.09 |
« |
0.56 |
0.05 |
33.07 |
0.14 |
2.43 |
|
18 |
ИМ-3/1 |
1.89 |
« |
0.50 |
0.22 |
0.02 |
0.79 |
57.98 |
0.02 |
0.08 |
« |
0.66 |
0.07 |
37.70 |
0.07 |
1.95 |
|
19 |
ИМ-3/2 |
11.21 |
0.15 |
2.99 |
0.96 |
0.05 |
1.14 |
55.25 |
0.03 |
0.10 |
« |
0.41 |
0.34 |
27.25 |
0.12 |
2.57 |
|
20 |
ИМ-4/1а |
1.56 |
н. о. |
0.44 |
0.26 |
0.01 |
0.65 |
58.58 |
0.03 |
0.08 |
« |
0.64 |
0.07 |
37.57 |
0.11 |
2.02 |
|
21 |
ИМ-4/1б |
1.53 |
« |
0.48 |
0.14 |
0.01 |
0.76 |
56.69 |
0.04 |
0.03 |
« |
0.65 |
0.06 |
39.46 |
0.15 |
1.82 |
|
22 |
ИМ-4/2 |
1.15 |
« |
н. о. |
0.24 |
0.01 |
0.79 |
59.83 |
0.06 |
0.09 |
« |
0.48 |
0.05 |
37.19 |
0.11 |
2.07 |
|
23 |
ИМ-8 |
6.42 |
« |
0.08 |
0.53 |
0.11 |
0.88 |
58.56 |
0.05 |
0.05 |
« |
0.46 |
0.18 |
32.57 |
0.11 |
2.28 |
|
24 |
ИМ-9 |
0.57 |
« |
н. о. |
0.46 |
0.05 |
0.77 |
58.44 |
0.04 |
0.11 |
« |
0.44 |
0.02 |
39.02 |
0.08 |
1.90 |
|
25 |
ИМ-10 |
8.58 |
« |
0.17 |
1.02 |
0.01 |
1.35 |
58.69 |
0.05 |
0.08 |
« |
0.42 |
0.23 |
29.30 |
0.10 |
2.52 |
|
26 |
ИМ-(10+11)/1 |
0.44 |
« |
н. о. |
0.09 |
0.03 |
0.75 |
60.84 |
0.05 |
0.11 |
« |
0.47 |
0.01 |
37.14 |
0.07 |
2.08 |
|
27 |
ИМ-(10+11)/2 |
1.04 |
« |
« |
0.13 |
0.01 |
0.80 |
58.98 |
0.05 |
0.07 |
« |
0.43 |
0.03 |
38.37 |
0.09 |
1.95 |
|
28 |
ИМ-12м |
1.65 |
« |
0.50 |
0.17 |
0.20 |
0.81 |
60.39 |
0.05 |
0.09 |
0.02 |
0.41 |
0.05 |
35.58 |
0.10 |
2.15 |
|
29 |
ИМ-12ф |
0.79 |
« |
0.32 |
0.12 |
0.40 |
0.77 |
60.69 |
0.05 |
0.05 |
н. о. |
0.43 |
0.04 |
36.23 |
0.11 |
2.12 |
|
Среднее |
2.94 |
0.01 |
0.51 |
0.46 |
0.12 |
0.88 |
58..84 |
0.04 |
0.07 |
0.005 |
0.46 |
0.1 |
35.45 |
0.12 |
2.14 |
|
|
СКО |
3.25 |
0.04 |
0.76 |
0.51 |
0.21 |
0.18 |
1.99 |
0.02 |
0.03 |
0.02 |
0.13 |
0.09 |
3.52 |
0.07 |
0.23 |
|
|
Коэф.вариации, % |
110 |
400 |
149 |
111 |
175 |
20 |
3 |
50 |
43 |
400 |
28 |
90 |
10 |
58 |
11 |
|
Таблица 7 . Нормативно-минеральный состав костей пещерных львов (1–14) и медведей (15–29), мол. %
|
№ п/п |
№ обр. |
Апатит + кальцит |
Кварц |
Полевые шпаты |
Слюда |
Хлориты |
Пирит |
Fe-Mn оксигидроксиды |
|
1 |
622 |
96.58 |
0.95 |
Не опр. |
0.54 |
1.51 |
0.13 |
0.29 |
|
2 |
823 |
97.62 |
0.35 |
« |
0.35 |
1.42 |
0.2 |
0.06 |
|
3 |
2847 |
98.28 |
0.31 |
« |
Не опр. |
1.18 |
0.18 |
0.05 |
|
4 |
492 |
97.7 |
0.64 |
« |
0.21 |
1.2 |
0.18 |
0.07 |
|
5 |
2869 |
96.49 |
0.94 |
« |
0.42 |
1.83 |
0.18 |
0.14 |
|
6 |
2803 |
96.37 |
1.11 |
« |
0.42 |
1.79 |
0.18 |
0.13 |
|
7 |
2411 |
98.0 |
0.24 |
« |
0.28 |
1.24 |
0.15 |
0.09 |
|
8 |
2871 |
95.78 |
1.82 |
« |
0.35 |
1.61 |
0.12 |
0.32 |
|
9 |
3111 |
97.03 |
0.53 |
« |
0.35 |
1.74 |
0.27 |
0.08 |
|
10 |
2987 |
97.08 |
0.7 |
« |
0.49 |
1.4 |
0.30 |
0.08 |
|
11 |
3046 |
98.31 |
0.18 |
« |
Не опр. |
1.24 |
0.24 |
0.03 |
|
12 |
2974 |
93.27 |
1.31 |
2.71 |
0.91 |
1.24 |
0.2 |
0.36 |
|
13 |
2975 |
99.05 |
0.18 |
Не опр. |
0.14 |
0.45 |
0.18 |
Не опр. |
|
14 |
3038 |
98.31 |
0.27 |
« |
0.1 |
1.11 |
0.19 |
0.02 |
|
Среднее |
97.13 |
0.68 |
0.19 |
0.35 |
1.35 |
0.19 |
0.12 |
|
Окончание табл. 7
|
№ п/п |
№ обр. |
Апатит + кальцит |
Кварц |
Полевые шпаты |
Слюда |
Хлориты |
Пирит |
Fe-Mn оксигидроксиды |
|
СКО |
1.45 |
0.49 |
0.72 |
0.23 |
0.35 |
0.12 |
0.12 |
|
|
Коэф. вариации, % |
1.5 |
72 |
379 |
66 |
26 |
63 |
100 |
|
|
15 |
1/1 |
94.59 |
0.64 |
1.21 |
0.65 |
0.65 |
Не опр. |
2.26 |
|
16 |
1/2 |
92.75 |
1.26 |
3.85 |
1.37 |
0.4 |
« |
0.37 |
|
17 |
2 |
97.38 |
Не опр. |
2.15 |
0.34 |
Не опр. |
« |
0.13 |
|
18 |
3/1 |
96.66 |
0.13 |
2.56 |
0.46 |
« |
« |
0.19 |
|
19 |
3/2 |
83.12 |
5.45 |
4.0 |
3.06 |
3.61 |
« |
0.77 |
|
20 |
4/1а |
97.31 |
0.57 |
1.26 |
0.65 |
Не опр. |
« |
0.21 |
|
21 |
4/б |
96.98 |
Не опр. |
2.5 |
0.4 |
« |
« |
0.12 |
|
22 |
4/2 |
99.7 |
« |
Не опр. |
Не опр. |
« |
0.13 |
0.17 |
|
23 |
8 |
93.13 |
5.95 |
« |
0.46 |
« |
Не опр. |
0.47 |
|
24 |
9 |
99.01 |
0.59 |
« |
Не опр. |
« |
« |
0.4 |
|
25 |
10 |
89.91 |
8.19 |
1.11 |
« |
« |
« |
0.79 |
|
26 |
(10+11)/1 |
99.46 |
0.45 |
Не опр. |
« |
« |
« |
0.09 |
|
27 |
(10+11)/2 |
98.82 |
1.07 |
« |
« |
« |
« |
0.11 |
|
28 |
12м |
96.94 |
0.29 |
1.99 |
0.47 |
« |
« |
0.31 |
|
29 |
12ф |
99.19 |
Не опр. |
0.34 |
0.28 |
« |
« |
0.19 |
|
Среднее |
95.66 |
1.64 |
1.4 |
0.54 |
0.31 |
0.01 |
0.44 |
|
|
СКО |
4.48 |
2.62 |
1.4 |
0.79 |
0.93 |
0.03 |
0.55 |
|
|
Коэф. вариации,% |
5 |
160 |
100 |
146 |
300 |
300 |
125 |
|
Непосредственное фазовое присутствие в исследуемых костях новообразованного карбоната доказывается методом рентгенолюминесцентной спектроскопии. Проведенные предварительные исследования показали, что для ископаемых костей, захороненных во внепещерных обстановках, рентгенолюми-несценция не характерна. В пещерном же костном детрите она, напротив, всегда возбуждается, достигая значительной интенсивности в полосе с максимумом 630 нм.
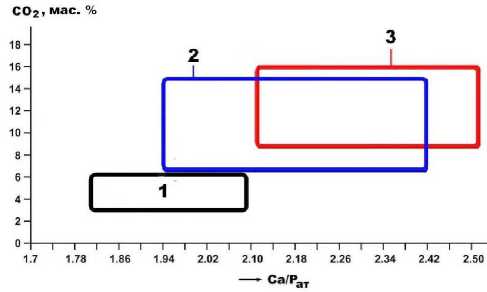
Рис. 11. Содержание СО2 в ископаемых костях как критерий степени их кальцитизации в процессе фоссилизации: 1 – мамонтовая фауна, фоссилизированная во внепещерных обстановках (Печорское Приуралье, Омское Прииртышье); 2, 3 – кости из пещеры Иманай, соответственно медвежьи и львиные
Последнее соответствует излучению на ионах-люминофорах Mn2+ в структуре кальцита.
Среднестатистическая нормализованная интенсивность марганцевой рентгенолюми-несценции костей иманайских пещерных львов и медведей составляет соответственно (7.88 ± 5.15) и (10.12 ± 5.44) отн. ед. Обобщенный анализ особенностей нормативноминерального состава костеносных грунтов, смывов с костей и иллювиированных в кости минеральных загрязнений приводит к следующим выводам (рис. 12).
