Многолетняя динамика обилия фоновых видов птиц Баргузинского хребта в гнездовой период

Автор: Ананин Александр Афанасьевич
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Биология
Статья в выпуске: 4, 2011 года.
Бесплатный доступ
В статье представлен анализ долговременных изменений обилия гнездящихся птиц в 1984-2007 гг.
Гнездящиеся птицы, динамика обилия, баргузинский хребет
Короткий адрес: https://sciup.org/148180284
IDR: 148180284 | УДК: 598.2
Long-term dynamics of abundance of background species birds of the Barguzinsky mountain ridge during the nesting period
In 1984-2007 the analysis of long-term changes of abundance of 55 species of nesting birds was conducted.
Текст научной статьи Многолетняя динамика обилия фоновых видов птиц Баргузинского хребта в гнездовой период
Изучение долговременных изменений численности популяций животных в природных условиях приобретает особую значимость в связи с тенденциями роста влияния антропогенных факторов. Длительные стационарные наблюдения за численностью вида позволяют выявить особенности его реагирования на изменения условий среды обитания, которые складываются в конкретный год. Удобным объектом для такого рода исследований во времени и пространстве являются птицы.
При изучении изменений численности в одном месте, на сравнительно ограниченной территории, чаще наблюдаются локальные изменения. О многолетней динамике большинства видов нельзя судить по результатам учетов на каком-либо одном участке, так как в разных местах ее изменения происходят неодинаково. Птицы способны ежегодно перераспределяться по территории в зависимости от хода весны и условий обитания, сложившихся в разных местах ко времени их прилета и началу гнездования (Данилов, 1966, 1980; Данилов и др., 1984).
В задачи работы входило выявление тенденций долговременных изменений обилия гнездящихся птиц на модельной территории Баргузин-ского хребта, в том числе для групп дальних и ближних мигрантов и оседлых видов.
Материал и методы
Ключевой участок для долговременного орнитологического мониторинга размещается на территории государственного природного биосферного заповедника «Баргузинский». Дина- мика численности летнего населения птиц прослежена в 1984-2007 гг. на постоянных учетных маршрутах, размещенных в долинах трех рек от побережья оз. Байкал до высокогорий Баргузин-ского хребта (460-1700 м над ур.м). На этих высотных профилях, разбитых на 11 участков, представлена большая часть разнообразия местообитаний нижней и верхней части горнолесного и подгольцово-субальпийского поясов. Здесь выделены прибрежно-равнинный (байкальские террасы), предгорный (нижняя часть горно-лесного пояса), горно-лесной (верхняя часть горно-лесного пояса) и подгольцовосубальпийский выделы (Ананин, 2006а-б, 2009а-б, 2010а-б).
Общая протяженность летних пеших маршрутных учетов, положенных в основу настоящей работы, – 5552 км. Обилие птиц рассчитано по методу Ю.С. Равкина (1967). Видовая классификация птиц принята по Л.С. Степаняну (2003). Статистические расчеты выполнены с применением пакета программ Statistica 6.0 с учетом методических рекомендаций, предложенных для анализа временных рядов данных (Коросов, 2007).
Результаты
Гнездовая численность каждого вида птиц на ключевом участке Баргузинского хребта за 24летний период исследований испытывала колебания различной амплитуды и длительности. Для большинства регулярно гнездящихся видов установлены кратковременные непрерывные флуктуации обилия с амплитудой, как правило, не превышающей 2-8-кратных значений.