Собственно пещерные грунты характеризуются промежуточным кварц-полевошпат-слюдисто-хлоритовым составом. Смыв с костей львов практически совпадает по нормативно-минеральному составу с грунтами, а смыв с костей медведей заметно обогащен относительно грунтов кварцем и полевыми шпатами. Иллювиированная в кости минеральная примесь заметно отклоняется от состава грунтов, причем для львиных и медвежьих костей разнонаправленно. В львиных костях она сильно обогащена хлоритами за счет кварца и полевых шпатов, а в медвежьих, напротив, – кварцем и полевыми шпатами за счет хлоритов. Примечательно выглядит существенное и разнонаправленное расхождение по нормативно-минеральному со- ставу собственно костей и смывов с них. Так, смыв с костей львов демонстрирует обогащение кварцем и полевыми шпатами, а кости содержат филлосиликаты. Анализ медвежьих костей, наоборот, показывает, что смывы имеют существенно слюдисто-хлоритовый состав, а кости – преимущественно кварц-полевошпатовый.
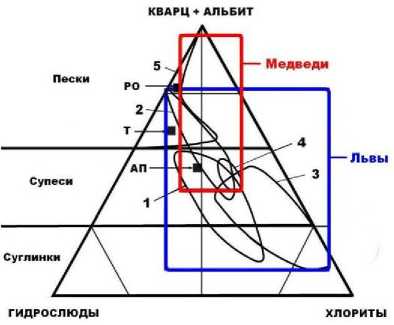
Рис. 12. Нормативно-минеральный состав костеносных грунтов, смывов с пещерных костей и иллювиированных в кости минеральных загрязнений: 1 – грунты; 2, 3 – соответственно смывы и кости львов; 4, 5 – соответственно смывы и кости медведей. Черные квадраты – средние составы речных отложений (РО), тиллов (Т) и псаммоалевропелитовых осадков (АП)
Получается, что минеральный состав ил-лювиированных загрязнений выявляет различие костей львов и медведей по характеру их проницаемости для микроминеральных примесей. В целом результаты проведенных исследований указывают на автохтонность пещерных костей по отношению к костеносным грунтам и выявляют при этом нетожде-ственность костей пещерных львов и медведей по этим свойствам и, следовательно, разную степень фоссилизации.
Микроэлементы
В составе исследуемых пещерных костей обнаружены 50 микроэлементов (табл. 8, 9), в том числе (в скобках доля в %, первые цифры – львы, вторые – медведи) 10 элемен-тов-эссенциалов – Э (36.9 ± 3.9 и 34.6 ± 9.1), 19 физиологически-активных элементов – ФА (60.9 ± 5.6 и 63.6 ± 9.4) и 21 элементов-антибионтов – АБ (2.2 ± 4.6 и 1.8 ± 3.0). Элементы первой группы в костях наследу- ются от живого организма, в котором они, как известно, отвечают за перенос кислорода, фиксацию азота, стимулируют метаболизм железа, углеводородный обмен, способствуют образованию ферментов, формированию структуры клеток, дифференциации и стабилизации клеточных мембран (Войнар, 1960). Вторая группа объединяет элементы, которые могут быть по происхождению как унаследованными от организма, так и ксеногенными. Третья группа включает элементы, которые по происхождению являются практически нацело ксеногенно-абиотическими, будучи заимствованными из среды захоронения. Обогащение такими элементами костей происходит не только вследствие иллю-виирования в кости вещества грунтов, но и по ионообменному механизму, когда кристаллохимически активные элементы из окружающей среды замещают в костном биоапатите кальций и фосфор.
Отношение групповых содержаний элементов Э/АБ можно рассматривать как критерий оценки степени фоссилизации ископаемых костей. Так, в живом, например, бактериальном веществе вследствие громадного превосходства элементов-эссенциалов над антибионтами величина упомянутого отношения лежит в пределах 110–300 (Кокин и др., 2020).
В костном детрите плейстоценовых животных, захороненных во внепещерных обстановках, значение такого отношения колеблется в интервале 0.5–2. В иманайских же костях рассматриваемая Э/АБ-пропорция имеет промежуточные, но сильно сдвинутые в сторону живого вещества значения, варьируясь у костей львов и медведей в интервалах соответственно 20–158 (61.39 ± 40.15) и 17–79 (40.83 ± 19.51).
Очевидно, что полученные по иманай-ским костям результаты обусловлены, прежде всего, очень низким содержанием в них АБ-элементов. Следовательно, фоссилизация костей в пещерных условиях принципиально отличается от таковой во внепещерных обстановках крайне низкой степенью эпигенетического обогащения микроэлементами вмещающей геологической среды. На это указывают и значения индикаторных геохимических модулей, которые для внепещер-ных костей и костей иманайских львов и ме-
Таблица 8. Микроэлементы в костном детрите иманайских пещерных львов, г/т
|
Элементы 2803 |
3046 |
3111 |
2987 |
2975 |
491 |
622 |
823 |
2847 |
2869 |
2871 |
2411 |
|
|
Элементы-эссенциалы (Э) |
||||||||||||
|
Ag |
0.04 |
0.047 |
0.109 |
0.075 |
0.02 |
0.029 |
0.025 |
0.034 |
0.031 |
0.021 |
0.032 |
0.053 |
|
As |
1.242 |
1.308 |
0.915 |
1.015 |
1.539 |
1.522 |
1.289 |
1.366 |
1.265 |
0.889 |
1.381 |
1.198 |
|
Bi |
Не обн. |
Не обн. |
0.011 |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
|
Cd |
1.013 |
1.142 |
2.254 |
1.662 |
1.293 |
0.815 |
1.379 |
2.448 |
1.516 |
1.694 |
2.439 |
2.152 |
|
Мо |
0.314 |
0.125 |
0.185 |
0.431 |
0.309 |
0.18 |
0.159 |
0.425 |
0.093 |
0.147 |
0.17 |
0.402 |
|
Se |
4.446 |
1.737 |
1.968 |
1.88 |
1.963 |
2.164 |
1.327 |
2.183 |
2.053 |
1.827 |
1.454 |
89.501 |
|
Rb |
1.138 |
0.176 |
0.511 |
0.988 |
0.212 |
0.705 |
1.374 |
0.485 |
0.683 |
1.069 |
2.011 |
0.948 |
|
Pb |
0.761 |
0.449 |
0.709 |
1.016 |
0.212 |
0.417 |
0.52 |
0.575 |
0.35 |
0.606 |
1.076 |
0.513 |
|
Th |
0.158 |
0.038 |
0.053 |
0.055 |
0.019 |
0.082 |
0.153 |
0.087 |
0.08 |
0.068 |
0.268 |
0.106 |
|
Zn |
237.794 |
306.255 |
308.027 |
306.932 |
307.708 |
267.367 |
271.816 |
359.712 |
306.29 |
381.38 |
324.385 |
287.60 |
|
Сумма Э |
246.906 |
311.277 |
314.742 |
314.054 |
313.275 |
273.281 |
278.042 |
367.315 |
312.361 |
387.701 |
333.216 |
382.473 |
|
Элементы физиологически-активные (ФА) |
||||||||||||
|
Ba |
167.204 |
128.802 |
155.683 |
150.31 |
132.745 |
159.802 |
146.98 |
180.742 |
119.331 |
130.435 |
150.599 |
147.425 |
|
V |
12.077 |
9.356 |
6.228 |
8.562 |
10.786 |
8.302 |
8.44 |
8.116 |
8.551 |
8.331 |
10.761 |
9.049 |
|
Ga |
0.796 |
0.613 |
0.596 |
0.836 |
0.762 |
0.685 |
0.705 |
0.726 |
0.667 |
0.695 |
0.775 |
0.687 |
|
Hf |
0.061 |
0.039 |
0.045 |
0.044 |
0.02 |
0.042 |
0.047 |
0.073 |
0.046 |
0.061 |
0.088 |
0.07 |
|
Y |
1.596 |
0.451 |
0.969 |
1.238 |
0.194 |
0.575 |
1.419 |
0.718 |
0.678 |
1.227 |
1.519 |
1.66 |
|
Co |
0.82 |
0.88 |
0.681 |
1.02 |
1.013 |
0.962 |
1.26 |
0.847 |
0.825 |
1.476 |
2.98 |
0.852 |
|
Cu |
33.124 |
37.699 |
28.029 |
21.302 |
34.241 |
15.723 |
30.97 |
87.723 |
35.27 |
29.338 |
31.529 |
34.522 |
|
Li |
1.084 |
0.685 |
0.533 |
0.872 |
1.65 |
1.194 |
1.191 |
1.243 |
1.004 |
1.049 |
1.469 |
0.894 |
|
Mn |
37.477 |
70.234 |
27.742 |
87.631 |
28.404 |
39.572 |
59.926 |
51.908 |
16.884 |
39.687 |
111.211 |
24.966 |
|
Ni |
23.085 |
25.301 |
23.283 |
25.62 |
20.752 |
19.453 |
26.685 |
22.736 |
25.966 |
23.238 |
30.338 |
22.672 |
|
Sb |
0.403 |
0.372 |
0.376 |
0.441 |
0.309 |
0.378 |
0.433 |
0.404 |
0.328 |
0.238 |
0.407 |
0.294 |
|
Sn |
0.056 |
0.022 |
0.011 |
0.006 |
0.02 |
0.086 |
0.047 |
0.095 |
0.131 |
0.015 |
0.053 |
0.031 |
|
Sr |
228.767 |
211.015 |
10.426 |
216.86 |
232.45 |
226.727 |
210.053 |
245.053 |
209.115 |
226.741 |
190.194 |
199.268 |
|
Ti |
3.435 |
32.778 |
8.922 |
13.112 |
4.773 |
11.655 |
15.296 |
11.768 |
9.382 |
11.545 |
24.1 |
10.441 |
|
Cs |
0.084 |
0.012 |
0.043 |
0.077 |
0.015 |
0.049 |
0.095 |
0.032 |
0.047 |
0.073 |
0.131 |
0.072 |
|
Zr |
3.183 |
2.252 |
3.359 |
3.448 |
0.979 |
2.037 |
2.875 |
5.321 |
2.665 |
3.642 |
5.267 |
5.807 |
|
Cr |
9.689 |
9.504 |
7.17 |
9.247 |
17.92 |
5.757 |
9.156 |
6.25 |
9.341 |
6.276 |
6.965 |
9.973 |
|
U |
20.754 |
24.62 |
16.999 |
16.446 |
28.298 |
49.631 |
14.731 |
7.304 |
12.741 |
12.577 |
4.863 |
17.959 |
|
Сумма ФА |
543.695 |
554.635 |
291.095 |
557.072 |
515.331 |
542.63 |
530.309 |
631.059 |
452.972 |
496.644 |
573.249 |
486.642 |
|
Элементы-антибионты (АБ) |
||||||||||||
|
B |
3.576 |
Не обн. |
Не обн. |
3.997 |
2.558 |
3.445 |
4.625 |
3.535 |
4.191 |
3.373 |
3.835 |
2.973 |
|
Nb |
0.042 |
0.066 |
0.042 |
0.046 |
0.025 |
0.04 |
0.04 |
0.034 |
0.025 |
0.031 |
0.077 |
0.032 |
|
Sc |
0.296 |
0.116 |
0.154 |
0.221 |
0.066 |
0.246 |
0.405 |
0.25 |
0.195 |
0.281 |
0.461 |
0.361 |
|
Ta |
Не обн. |
0.011 |
0.01 |
Не обн. |
0.025 |
0.009 |
Не обн. |
0.014 |
Не обн. |
Не обн. |
Не обн. |
Не обн. |
|
Te |
0.043 |
0.031 |
0.039 |
0.026 |
0.033 |
0.065 |
0.071 |
0.053 |
0.026 |
0.01 |
0.044 |
0.028 |
|
Tl |
3.891 |
0.655 |
0.825 |
0.911 |
0.277 |
1.210 |
1.293 |
0.576 |
0.943 |
2.612 |
1.166 |
1.17 |
|
W |
0.092 |
0.129 |
0.051 |
0.069 |
0.065 |
0.054 |
0.072 |
0.071 |
0.047 |
0.055 |
0.078 |
0.044 |
|
La |
0.888 |
0.199 |
0.36 |
0.624 |
0.134 |
0.396 |
0.835 |
0.397 |
0.43 |
0.725 |
1.181 |
0.611 |
|
Ce |
1.386 |
0.293 |
0.572 |
0.984 |
0.212 |
0.706 |
1.324 |
0.621 |
0.647 |
1.097 |
2.178 |
0.928 |
|
Pr |
0.207 |
0.042 |
0.086 |
0.14 |
0.03 |
0.086 |
0.191 |
0.081 |
0.094 |
0.162 |
0.272 |
0.138 |
|
Nd |
0.876 |
0.18 |
0.372 |
0.627 |
0.124 |
0.377 |
0.787 |
0.338 |
0.366 |
0.641 |
1.077 |
0.58 |
|
Sm |
0.188 |
0.034 |
0.096 |
0.126 |
0.026 |
0.077 |
0.168 |
0.075 |
0.067 |
0.151 |
0.238 |
0.133 |
|
Eu |
0.057 |
0.025 |
0.037 |
0.05 |
0.022 |
0.036 |
0.057 |
0.035 |
0.037 |
0.046 |
0.067 |
0.049 |
|
Gd |
0.215 |
0.05 |
0.105 |
0.169 |
0.031 |
0.094 |
0.204 |
0.098 |
0.094 |
0.168 |
0.265 |
0.174 |
|
Tb |
0.03 |
0.008 |
0.018 |
0.023 |
0.006 |
0.013 |
0.029 |
0.013 |
0.014 |
0.023 |
0.037 |
0.024 |
|
Dy |
0.21 |
0.047 |
0.114 |
0.189 |
0.023 |
0.073 |
0.184 |
0.086 |
0.085 |
0.145 |
0.214 |
0.181 |
|
Ho |
0.044 |
0.01 |
0.026 |
0.036 |
0.006 |
0.015 |
0.043 |
0.019 |
0.02 |
0.033 |
0.044 |
0.042 |
|
Er |
0.124 |
0.04 |
0.083 |
0.103 |
0.021 |
0.047 |
0.126 |
0.058 |
0.055 |
0.097 |
0.133 |
0.142 |
|
Tm |
0.02 |
0.005 |
0.011 |
0.015 |
0.002 |
0.006 |
0.017 |
0.008 |
0.007 |
0.014 |
0.017 |
0.017 |
|
Yb |
0.112 |
0.032 |
0.078 |
0.093 |
0.013 |
0.047 |
0.101 |
0.046 |
0.051 |
0.086 |
0.116 |
0.129 |
|
Lu |
0.016 |
0.006 |
0.014 |
0.016 |
0.003 |
0.006 |
0.013 |
0.008 |
0.007 |
0.013 |
0.016 |
0.021 |
|
Сумма Ln |
4.373 |
0.971 |
1.972 |
3.195 |
0.653 |
1.979 |
4.079 |
1.883 |
1.974 |
3.401 |
5.855 |
3.169 |
|
Сумма АБ |
12.313 |
1.979 |
3.093 |
3.465 |
3.702 |
7.048 |
10.585 |
6.416 |
7.401 |
9.763 |
11.516 |
7.777 |
|
Итого |
802.914 |
867.891 |
608.93 |
874.591 |
832.308 |
822.959 |
818.936 |
1004.734 |
772.734 |
894.108 |
917.981 |
876.892 |
|
Э/АБ |
20.05 |
157.29 |
101.76 |
90.64 |
84.62 |
38.77 |
26.27 |
57.25 |
42.21 |
39.71 |
28.94 |
49.18 |
Таблица 9. Микроэлементы в костном детрите иманайских пещерных медведей, г/т
Особое место среди микроэлементов в костях занимают лантаноиды, которые приобрели в настоящее время большую популярность в качестве геохимических критериев степени фоссилизации и относительного возраста ископаемого костного детрита (Herwartz et al., 2013). В нашем случае суммарное содержание лантаноидов в костях львов и медведей составляет соответственно 2.792 ± 1.505 и 5.657 ± 5.52 г/т, что в среднем в 70–140 раз уступает содержанию лантаноидов во внепещерных костях плейтоценовых животных, но в 10–20 тыс. раз превосходит содержание лантаноидов в живых организмах. Последовательность обогащения костей иманайских львов лантаноидами имеет следующий вид: (клык, сесамовидные кости) < позвонки < (ребра, фаланги). Аналогичная последовательность для медвежьих костей: (зубы, клыки, челюсть) < кости метаподия < (череп, бедро, атлант) < (ребро, фаланги).
Тренд нормированных на PAAS (средний постархейский австралийский сланец) концентраций лантаноидов в исследованных костях (рис. 13) демонстрирует их относительное обогащение средними лантаноидами (Sm, Eu, Gd) с проявлением, хотя и в несколько завуалированной форме, так называемого тетрадного эффекта (McLennan, 1994). Суть этого эффекта состоит в том, что в цериевой и иттриевой подгруппах лантаноидов в ходе фоссилизации костей сокращается содержание относительно легких элементов, склонных к выщелачиванию, но растет содержание относительно тяжелых элементов, склонных к образованию немигрирующих комплексных соединений. В результате возникает ступенчатая неоднородность распределения элементов с более или менее выраженными скачками концентраций на грани- цах так называемых тетрад: La–Nd (относительный минимум) → Pm–Gd (максимум) → Tb–Ho (минимум) → Er–Lu (максимум). Считается, что степень реализации тетрадного эффекта зависит от активности грунтовых вод и возраста ископаемых костей — в более древних и более фоссилизированных в условиях обводненных грунтов костях он выражен более отчетливо (Иванова, Никольский, 2015). Анализируя с этих позиций исследуемые пещерные кости, нельзя не прийти к выводу, что кости пещерных животных, несмотря на существенно более низкие концентрации лантаноидов, чем во внепещер-ных костях, демонстрируют тем не менее отчетливую тенденцию к обогащению средними лантаноидами и некоторые признаки тетрадного эффекта. Причем в львиных костях все это выражено отчетливее.
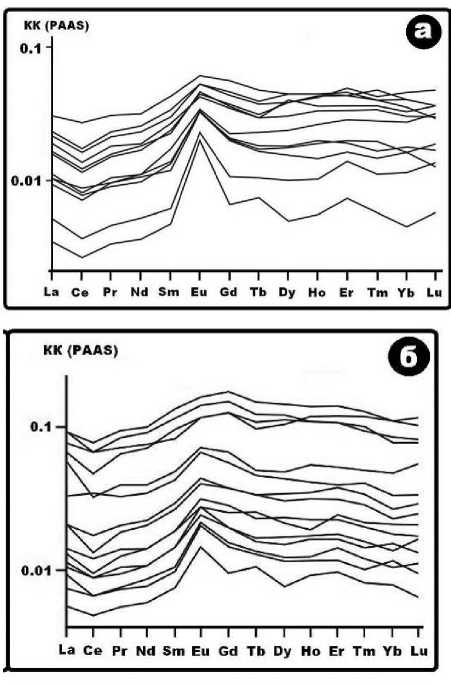
Рис. 13 . Тренды нормированных концентраций лантаноидов в иманайском костном детрите пещерных львов (а) и медведей (б)
Суммарная концентрация микроэлементов в исследуемых образцах пещерных костей варьируется для львов в диапазоне 0.841 ± 0.095, а для медведей в диапазоне 0.984 ± 0.509 г/т. Последовательности возрастания валовых концентраций элементов в анатомических разностях иманайских костей: кости львов – сесамовидные кости < клык < позвонки < ребро; кости медведей – (зубы, череп, ребро) < (клык, челюсть, атлант) < (бедро, кости метаподия, фаланги). На генеральной диаграмме обогащения микроэлементами (рис. 14) данные состава костей иманай-ских львов и медведей накладываются на область состава лишь минимально обогащенных микроэлементами зубов и бивней плейстоценовых животных, захороненных в условиях внепещерных обстановок. Все остальные внепещерные кости в 2–3 раза богаче микроэлементами. Таким образом, обнаруживается, что пещерные ископаемые кости характеризуются значительно меньшим валовым обогащением микроэлементами, чем внепещерные.
Костный биоапатит
Кости млекопитающих представляют собой органоминеральный композит, состоящий из органической матрицы (коллагенового белка) и производимого ею минерального вещества, которое в силу происхождения называется биоапатитом. Известно, что в ходе фоссилизации погребенных костей биоапатит подвергается вторичным изменениям, которые могут быть источником важной генетической информации.