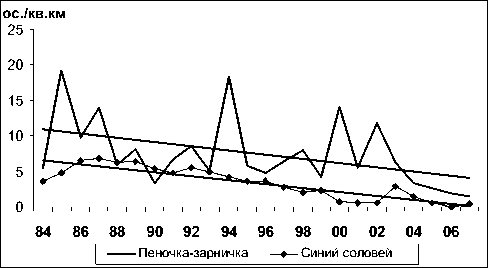
Высокая изменчивость среди фоновых видов ключевого участка выявлена для таких видов, как белокрылый клест ( Loxia leucoptera ), пеноч-ка-зарничка ( Phylloscopus inornatus ), синехвостка ( Tarsiger cyanurus ) и длиннохвостая синица ( Aegithalos caudatus ) (13-19-кратные изменения), обыкновенная пищуха ( Certhia familiaris ) и пятнистый сверчок ( Locustella lanceolata ) (27-37-кратные), а наибольшая – для вьюрка ( Fringilla montifringilla ) (52-кратные), склонного к значительным ежегодным перемещениям в пределах гнездового ареала. Большой размах изменчивости характерен также для синего соловья ( Lus-cinia cyane ), дубровника ( Emberiza aureola ), сибирской мухоловки ( Muscicapa sibirica ), для которых уровень численности в последнем десятилетии существенно снизился и в отдельные годы они почти отсутствовали на учетных маршрутах. Асинхронный характер колебаний обилия у разных видов способствует стабилизации плотности населения в целом.
Из 55 включенных в анализ фоновых видов, регулярно встречающихся на ключевом участке, статистически достоверная тенденция увеличения обилия (положительный тренд, рассчитанный как коэффициент ранговой корреляции многолетних изменений плотности гнездования с временным рядом) (рис. 1) обнаружена у 8 видов: чижа (Spinus spinus), рябчика (Tetrastes bonasia), бурой пеночки (Phylloscopus fuscatus), белокрылого клеста, желтоголового королька (Regulus regulus), певчего сверчка (Locustella certhiola), пестрого дрозда (Zoothera dauma) и черныша (Tringa ochropus) (табл. 1). Статистически значимая тенденция сокращения обилия (рис. 2) зафиксирована для 16 видов: буроголовой гаички (Parus montanus), пеночки-зарнички, горной трясогузки (Motacilla cinerea), обыкновенной (Carpodacus erythrinus) и сибирской (C. roseus) чечевиц, овсянок: рыжей (Emberiza rutila), желтобровой (E. chrysophrys), седоголовой (E. spodocephala), белошапочной (E. leuco-cephala) и дубровника, синего соловья, длиннохвостой синицы, сибирской мухоловки, красно-зобого (Turdus ruficollis) и оливкового (T. obscu-rus) дроздов, а также щура (Pinicola enucleator). Еще у 31 вида статистически существенные тренды не проявились (рис. 3), плотность гнездования была относительно стабильной.
Отрицательные тенденции преобладают у дальних мигрантов (11 видов против 4 с положительными трендами). У оседлых видов, как и у ближних мигрантов, это соотношение равное (3:3 и 1:2 соответственно) (табл. 2).
Максимальная доля видов, снижающих гнездовое обилие, относится к периферийной и погранично-ареальной группам (35,7 и 42,9% соответственно) (табл. 2).
Обсуждение результатов
Анализ долговременных изменений численности видов птиц на ключевом участке, выполненный нами ранее за 1984-1998 гг. (Ананин, 2001) для 50 видов птиц со средней многолетней численностью более 1 ос./км2, показал, что число видов с отрицательными и положительными статистически значимыми трендами в Северо-Восточном Прибайкалье составило 16 и 17, что было очень близко к соотношению, полученному в Средней Сибири в условиях долины р. Енисей в период 1978-1987 гг. – 19 и 17 (Бурский, 1988). В Прибалтике это соотношение было нарушено: 23 и 6 видов (Паевский, 1985). В изменении численности мигрирующих видов Северо-Восточного Прибайкалья в конце ХХ в. преобладали отрицательные тенденции, а у видов, зимующих в Восточной Сибири, – положительные, что соответствовало и наблюдениям О.В. Бурского в долине р. Енисей (Бурский, 1988, 2007).
На ключевом участке в 2000-е гг. проявилась тенденция снижения численности всего населения практически во всех исследованных выде-лах, что явилось следствием превышения доли видов с отрицательными трендами над числом видов с положительным трендом (табл. 1). При этом возросла доля видов с отрицательными тенденциями долговременных изменений численности среди дальних мигрантов, зимующих на всех южно-азиатских зимовках: пакистаноиндийской, китайской, индокитайской и в меньшей степени на филлипино-малакко-индоне-зийской зимовках, что может свидетельствовать о их неблагополучном состоянии. Такой же точки зрения в отношении причин снижения численности хищных птиц региона придерживается В.В. Рябцев (2009, 2010).
Таблица 1
Тренды долговременных изменений гнездового обилия фоновых видов птиц на ключевом участке Баргузинского хребта в 1984-2007 гг .
( уровень значимости : *p<0,05; **p<0,01; ***p<0,001)
|
Вид |
КУ |
ПР |
ПГ |
ГЛ |
ПС |
|
1 |
2 |
3 |
4 |
5 |
6 |
|
Всего : |
— *** |
— *** |
0 |
— *** |
0 |
|
Sitta europaea |
0 |
0 |
0 |
0 |
0 |
|
Parus ater |
0 |
0 |
0 |
— * |
0 |
|
Parus montanus |
— *** |
— *** |
— * |
— *** |
0 |
|
Phylloscopus proregulus |
0 |
0 |
— *** |
0 |
0 |
|
Anthus hodgsoni |
0 |
0 |
0 |
— ** |
0 |
|
Tarsiger cyanurus |
0 |
0 |
0 |
0 |
0 |
|
Phylloscopus inornatus |
— ** |
— * |
0 |
0 |
0 |
|
Spinus spinus |
+** |
+*** |
+* |
0 |
0 |
|
Phylloscopus trochiloides |
0 |
0 |
0 |
— * |
0 |
|
Motacilla cinerea |
— *** |
0 |
— *** |
0 |
— * |
|
Carpodacus erythrinu |
— ** |
— ** |
0 |
0 |
0 |
|
Tetrastes bonasia |
+** |
+* |
0 |
0 |
0 |
|
Emberiza rutila |
— *** |
— *** |
— ** |
||
|
Dendrocopos major |
0 |
0 |
0 |
0 |
|
|
Nucifraga caryocatactes |
0 |
0 |
0 |
0 |
0 |
|
Pyrrhula cineracea |
0 |
0 |
0 |
0 |
0 |
|
Phylloscopus fuscatus |
+** |
+** |
0 |
||
|
Loxia leucoptera |
+** |
+* |
0 |
0 |
0 |
|
Ficedula mugimaki |
0 |
+* |
0 |
||
|
Emberiza chrysophrys |
— ** |
0 |
— * |
0 |
|
|
Fringilla montifringilla |
0 |
0 |
0 |
0 |
0 |
|
Luscinia calliope |
0 |
0 |
0 |
0 |
0 |
|
Luscinia cyane |
— *** |
— *** |
— *** |
— *** |
|
|
Aegithalos caudatus |
— * |
0 |
0 |
— * |
|
|
Perisoreus infaustus |
0 |
0 |
0 |
0 |
0 |
|
Carpodacus roseus |
— *** |
— ** |
0 |
||
|
Actitis hypoleucos |
0 |
0 |
0 |
||
|
Emberiza aureola |
— *** |
— *** |
— * |
||
|
Certhia familiaris |
0 |
0 |
0 |
0 |
|
|
Ficedula parva |
0 |
0 |
0 |
0 |
0 |
|