На рентгеновских дифрактограммах биоапатита (рис. 15, 16) исследуемых костных образцов регистрируются все важнейшие отражения, характерные для этого минерала, но с разной степенью разрешения, возрастающей в направлении от костей скелета к дентину (зубной кости) и далее зубной эмали.
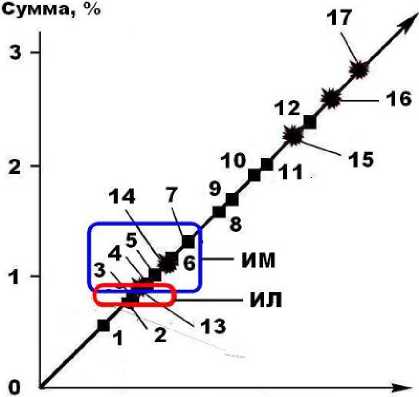
Продолжительность фоссилизации
Рис. 14. Суммарное содержание микроэлементов в костном детрите плейстоценовых млекопитающих из местонахождений в Печорском Приуралье (1–12), со стоянки Заозерье на Среднем Урале (13–17) и из пещеры Иманай на Южном Урале (ИЛ – львы, ИМ – медведи): 1 – мамонт, моляр; 2 – лошадь, резец; 3 – медведь пещерный, череп; 4, 5 – мамонт, зубы; 6, 7, 9 – мамонт, бивень; 8 – носорог шерстистый; 10 – мамонт, череп; 11 – мамонт, лопатка; 12 – северный олень, берцовая кость; 13–17 – костный детрит лошадей со стоянки Заозерье, соответственно моляры (13, 14), тазовая кость, ребро, пяточная кость
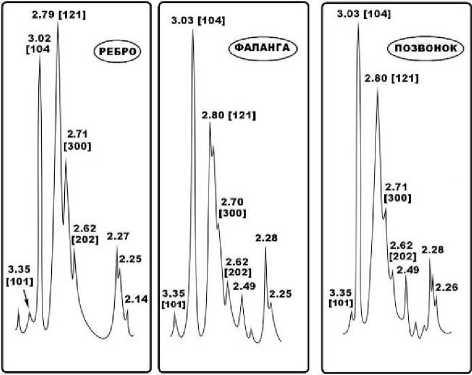
Рис. 15. Типичные рентгеновские дифракто-граммы для биоапатита в костях иманайских пещерных львов
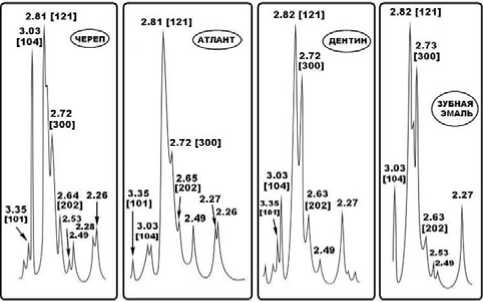
Рис. 16. Типичные рентгеновские дифракто-граммы, полученные для биоапатита костей иманайских пещерных медведей
В нашем случае индекс кристалличности (CI) биоапатита оценивался по отношению интенсивностей (I 300 +I 202 )/I 121 . Рост значений этого индекса свидетельствует об увеличении степени расщепления основного рентгеновского отражения в структуре апатита, а следовательно, указывает на возрастание степени окристаллизованности последнего.
Согласно полученным результатам, средние значения CI для биоапатита в костях львов и медведей составляют соответственно 0.74 ± 0.05 и 0.81 ± 0.17. В анатомических разностях костей львов эти значения увеличиваются в последовательности: сесамовидные кости (0.71 ± 0.04) < позвонки (0.72 ± 0.06) < ребра (0.76 ± 0.04) < клык (0.8). Аналогичный ряд для костей медведей имеет вид: ребро (0.51) < атлант (0.62) < фаланги (0.71 ± 0.01) < бедро (0.75) < череп (0.78) < зубы и клыки (0.96 ± 0.1). Эти данные в среднем близки к аналогичным результатам для внепещерных костей плейстоценовых животных, но раза в 1.5 уступают им по зубам и клыкам (Силаев и др., 2019).
Содержание основных ксеноминеральных примесей в исследуемых костях оценивалось по отношению главных отражений в кварце (101) и кальците (104) к основному отражению в биоапатите (121). Для костей пещерных львов и медведей значения «кварцевого» отношения I 101 /I 121 составляют соответственно 0.02 ± 0.04 и 0.13 ± 0.2, а значения «кальцитового» – 0.77 ± 0.45 и 0.76 ± 0.65. Следовательно, костный детрит львов и медведей по-разному обогащен упомянутыми примесями – в костях львов многократно меньше примеси кварца, но больше примеси кальцита.
В ИК спектрах исследуемых костей (рис. 17, 18) отчетливо регистрируются основные полосы ν 3 и ν 4 поглощения на РО 4 -группах, а также полосы ν 2 –ν 4 поглощения на группах СО 3 , изоморфно замещающих фосфатные анионы в биоапатите. Признаков поглощения на группах СО 3 , замещающих ОН-группы (валентные анионы), не обнаружено. Следовательно, биоапатит в костях иманай-ских животных, несмотря на кальцитизацию, является практически нацело карбонатапати-том В-типа.
Степень карбонизации биоапатита в рамках ИКС метода можно оценить отношением интенсивностей поглощения в основных полосах фосфатных и карбонатных групп – Iν3[PO4] / Iν3[CO3]. Величина этого отношения для исследованных костей пещерных львов и медведей составила соответственно 2.19 ± 0.83 и 2.55 ± 0.96, из чего следует, что костный биоапатит иманайских львов карбонизирован на 15–20 % сильнее, чем медвежий. Из приведенных данных следует также, что по степени карбонизации исследованные кости оказались неоднородными. В группе львиных костей степень карбонизации возрастает в последовательности: клык (3.49) < сесамовидные кости (3.19 ± 0.63) < позвонки (1.75 ± 0.54) < ребра (1.59 ± 0.4) < фаланги (1.58). Аналогичная последовательность для медвежьих костей имеет вид: зубы, клыки (3.47 ± 0.64) < череп (3.32) < бедро (2.76) < атлант (2.72) < челюсть (2.56) < кости мета-подия (1.88 < 0.49) < фаланги (1.62 ± 1.04) < ребро (1.36).
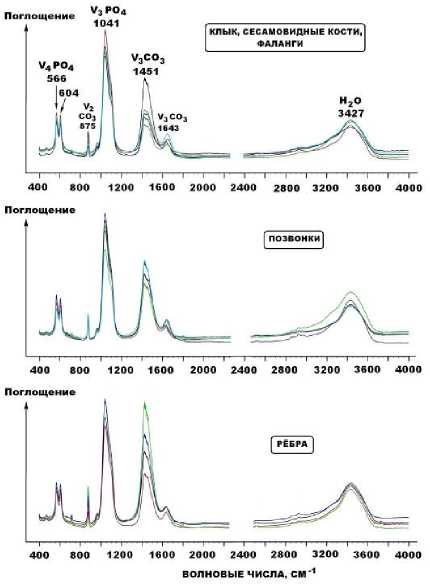
Рис. 17 . ИК спектры костей иманайских пещерных львов
Химический состав биоапатита в исследуемых костях определялся рентгеноспектральным микрозондовым методом. Согласно полученным данным, этот минерал характеризуется сложным и притом разным для львов и медведей составом как в катионной, так и анионной подрешетках, что хорошо отражают соответствующие эмпирические формулы: кости львов – (Ca9.77–10Zn0–0.14Fe0– 0.12)10[(P5.19–5.56C0.44–0.81)6O24](OH)0.83–1.09 Cl0.36– 0.47; кости медведей – (Ca9.88–10Sr0–0.12)10[(P5.14– 5.55S0–0.26C0.45–0.80)6O24] (OH)1.29–1.56Cl0–0.4. Из приведенных формул следует, что иманай-ские пещерные кости в целом характеризуются аномально высокими значениями атомного отношения Ca/P, чем они радикально отличаются от костей внепещерных плейстоценовых животных (рис. 19).
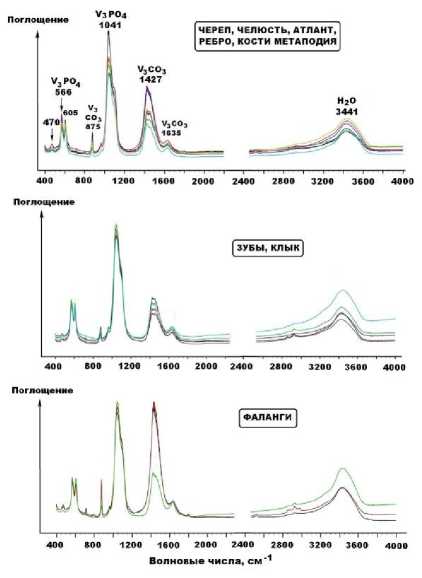
Рис. 18 . ИК спектры костей иманайских пещерных медведей
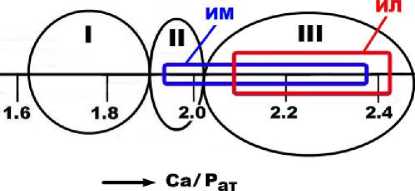
Рис. 19 . Вариации коэффициента атомного отношения Ca/P в первичных и слабоизмененных голоценовых костях внепещерного захоронения (I), в измененных плейстоценовых костях внепе-щерного захоронения (II) и в пещерных кальцинированных костях (III). ИМ и ИЛ – поля значений для костного детрита соответственно медведей и львов из пещеры Иманай
Кроме того, установлено, что биоапатит в костях иманайских львов и медведей системно отличается по структурным примесям: в первых примеси представлены Zn и Fe, а во вторых – Sr и S. Получается, что фоссилизация костей животных происходила не тождественно. Проведенные СЭМ исследования показали, что в биоапатите иманай-ских костей имеется множество микрометровых включений разнообразных ксеногенных минералов. При этом «львиные» и «медвежьи» парагенезисы таких минералов оказались весьма различными. В костях пещерных львов установлены кварц, циркон, рутил состава (Ti 0.98 V 0.02) O 2 ), ильменит (Fe 0.89– 0.99 Mn 0–0.11 Cu 0–0.02 ) 1–1.02 (Ti 0.95–0.98 Al 0–003 V 0.01– 0.02 )O 3 , ре-дкоземельные ортофосфаты
(Ce 0.21–0.47 La 0.1–0.27 Nd 0.06–0.1 Al 0–0.53 Fe 0–0.12 Ca 0– 0.07 Cu 0.03–0.1 Zn 0–0.07 )[PO 4 ] 0.93–0.96 и необычно поликомпонентные шпинелиды (Fe 0.61–
0.94 Mg 0–0.31 Mn 0.02–0.06 Cu 0–0.04 Zn 0–0.05 )(Fe 0.89–
2 Ti 0–0.6 Al 0–0.49 V 0–0.02 ) 2 O 4 ) (миналы, мол. %: магнетит 15–88, ульвит 0–60, герцинит 0–18, магноферрит 0–31, кулсонит 0–1, якобсит 0– 3, галаксит 0–6, ганит 0–3, купрошпинель 0– 4, франклинит 0–5).