Emberiza spodocephala |
— ** |
0 |
|||
|
Troglodytes troglodytes |
0 |
0 |
0 |
0 |
|
|
Muscicapa sibirica |
— *** |
— *** |
— *** |
— *** |
|
|
Locustella lanceolata |
0 |
0 |
0 |
||
|
Prunella montanella |
0 |
0 |
0 |
||
|
Regulus regulus |
+* |
+* |
+** |
0 |
|
|
Anthus spinoletta + A. rubescens |
0 |
0 |
|||
|
Cinclus cinclus |
0 |
0 |
0 |
||
|
Cuculus saturatus |
0 |
0 |
0 |
0 |
0 |
|
Turdus ruficollis |
— *** |
— ** |
0 |
||
|
Picoides tridactylus |
0 |
— *** |
0 |
0 |
0 |
|
Streptopelia orientalis |
0 |
0 |
0 |
0 |
|
|
Locustella certhiola |
+*** |
+*** |
|||
|
Emberiza leucocephala |
— ** |
— *** |
|||
|
Turdus philomelos |
0 |
0 |
0 |
0 |
|
Phylloscopus borealis |
0 |
0 |
|||
|
Lanius cristatus |
0 |
— * |
|||
|
Zoothera dauma |
+* |
+** |
0 |
0 |
0 |
|
Dryocopus martius |
0 |
0 |
0 |
0 |
|
|
Tringa ochropus |
+** |
+* |
+* |
— * |
|
|
Motacilla alba |
0 |
0 |
— * |
||
|
Turdus obscurus |
— ** |
0 |
0 |
0 |
|
|
Cuculus canorus |
0 |
0 |
0 |
0 |
0 |
|
Pinicola enucleator |
— ** |
— ** |
— * |
||
|
Bombycilla garrulus |
0 |
0 |
0 |
||
|
Всего видов |
55 |
47 |
43 |
39 |
34 |
|
Тренд положительный |
8/ 14,5 |
9/ 19,1 |
2/ 4,7 |
0 |
0 |
|
Тренд отрицательный |
16/ 29,1 |
10/ 21,3 |
9/ 20,9 |
9/ 23,1 |
4/ 11,8 |
Примечания: — - тренд отрицательный; + - тренд положительный; 0 - тренд не выражен; пропуск данных -вид в данном выделе регулярно не встречается. В характеристике тренда: числитель – число видов, знаменатель – доля от числа видов, регулярно встречающихся в выделе (в %). Наименования выделов: КУ – ключевой участок, ПР – прибрежно-равнинный, ПГ – предгорный, ГЛ – горно-лесной, ПС – подгольцово-субальпийский.