В костях пещерных медведей состав мик-роминерального парагенезиса гораздо более сложный: кварц, циркон, ортоклаз, титанит состава Ca 0.99–1.01 (Ti 0.72–0.96 Al 0.04–0.25 Fe 0–0.02 V 0– 0.01 ) 0.99–1 [SiO 4.72–4.96 (OH) 0.04–0.28 ] (миналы, мол. %: титанит 71.65–96.47, вьюаньяит 3.53– 28.35), клиноцоизит (Ca 1.97 Cu 0.06 ) 2.03 (Al 2.26 Fe 0.75 ) 3.01 [Si 3 O 11 ](OH) 1.09 , гранаты (Ca 2.7–3.01 Mg 0–0.37 Mn 0–0.05 ) 2.99–3.03 (Fe 0.06–0.46 Al 1.46–1.88 Ti 0.04–0.08 ) 2 [Si 3 O 12 ] (миналы, мол. %: гроссуляр 65.26–84.03, андрадит 5.93–27.11, пироп 0–12.25, спессартин 0–1.66), шпинелиды (Fe 0.58–1 Mn 0–0.02 Cu 0–0.04 )(Fe 1.65–1.79 Cr 0–0.35 Ti 0–0.15 Al 0–0.06 ) 2 O 4 (миналы, мол. %: магнетит 43.5– 82.5, хромит 0–17.5, ульвит 0–15, герцинит 0–3, якобсит 0–2, купрошпинель 0–4, франклинит 0–36), поликомпонентные сульфато-карбонаты (бариевые (Ba 0.57–0.96 Zn 0–0.36 Cu 0–0.06 Ca 0–0.23 Sr 0–0.04 Fe 0–0.03 )[SO 4 ] 0.56–1.03 [CO 3 ] 0–0.44 , кальциевые (Ca 0.81 Na 0.27 K 0.11 ) 1.19 [SO 4 ], стронциевые (Sr 0.51 Zn 0.43 Cu 0.03 Ca 0.01 K 0.02 )[SO 4 ] 0.44
[CO3]0.55), оксигидроксиды (Mn0.79–0.88Fe0–0.21 Ni0–0.06Cu0–0.03Zn0–0.03Ba0–0.05Ca0–0.07)O(OH)0.86–1 и металлические сплавы (железоникелевые Ni0.87–0.99Fe0.01–0.13S0–0.02, медно-хромово-свин- цовые Pb0.39–0.46Cr0.38–0.39Zn0.08–0.11 Cu0.08–0.12 и свинцово-оловянные Sn0.69Pb0.31).
Таким образом, костный детрит пещерных львов и медведей принципиально различается. В костях львов выявлено 6 минералов и минеральных твердых растворов, относящихся к оксидам, кислородным солям (фосфатам), силикатам, а в костях медведей установлены 13 минералов гораздо более широкого ассортимента: самородные металлы, оксиды, кислородные соли (карбонатсульфаты), силикаты, алюмосиликаты, тита-носиликаты.
Органический матрикс
Исследуемые кости характеризуются аномально низким содержанием С орг – для пещерных львов оно составляет 1.05 ± 0.94, а для медведей 3.57 ± 2.87 мас.% Это примерно на порядок меньше, чем в живых костях, и в 1.5 – 6 раз уступает внепещерным костям плейстоценовых животных. Столь низкое содержание органического углерода в пещерных костях вполне объяснимо. Как известно, при фоссилизации в захороненных костях органический матрикс подвергается не только бактериальному выеданию, но и в еще большей степени окислению и гидролизу (Lindahl, 2000). Понятно, что в условиях пещерных захоронений процесс гидролитического разложения костного коллагена идет особенно быстро вследствие повышенной кислотности внутрипещерных вод.
Физическое присутствие органического вещества в иманайских костях выявлено термическим методом – на кривых нагревания наблюдаются характерные для выгорания органического вещества экзотермические пики. На термограммах львиных костей, как правило, регистрируется единичный экстремум с максимумом при 314– 330°С, а на термограммах костей медведей часто встречаются двухвершинные кривые нагревания с максимумами при 183–332 и 372–415°С. В этом случае экстремумы по своему положению примерно отвечают соответственно «углеводно-липидному» и «аминокислотно-белковому» пикам на кривых нагревания относительно хорошо сохранившегося костного коллагена (Смирнов и др., 2009). Кроме того, на кривых нагревания отчетливо проявляется эндотермический пик с максимумом при 739–785°С, отвечающий диссоциации карбонатов.
Выделение коллагена из исследуемых костей осуществлялось химическим методом деминерализации. Его выход из костных остатков пещерных львов и медведей составил соответственно 0.03–2.9 и 0.5–10.77 мас. %. Полученное органическое вещество варьирует по окраске от светло- до темно-бурого, сохраняя признаки первично-волокнистой текстуры, но львиный коллаген оказался в среднем несколько светлее (рис. 20).
Рис. 20 . Внешний вид коллагена, выделенного из иманайских пещерных костей
По валовому химическому составу (табл. 10) костный коллаген львов и медведей в целом аналогичен, хотя в некоторых деталях не совпадает. Так, львиный костный коллаген в отличие от медвежьего содержит примесь Cu и Zn, но не содержит примеси Ti и Mn. Кроме того, в львином коллагене в два раза меньше железа, в три раза меньше калия и в 6–7 раз меньше хлора.
Элементный состав коллагена, определенный методом газовой хроматографии, у львов и медведей несколько различается, коррелируя с отмеченной выше степенью его сохранности в ископаемых костях (мас. %): львы – С = 35.9–28.4 (32.13 ± 5.3), N = 9.3– 12.5 (10.5 ± 2.26); медведи – С = 40.1–44.3 (42.23 ± 2.1), N = 14.3–15.9 (15.29 ± 0.8).
Тем не менее атомное отношение С/N в коллагене всех образцов оказалось вполне на уровне приемлемости для изотопных исследований (Ambrose, 1990): львы – 3.71 ± 0.16; медведи – 3.49 ± 0.02.
Изотопно-геохимическая информация и палеоэкологические реконструкции
Изотопный состав анализировался во всех образцах последовательно, сначала в костном биоапатите, а затем в выделенном из того же образца коллагене. Полученные результаты (табл. 11) сопоставлялись с аналогичными данными по костным остаткам, захороненным как в пещерных, так и внепе-щерных обстановках на территории Западной Европы, Европейской части России и Северной Евразии.
По изотопному составу углерода и кисло- рода в биоапатите костный детрит иманай-ских пещерных львов и медведей в сравнении с внепещерными костными остатками плейстоценовых животных является аномальным, отличаясь сильным изотопным утяжелением по обоим элементам (рис. 21).
При этом кости иманайских животных характеризуются изотопной неоднородностью в ряду остеологических разностей. Так, в ряду львиных костей относительное утяжеление изотопного состава углерода и кислорода происходит в последовательности: ребро ↔ позвонки → сесамовидные кости. Утяжеление изотопного состава углерода и кислорода в биоапатите медведей усиливается в последовательности: (челюсть, зубы, фаланги) → (череп, атлант) → (кости метаподия, ребро) → бедро.
Проведенный сравнительный анализ показал, что иманайские кости по изотопному составу биоапатита, хотя и являются особенными, но при этом по степени изотопного утяжеления не лидируют среди пещерных ориктоценозов.
Например, кости пещерного медведя из Печорского Приуралья и костные остатки западно-европейских пещерных львов и медведей характеризуются гораздо большим изотопным утяжелением как по углероду, так и кислороду.
Таблица 10 . Валовый химический состав коллагена, выделенного из костей иманайских пещерных львов (1–4) и медведей (5–16), мас. %
|
№п/п |
№ обр. |
SiO 2 |
TiO 2 |
Al 2 O 3 |
Fe 2 O 3 |
MnO |
CuO |
ZnO |
CaO |
BaO |
K 2 O |
P 2 O 5 |
SO 3 |
Cl |
Сумма |
|
1 |
492 |
6.62 |
Н. о. |
0.78 |
0.75 |
Н. о. |
0.83 |
0.89 |
0.18 |
Н. о. |
Н. о. |
Н. о. |
1.03 |
Не обн |
11.08 |
|
2 |
« |
6.63 |
« |
1.07 |
0.55 |
« |
0.62 |
0.61 |
0.53 |
« |
« |
« |
1.92 |
0.26 |
12.19 |
|
3 |
2871 |
0.35 |
« |
Н. о. |
Н. о. |
« |
Н. о. |
Н. о. |
Н. о. |
« |
« |
« |
« |
Н. о. |
0.35 |
|
4 |
2879 |
3.59 |
« |
0.76 |
« |
« |
« |
« |
« |
« |
0.31 |
« |
« |
0.17 |
4.83 |
|
Среднее |
4.3 |
0 |
0.65 |
0.33 |
0 |
0.36 |
0.38 |
0.18 |
0 |
0.08 |
0 |
0.74 |
0.11 |
7.11 |
|
|
СКО |
2.99 |
0.46 |
0.38 |
0.43 |
0.45 |
0.25 |
0.15 |
0.93 |
0.13 |
5.55 |
|||||
|
5 |
1/2 |
0.36 |
« |
Н. о. |
0.75 |
« |
« |
« |
« |
« |
Н. о. |
« |
« |
Н. о. |
1.11 |
|
6 |
« |
3.76 |
« |
0.74 |
0.78 |
« |
« |
« |
« |
« |
0.36 |
« |
0.67 |
6.31 |
|
|
7 |
2 |
Н. о. |
0.33 |
Н. о. |
Н. о. |
« |
« |
« |
« |
« |
0.55 |
« |
1.29 |
0.35 |
2.52 |
|
8 |
« |
0.49 |
Н. о. |
« |
« |
« |
« |
« |
« |
« |
Н. о. |
« |
0.4 |
0.84 |
1.73 |
|
9 |
« |
0.94 |
« |
« |
0.52 |
« |
« |
« |
« |
« |
« |
« |
Н. о. |
0.94 |
2.4 |
|
10 |
(10+11)/1 |
2.38 |
« |
0.5 |
Н. о. |
« |
« |
« |
0.71 |
« |
1.08 |
0.69 |
1.43 |
0.18 |
6.97 |
|
11 |
« |
2.68 |
« |
0.47 |
0.34 |
« |
« |
« |
0.64 |
« |
Н. о. |
0.46 |
1.41 |
0.15 |
6.15 |
|
12 |
« |
3.08 |
« |
0.58 |
Н. о.. |
« |
« |
« |
0.54 |
« |
« |
0.67 |
1.91 |
0.18 |
6.96 |
|
13 |
« |
0.31 |
« |
Н. о. |
« |
« |
« |
« |
Н. о. |
« |
« |
Н. о. |
0.79 |
1.01 |
2.11 |
|
14 |
« |
9.46 |
« |
0.99 |
1.95 |
0.41 |
« |
« |
« |
« |
0.5 |
« |
Н. о. |
0.77 |
14.08 |
|
15 |
« |
6.59 |
« |
1.37 |
1.19 |
Н. о. |
« |
« |
0.96 |
« |
Н. о. |
0.53 |
0.96 |
2.77 |
14.37 |
|
16 |
« |
10.75 |
« |
2.2 |
2.17 |
« |
« |
« |
0.46 |
« |
0.61 |
Н. о. |
0.46 |
1.18 |
17.83 |
|
Среднее |
3.4 |
0.03 |
0.57 |
0.64 |
0.03 |
0 |
0 |
0.28 |
0 |
0.26 |
0.2 |
0.78 |
0.7 |
6.88 |
|
|
СКО |
3.66 |
0.09 |
0.68 |
0.77 |
0.12 |
0.36 |
0.36 |
0.29 |
0.64 |
0.78 |
5.64 |
||||
Таблица 11 . Изотопные характеристики костного детрита иманайских пещерных львов (1–13) и медведей (14–37), ‰
|
№ п/п |
№ обр. |
Остеология |
Биоапатит |
Коллаген |
||
|
δ13С PDB |
δ18O SMOW |
δ13С PDB |
δ15N Air |
|||
|
1 |
492 |
Позвонок |
–7.11 |
21.25 |
–24.51 |
9.1 |
|
2 |
622 |
« |
–8.86 |
20.65 |
–20.7 |
6.1 |
|
3 |
823 |
« |
–7.98 |
21.14 |
–20.0 |
7.5 |
|
4 |
2847 |
« |
–7.69 |
22.62 |
–22.42 |
6.8 |
|
5 |
2869 |
« |
–6.96 |
21.62 |
–24.09 |
8.3 |
|
6 |
2875 |
« |
–7.52 |
19.74 |
–25.6 |
7.7 |
|
7 |
2411 |
Ребро |
–8.36 |
21.43 |
–19.9 |
7.1 |
|
8 |
2803 |
« |
–7.51 |
20.08 |
–19.5 |
8.2 |
|
9 |
2871 |
« |
–8.24 |
21.46 |
–25.73 |
8.1 |
|
10 |
2987 |
Сесамовидная кость |
–6.95 |
19.84 |
–22.7 |
6.6 |
|
11 |
3111 |
« |
–6.14 |
21.06 |
–19.9 |
7.1 |
|
12 |
3046 |
« |
–8.87 |
19.57 |
–22.36 |
9.6 |
|
13 |
2974 |
Фаланга |
–8.11 |
20.68 |
–20.8 |
10.1 |
|
Статистика |
–7.8 ± 0.82 |
20.77 ± 0.91 |
–22.12 ± 2.28 |
7.87 ± 1.19 |
||
|
14 |
7 |
Премоляр |
–6.66 |
21.77 |
Не опр. |
|
|
15 |
(10+11)/2 |
Зуб |
–8.65 |
20.6 |
||
|
16 |
11 |
Премоляр |
–7.12 |
21.35 |
||
|
17 |
4/1a |
Дентин |
–9.41 |
19.94 |
–19.6 1 4.5 |
|
|
18 |
4/1б |
Зубная эмаль |
–9.78 |
20.32 |
Не опр. |
|
|
19 |
4/2 |
Клык |
–9.48 |
17.98 |
–19.6 1 3.6 |
|
|
20 |
9 |
« |
–9.3 |
16.88 |
Не опр. |
|
|
21 |
8 |
Бедренная кость |
–5.07 |
22.02 |
–20.8 |
4.3 |
|
22 |
9 |
« |
–3.38 |
28.01 |
–20.2 |
5.6 |
|
23 |
8+9 |
« |
–8.64 |
21.41 |
–28.72 |
2.9 |
|
24 |
2 |
Кость метаподия |
–6.64 |
21.77 |
–25.61 |
7.2 |
|
25 |
2 |
« |
–6.64 |
21.78 |
–22.68 |
8.12 |
|
26 |
5 |
« |
0.02 |
32.41 |
–21.1 |
4.8 |
|
27 |
12м |
« |
–8.65 |
20.6 |
–24.92 |
6.6 |
|
28 |
12м |
« |
–8.95 |
19.97 |
–20.4 |
4.9 |
|
29 |
1/1 |
Фаланга |
–10.84 |
19.07 |
–23.66 |
5.7 |
|
30 |
10 |
« |
–8.67 |
22.43 |
–23.3 |
3.3 |
|
31 |
10 |
« |
–7.67 |
21.01 |
–22.8 |
3.9 |
|
32 |
12ф |
« |
–8.95 |
19.97 |
–20.4 |
4.9 |
|
33 |
12ф |
« |
–8.67 |
22.43 |
–23.1 |
5.9 |
|
34 |
1/2 |
Череп |
–7.36 |
20.07 |
–23.13 |
5.9 |
|
35 |
(10+11)/1 |
Челюсть |
–9.34 |
19.89 |
–24.92 |
5.2 |
|
36 |
3/1 |
Атлант |
–7.97 |
20.73 |
–19.3 |
3.2 |
|
37 |
3/2 |
Ребро |
–6.17 |
22.39 |
–20.84 |
6.1 |
|
Статистика |
–7.62 ± 2.36 |
21.5 ± 3.01 |
–22.51 ± 2.41 |
5.14 ± 1.42 |
||
Таким образом, в ходе проведенных исследований установлен факт системного изотопного утяжеления углерода и кислорода в биоапатите пещерного костного детрита в сравнении с биоапатитом во внепещерных костных остатках. Очевидно, что такая изотопно-геохимическая особенность пещерных костей обусловлена их кальцитизацией под воздействием углекислотных вод, образующихся за счет карстующихся морских карбонатных пород. Именно возрастанию степени эпигенетической кальцитизации костей за счет вещества осадочных карбонатных пород и соответствует показанный стрелкой генеральный тренд согласованного изотопного утяжеления углерода и кислорода в пещерных костях на рис. 21. Схематическое уравнение такого диспропорционирования имеет следующий вид: 2H12,13CO3– + Ca2+ = Ca13CO3 ↓ + H2O + 12CO2↑ + H2O.
Изотопный состав коллагена в костях иманайских пещерных львов и медведей рассмотрен на фоне аналогичных данных по костному коллагену, волосам и коже плей- стоценовых и современных млекопитающих. На соответствующей диаграмме (рис. 22) поля данных по иманайским животным занимают вполне закономерные места. Поле данных по иманайским львам совмещается с полями хищников, а поле для иманайских медведей тяготеет к полям преимущественно растительноядных животных, в частности современных бурых медведей.
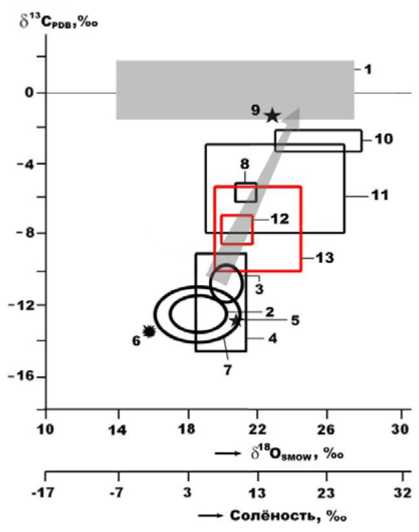
Рис. 21 . Изотопный состав биоапатита в ископаемых костях плейстоценовых млекопитающих на фоне морских карбонатных пород: 1 – карбо-натолиты; 2 – мамонтовая фауна Западной Европы (Tutken et al., 2007); 3 – мамонтовая фауна Печорского Приуралья (Silaev et al., 2017); 4, 5 – соответственно мамонтовая фауна и неоплей-стоценовый бурый медведь Омского Прииртышья (коллекция А. А. Бондарева); 6 – пещерная гиена с Южного Прибайкалья (обр. от Д. В. Кобылкина); 7 – предголоценовые таймырские мамонты (обр. от Г. В. Шнейдера и Д. Н. Костина); 8, 9 – кости пещерного медведя с Печорского Приуралья с разной степенью кальцитизации; 10, 11 – соответственно пещерные львы и медведи Западной Европы (Tutken et al., 2007); 12, 13 – костный детрит соответственно львов и медведей из пещеры Иманай. Шкала солености воды построена по данным, вычисленным по уравнению В. Н. Кулешова (Кулешов, Гаврилов, 2001). Стрелкой показан генеральный тренд изотопного утяжеления углерода и кислорода при переходе от ископаемых костей внепещер-ного захоронения к пещерным костным остаткам
При этом полученные по коллагену има-найских костей изотопные данные свидетельствуют об обитании и львов, и медведей в условиях ландшафтов, переходных от лесов к лесостепям с преобладанием растений с С 3 -типом фотосинтеза7 (большинство растений в зоне прохладного и умеренно влажного климата). Все это вполне соответствует геологическим палеоландшафтным реконструкциям на территорию Южного Урала в конце неоплейстоцена.