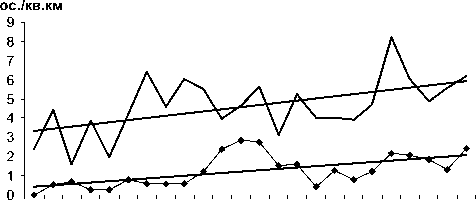
84 86 88 90 92 94 96 98 00 02 04 06
Бурая пеночка
Певчий сверчок ос./кв.км
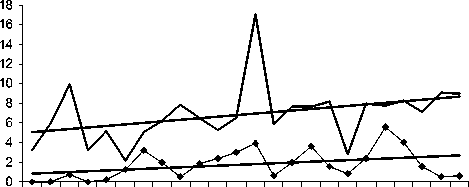
84 86 88 90 92 94 96 98 00 02 04 06
Рябчик
Желтоголовый королек
ос ./ кв . км
4,5
4,0
3,5
3,0
2,5
2,0
1,5
1,0
0,5
0,0
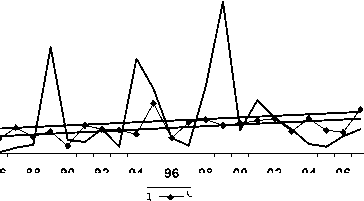
Черныш
84 86 88 90 92 94
98 00 02 04 06
Пестрый дрозд
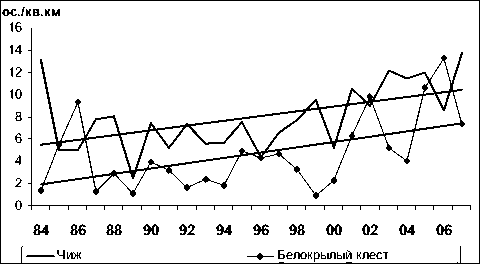
Рис. 1. Изменения гнездового обилия видов птиц ключевого участка Баргузинского хребта с положительным трендом в 1984-2007 гг. (особ./км2)
84 86 88 90 92 94 96 98 00 02 04 06
Горная трясогузка Краснозобый дрозд
84 86 88 90 92 94 96 98 00 02 04 0 6
\ Обыкновенная чечевица ♦ Сибирская мухоловка I
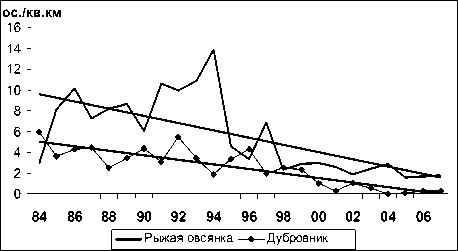
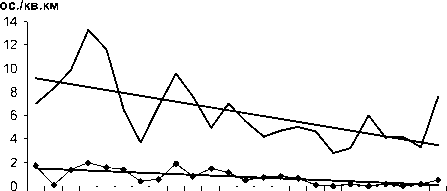
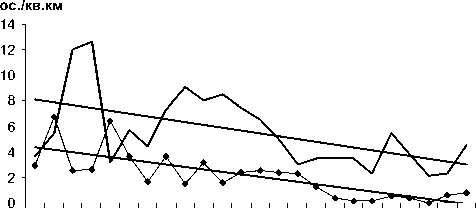
Рис. 2. Изменения гнездового обилия видов птиц ключевого участка Баргузинского хребта с отрицательным трендом в 1984-2007 гг. (особ./км2)
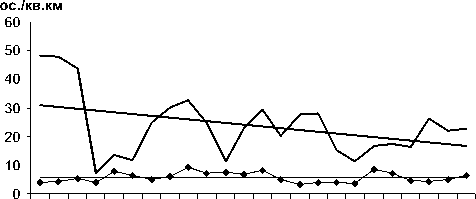
84 86 88 90 92 94 96 98 00 02 04 06
I Обыкновенный поползень ♦ Пестрый дятел

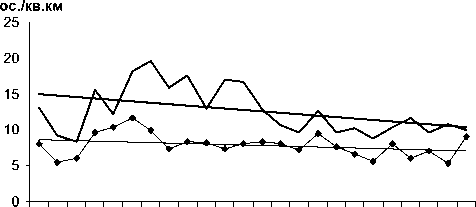
84 86 88 90 92 94 96 98 00 02 04 06
I---- Корольковая пеночка, ♦ Зеленая пеночка
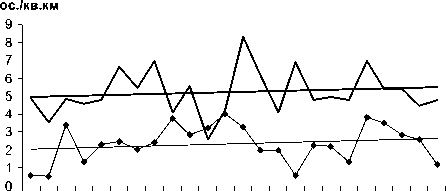
84 86 88 90 92 94 96 98 00 02 04 06
I Серый снегирь —Крапивник
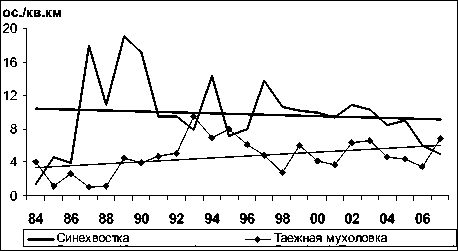
Рис. 3. Изменения гнездового обилия видов птиц ключевого участка Баргузинского хребта со статистически незначимым трендом в 1984-2007 гг. (особ./км2).
Таблица 2
Состав групп видов птиц ключевого участка Баргузинского хребта с различной тенденцией долговременных изменений гнездового обилия в 1984-2007 гг .