Особое значение имеет сравнение полученных нами «коллагеновых» изотопных данных по иманайским костным остаткам с данными Г. Бошрена с соавторами для до-LGM (50—25 тыс. лет назад) и пост-LGM пещерных львов и медведей Западной Европы (Bocherens et al., 2011). На рис. 22 эти данные показаны после проведенной нами их статистической обработки. В сравнении с результатами по иманайской пещере все поля изотопного состава костного коллагена западно-европейских пещерных львов и медведей сдвинуты в область более изотопнотяжелого углерода, что отвечает переходу от лесостепных и лугово-степных ландшафтов преимущественно с растениями С 3 -типа к саваннам с растениями САМ-типа8. При этом обнаруживается, что качество диеты до-LGM популяции западно-европейских пещерных львов вполне сопоставимо с качеством питания иманайских пещерных львов, а вот качество диеты пост-LGM популяции пещерных львов в Западной Европе резко снизилось. Последнее выглядит вполне объяснимо, если учесть постледниковое вымирание мегафауны, служившей пищевой базой для крупных хищников в течение почти всего плейстоцена.
Значительный интерес представляет изотопная реконструкция пищевой цепи пещерных львов. Как известно, в основе таких реконструкций лежат уже хорошо обоснованные сдвиги в изотопном составе углерода и азота при переходе по трофическим цепям от растительноядных животных к хищникам (De Niro, 1985; Bocherens, Drucker, 2003).
Согласно выводу Г. Бошрена, основной пищевой жертвой до-LGM популяции западноевропейских пещерных львов был молодняк пещерных медведей. Проведенные нами расчеты на основании данных Г. Бошрена вполне подтвердили этот вывод (правая стрелка на рис. 22). Аналогичные расчеты с использованием изотопных данных по има-найским пещерным львам показывают, что их пищевой жертвой являлись не пещерные медведи, а с достаточно большой вероятностью шерстистые носороги (левая стрелка на рис. 22). Такой вывод не является особенно оригинальным для неоплейстоцена. Молодь шерстистых носорогов и даже шерстистых мамонтов достаточно часто рассматривается в качестве типичной пищевой жертвы пещерных львов, в частности на территориях Аляски и Северной Евразии (Guthrie, 1968; Turner, 2009; Stuart, Lister, 2011). Хотя, конечно, в случае с иманайскими львами нельзя исключать и таких пищевых жертв, как северный олень и лошади.
В целом, судя по изотопному составу коллагенового углерода и азота, неоплейстоце-новые пещерные медведи как в Западной Европе, так и на Южном Урале, были растительноядными животными и вполне могли оказаться (по версии Г. Бошрена) пищей для хищных пещерных львов, если бы, конечно, пересекались с ними по жизни. Однако при этом львы вовсе не были «ужасными хищниками» ни во времена своего существования, ни в проекции на современность. Например, для неоплейстоцена Западной Европы трофическая цепь реконструирована в следующем виде: (растительноядные животные, включая пещерного медведя) → бурый медведь → рысь → лисица → пещерный лев → волк → росомаха → пещерная гиена (Bocherens et al., 2011; Bocherens, 2019).
В рамках этой реконструкции пещерный лев находится лишь в средине пищевой цепи, чему соответствуют конкретные изотопные данные как по Западной Европе, так и Евразии (рис. 22). Например, пещерная гиена из Южного Прибайкалья, судя по ее исследованному нами костному коллагену, была более продвинутым хищником, чем евразийские пещерные львы, практически ничем в этом смысле не отличаясь от западноевропейских пещерных гиен.
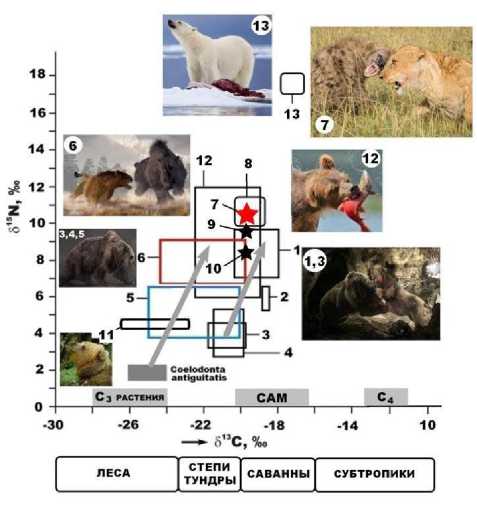
Рис. 22 . Изотопный состав костного коллагена, волос и кожи плейстоценовых и современных млекопитающих: 1, 3 и 2, 4 – соответственно до-LGM и пост-LGM популяции пещерных львов и медведей Западной Европы (Bocherens et al., 2011); 5, 6 – соответственно львы и медведи из пещеры Иманай; 7 – пещерная гиена с Южного Прибайкалья (обр. от Д. В Кобылкина); 8–10 – соответственно пещерная гиена, ископаемые волк и росомаха Западной Европы (Bocherens, 2019); 11 – современные таежные бурые медведи (волосы и кожа); 12 – современные камчатские бурые медведи (волосы и кожа); 13 – современные белые медведи (волосы и кожа). Шкала ландшафтов показана по Г. Бошрену и Д. Дракер (Bocherens, Drucker, 2003)
Как показали проведенные нами исследования, пещерные львы по изотопной диете статистически уступали даже некоторым современным бурым медведям, а именно камчатским, не говоря уже о белом медведе, вероятно, величайшем хищнике кайнозоя. Правда, следует учесть, что значительную долю пищи камчатского бурого медведя составляет тихоокеанская рыба, а рацион питания белого медведя и вовсе состоит из океанской рыбы и зверя, что весьма значительно утяжеляет изотопный состав органического азота. Потреблением рыбы, кстати, можно объяснить и недавно выявленные факты изотопного утяжеления азота в костном коллагене некоторых плейстоценовых пещерных медведей в Западной Европе (Bocherens, Drucker, 2019).
Присутствие древних людей в пещере Иманай
В ходе раскопок в пещере Иманай были обнаружены многочисленные артефакты, диагностированные археологами как каменная индустрия мустьерской эпохи (среднего палеолита), приписываемая поздним неандертальцам. Кроме того, в черном материале из базального горизонта № 12 нами были обнаружены частицы, очень похожие на частицы древесного угля, который может иметь отношение к древним кострищам. Как известно, в настоящее время в качестве признаков таких кострищ археологами рассматриваются «красные глины», «обгоревшие камни или породы», «обугленные кости», «кремни со следами обработки огнем», «зольные пятна», «древесный уголь» (James, 1989; Амирханов и др., 2013). Из этого списка наиболее приемлемым признаком древних кострищ являются, разумеется, частицы древесного угля, но их диагностика требует использования специальной аналитики.
В нашем случае в качестве такой аналитики был выбран метод КР спектроскопии, которым обычно и диагностируют углеродистые вещества (Силаев и др., 2013). Полученные от черных частиц из Иманайской пещеры и частиц современного древесного угля, образовавшегося в результате сгорания древесины на одном из действующих вулканов Курильской островной гряды, спектры были отфильтрованы от сопутствующей люминесценции. После этого в спектрах обнаружились две рамановские полосы – D (1340 и 1374 см-1) и G (1594 и 1606 см-1) – с харак-

Рис. 23 . Орудия культуры мустье (средний палеолит), Иманай (а) и частицы древесного угля из пещеры Иманай (б) и с современного вулкана Курильской островной гряды (в)
теристиками, отвечающими неметаморфизо-ванному углистому веществу растительного происхождения (рис. 23). Таким образом, обнаруженные в базальном горизонте элювиальных грунтов в иманайской пещере углеродистые частицы оказались именно частицами древесного угля, указывая тем самым на древние кострища и подтверждая факт обитания в пещере Иманай неоплей-стоценовых людей. Уже выдвинуты несколько версий использования пещеры Има-най неандертальцами (Паршукова, 2016): в качестве жилища, святилища для отправления культов, ловушки для промысловых животных (например, пещерного медведя), «холодильника» для сохранения охотничьих припасов. В принципе способность палеоантропов к такому социально сложному поведению уже давно не вызывает сомнений (Вишняцкий, 2010; Формозов, 2019), но для конкретного решения этого вопроса в отношении иманайской пещеры необходимо продолжение раскопок с нацеленностью на поиски антропогенных костных останков и последующее их мультидисциплинарное исследование.
Факт обнаружения признаков обитания людей в пещере Иманай далеко (400–500 км) за пределами ареала распространения поздних неандертальцев на территории Западной Европы и Южной Евразии (рис. 24) является сильным подтверждением теории начавшейся примерно 50–40 тыс. лет назад необратимой миграции неандертальцев на север вследствие, прежде всего, изменения климата. В ходе такой миграции Южный Урал вовсе не являлся крайним рубежом. На это ука- зывает комплекс данных, полученных П.Ю. Павловым при раскопках стоянки начала верхнего палеолита Заозерье (Силаев и др., 2019). Более того, обнаружение П.Ю. Павловым на расположенной гораздо севернее стоянке Бызовой многочисленных артефактов, связанных с жизнью и деятельностью палеоантропов (Slimak et al., 2011), показывает, что уже к середине каргинского интерстадиала неандертальцы практически достигли Полярного круга, а возможно, и выходили непосредственно на побережье Арктического океана.
Справедливости ради следует отметить, что каргинский интерстадиал был временем больших миграций на север не только неандертальцев, но и людей современного типа. На последнее, в частности, указывает находка костных останков усть-ишимского человека на территории Омского Прииртышья (Силаев и др., 2017). Местонахождение усть-ишимской кости – на 57° с. ш., т. е. на 1500– 2000 км севернее недавней находки современника усть-ишимского человека в пещере Бачо Киро на территории современной Болгарии (Hublin et al., 2020), что считается более естественным местом обитания людей современного типа в позднем палеолите.
Рис. 24. Некоторые стоянки поздних неандертальцев в Европейской части России на фоне ареала их традиционного распространения по территории Западной Европы и Южной Евразии (показан буроватым цветом). Стрелка – направление миграции неандертальцев
Тем не менее следы пребывания современников усть-ишимского человека установлены и на Таймыре – на 72° с. ш., что, действительно, свидетельствует о широком заселении Арктики людьми современного типа сразу после ранневалдайского оледенения (Pitulko et. al., 2016).
Заключение
Полученные экспериментальные данные аргументируют вывод о том, что костные остатки неоплейстоценовых медведей и львов из пещеры Иманай принципиально отличаются практически по всем минералогогеохимическим свойствам от аналогичных по возрасту костных остатков, фоссилизиро-ванных во внепещерных обстановках.
Иманайский пещерный костный детрит в целом обнаруживает гораздо большую степень вторичных изменений, чем внепещер-ные костные остатки, что наиболее наглядно выражается в коллофанизации биоапатита и в деградации наноструктуры костей. При этом пещерные кости многократно уступают внепещерным по содержанию иллю-виированных в кости минеральных загрязнений .
Типоморфным признаком пещерных костей является их интенсивная эпигенетическая кальцитизация , т. е. замещение первичного биоапатита вторичными карбонатами. Это выражается в 3–5 раз большем содержании СО 2 в костях и в аномально высоких значениях атомного отношения Ca/P, что совершенно не свойственно внепещерному костному детриту. Факт кальцитизации пещерных костей коррелируется с фосфатиза-цией вмещающих кости грунтов, что объясняется выносом в последние замещенного в костях фосфатного вещества.
В составе исследуемых пещерных костей обнаружены 50 микроэлементов, в том числе элементы-эссенциалы (доля в балансе элементов 25–45 %), физиологически-активные элементы (60–72 %) и элементы-антибионты (0–5 %). Валовая концентрация микроэлементов в иманайских костях в 2–3 раза уступает концентрации микроэлементов во вне-пещерных костях. Отношение суммарных содержаний элементов-эссенциалов и анти-бионтов колеблется в пределах 17–158, что гораздо ближе к живым организмам, чем к ископаемым костям, фоссилизированным во внепещерных условиях. Кроме того, пещерные кости кардинально отличаются от вне- пещерных по всем геохимическим модулям. Все эти геохимические различия обусловлены аномально низким содержанием в пещерных костях микроэлементов, имеющих источник в костеносных грунтах. Таким образом, фоссилизация костей в пещерных условиях принципиально отличается от таковой во внепещерных обстановках крайне низкой степенью эпигенетического обогащения микроэлементами за счет вмещающей геологической среды.
Значения рентгеновского индекса кристалличности биоапатита в костях львов и медведей в среднем близки к аналогичным данным, полученным для внепещерных костей, но в 1.5 раза уступают по зубам и клыкам. ИК спектроскопический индекс Iν 3 [PO 4 ] / Iν 3 [CO 3 ] биоапатита в иманайских костях варьируется в пределах 1.36–3.51, что в среднем на 35–40 % ниже, чем для внепе-щерных костей. Это коррелируется с фактом значительной кальцитизации пещерных костных остатков.
Иманайские пещерные кости характеризуются на порядок меньшим содержанием С орг по сравнению с живыми костями и в 1.5–6 раз меньшим по сравнению с внепещерным костным детритом . Выход коллагена из исследованных костных образцов варьируется от 0.5 до 11 вес. %. По составу примесей он различается: львиный костный коллаген в отличие от медвежьего содержит примесь Cu и Zn, но не содержит примеси Ti и Mn. Кроме того, в львином коллагене в два раза меньше железа, в три раза меньше калия и в 6–7 раз меньше хлора. По атомному отношению С/N (3.47–3.87) полученный из костей коллаген находится на уровне, вполне приемлемом для изотопных исследований.
По углероду и кислороду в биоапатите костный детрит иманайских пещерных львов и медведей является изотопно аномально тяжелым, сближаясь с изотопным составом углерода и кислорода в морских известняках. Как показали наши и зарубежные исследования, такая изотопия биоапатита является типоморфным признаком пещерных ориктоценозов, будучи обусловленной эпигенетической кальцитизацией под воздействием углекислотных вод, образующихся в карстующихся карбонатных породах.
Изотопные данные, полученные по коллагену, свидетельствуют об обитании иманайских пещерных львов и медведей в условиях ландшафтов, переходных от лесов к лесостепям с преобладанием растений С 3 по типу фотосинтеза (большинство растений в зоне прохладного и умеренно влажного климата). Результат расчета трофической цепи с использованием изотопных данных по коллагеновому углероду и азоту позволяет предположить, что вероятной пищевой жертвой иманайских пещерных львов в отличие от западно-европейских были не пещерные медведи, а шерстистые носороги.
Сравнительный анализ полученных данных показывает, что при всей общей специфичности костные остатки иманай-ских львов и медведей по многим свойствам не совпадают. Они статистически расходятся по размерам: обломки скелета львов в среднем почти в три раза крупнее, чем обломки скелета медведей. Львиные и медвежьи кости существенно различаются по химическому и нормативноминеральному составу иллювиированных минеральных загрязнений, а также по валовому содержанию микроэлементов. Эти кости демонстрируют несколько разную степень измененности физических свойств: в львиных костях биоапатит сильнее коллофа-низирован, а медвежьи кости отличаются большей степенью деградации наноструктуры. Кости львов карбонизированы на 15–20 % значительнее, чем кости медведей, и различаются по парагенезисам микроминералов-примесей: в львиных костях установлены оксиды, кислородные соли (фосфаты) и силикаты, а в костях медведей выявлена гораздо более широкая ассоциация микроминералов – самородные металлы и сплавы, оксиды, кислородные соли (карбонатсульфаты), силикаты, алюмосиликаты, тита-носиликаты. Собственно биоапатит в има-найских костях кристаллохимически тоже нетождественен как в катионной, так и анионной подрешетках. Для львиных костей характерны примеси Zn и Fe, а для медвежьих – Sr и S.
Проведенные исследования выявили и некоторые анатомические различия фоссили-зации пещерных костей. Так, в направлении от зубов к костям скелета снижается индекс кристалличности биоапатита, но увеличиваются интенсивность кальцити-зации, степень деградация наноструктуры и валовая концентрация микроэлементов.
Совместное нахождение разновозрастного и в разной степени фоссилизированного костного детрита в элювиальных грунтах пещеры Иманай представляет собой очевидный спелеопалеонтологический парадокс . Объяснение этого парадокса может состоять в более молодом, чем кости, возрасте костеносных грунтов, в которых костные остатки переотлагались с перемешиванием. Исходя из климатической истории конца плейстоцена–начала голоцена можно предположить, что упомянутые грунты, будучи результатом растворения карстующихся известняков, образовались не раньше 15–10 тыс. лет назад. Если принять такую гипотезу, то следует допустить и то, что до захоронения в элювиальных грунтах костные остатки львов и медведей в течение соответственно 32 – 28 и 25 – 20 тыс. лет назад подвергались фос-силизации в особых условиях внутрипе-щерной атмосферы и капельной углекислотной гидросферы. Время такой фосси-лизации можно определить как этап спелеовыветривания, который позже сменился этапом фоссилизации в грунтах (рис. 25).
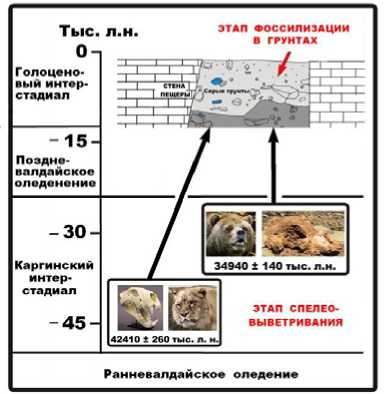
Рис. 25. Схема хронологии и этапности пещерной фоссилизации костных остатков плейстоценовых млекопитающих на примере пещеры Иманай
Именно разная продолжительность (на 7– 8 тыс. лет) этапа спелеовыветривания костных остатков львов и медведей и послужила, вероятно, причиной существенного и системного расхождения их минералогогеохимических свойств.
tбнаруженные в пещерно-элювиальных грунтах каменные артефакты среднего палеолита и частицы древесного угля в базальном горизонте осадочного профиля свидетельствуют об обитании в пещере Иманай поздних неандертальцев далеко за пределами традиционного ареала их распространения на территории Западной Европы и Южной Евразии. Это отражает начавшуюся примерно 50–40 тыс. лет назад миграцию неандертальцев на север, в ходе которой они к концу среднего валдая достигли, по крайней мере в европейской части современной России, Полярного круга.
За аналитическое обеспечение авторы благодарят ведущего инженера, к. г.-м. н. Е.А. Васильева (Санкт-Петербургский горный университет), руководителя лабораторией химии минерального сырья О.В. Кокшарову (ИГ Коми НЦ УрО РАН), с. н. с., к. г.-м. н. Ю.С. Симакову и Ю.В. Глухова (ИГ Коми НЦ УрО РАН), старшего физика М.Ф. Самотолкову, химиков-аналитиков С.Т. Неверова (ИГ Коми НЦ УрО РАН) и Е.А Туманову (ИБ Коми НЦ УрО РАН). Выражают также признательность д. и. н. П.Ю. Павлову (ИЯЛИ Коми НЦ УрО РАН) и к. г.- м. н. Д.В. Пономареву (ИГ Коми НЦ УрО РАН) за ценное научное консультирование.
Список литературы Минералого-геохимические особенности пещерной фоссилизации ископаемых костей на примере пещеры Иманай (Южный Урал)
- Амирханов Х.А., Бронникова М.А., Таймазов А.И. О следах огня на стоянке олдована Айникаб 1 в Центральном Дагестане // Древнейший Кавказ: перекресток Европы и Азии / ИИМК РАН. СПб., 2013. С. 7-19.
- Вишняцкий Л. Неандертальцы: история несостоявшегося человечества. СПб.: Нестор-Исто-рия, 2010. 312 с.
- Войнар А. О. Биологическая роль микроэлементов в организме животных и человека. М.: Высшая школа, 1960. 541 с.
- Вотяков С.Л., Киселёва Д.В., Щапова Ю.В., Смирнов Н.Г., Садыкова Н.О. Физико химические характеристики ископаемых костных остатков млекопитающих и проблема их относительного возраста. Ч. 2. ИК и радиоспектроскопия. Екатеринбург: Изд-во "Гощицкий", 2009. 82 с.
- Гимранов Д.О., Косинцев П.А., Нурмухаметов И.М., Некрасов А.Е. Первая находка дикообраза (Hystrix sp.) и кабана (Sus scrofa) в позднем плейстоцене Южного Урала // Фундаментальные проблемы квартера: Итоги изучения и основные направления исследований: матер. X Всерос. совещания по изучению четвертичного периода. М.: Геос, 2017. С. 89.