( число видов )
|
Тип группировки |
Численность птиц |
|||||
|
возрастает |
снижается |
стабильная |
||||
|
абс. |
% |
абс. |
% |
абс. |
% |
|
|
1.0. Район зимовки |
8 |
14,5 |
16 |
29,1 |
31 |
56,4 |
|
1.1. Оседлые |
3 |
20,0 |
3 |
20,0 |
9 |
60,0 |
|
1.2. Ближние мигранты |
1 |
14,3 |
2 |
28,6 |
4 |
57,1 |
|
1.3. Дальние мигранты |
4 |
12,1 |
11 |
33,3 |
18 |
54,6 |
|
2.0. Местоположение района исследова ний в ареале вида |
8 |
14,5 |
16 |
29,1 |
31 |
56,4 |
|
2.1. Центр ареала |
4 |
11,8 |
8 |
23,5 |
22 |
64,7 |
|
2.2. Периферия ареала |
4 |
28,6 |
5 |
35,7 |
5 |
35,7 |
|
2.3. Граница ареала |
0 |
0 |
3 |
42,9 |
4 |
57,1 |
В.А. Паевский (2006, 2008) на основе анализа помесячного распределения количества погибших особей у мигрантов обнаружил, что гибель птиц в периоды миграций не выше, чем в другие периоды годового цикла, а смертность во время зимовок соответствует уровню смертности во все остальные фазы жизни. Детальные исследования (анализ ключевых факторов и выживаемости) на европейских птицах показали, что причинами самых резких снижений их численности были в ряде случаев не нарушения процесса размножения, а снижение выживаемости птиц на территориях зимовок, в особенности африканских. Нет никаких оснований предполагать, что этот вывод не пригоден в отношении южно-азиатских зимовок. В современный период для этих территорий характерно повышение уровня антропогенного пресса на местообитания вследствие огромной концентрации населения и возрастания степени загрязнения природной среды различными ядохимикатами, а также массового отлова мелких воробьиных птиц местным населением в период миграций и на зимовках с целью традиционного использования в пищу.
Исследование закономерностей движения численности видов позволяет не только прогнозировать изменения их популяций, но и получать оперативную информацию о современных изменениях, происходящих в природных комплексах. Такие оценки и прогнозы возможны на основе представлений о том, что многолетняя цикличность – это свойство и естественное состояние природных сообществ, форма их существования и развития (Максимов, 1984).
При рассмотрении результатов изменения численности видов на ключевом участке за продолжительный период наблюдений (свыше 20 лет) выявляются периоды с достоверным ростом или снижением плотности (рис. 1). В целом же за все годы тенденция может быть статистически не значимой, то есть численность вида колеблется случайно или квазициклически.
Раздельный анализ таких тенденций на отдельных высотно-поясных выделах показал, что статистически значимые тренды могут быть обнаружены не только у тех видов, для которых они выявлены для всего ключевого участка, но и у видов со статистически недостоверными суммарными направленностями долговременных изменений. При этом для некоторых видов на разных участках могут вскрываться разнонаправленные статистически значимые тренды.
Пятнистый конек ( Anthus hodgsoni ), например, на соседних (предгорном и горно-лесном)
участках долины р. Езовка имеет разнонаправленные долговременные изменения обилия, а также отрицательный тренд на равнинноморенном участке р. Большой и положительные тенденции на выделах в долине р. Давша. Чиж, обладая общим положительным трендом изменений численности, на прибрежно-равнинном участке р. Езовка обнаруживает отрицательную тенденцию.
У кукши ( Perisoreus infaustus ) разнонаправленные статистически значимые тренды выявлены на предгорном участке долины р. Езовка и прибрежно-равнинном – р. Большая, у обыкновенной пищухи соответственно на соседних равнинном и равнинно-моренном выделах р. Большая. Трехпалый дятел ( Picoides tridactylus ) обнаруживает отрицательный тренд изменений численности на прибрежно-равнинном участке р. Давша и положительный – на предгорном р. Езовка. Для трехпалого дятла она очевидно связана с появлением свежих гарей с усыхающим древостоем на предгорном выделе р. Езовка в середине 1990-х и в начале 2000-х гг. В долине р. Давша, напротив, количество усыхающих деревьев было значительно выше в первую половину периода наблюдений и в последнее десятилетие существенно уменьшилось.
Заключение
Для гнездовой численности видов птиц на ключевом участке Баргузинского хребта характерны разнонаправленные флуктуации обилия, которые связаны с условиями зимовки, определяющими уровень зимней смертности в популяциях, и условиями, складывающимися на местах гнездования ко времени их возвращения и занятия гнездовых территорий. На конкретных временных периодах эти флуктуации могут формировать статистически значимые тренды долговременных изменений. Из 55 видов для 8 отмечен статистически значимый рост обилия, для 16 – снижение и для 31 вида плотность гнездования была относительно стабильной.
Негативные тенденции преобладают у дальних мигрантов. У оседлых видов и у ближних мигрантов имеется равное соотношение положительных и отрицательных трендов. Максимальная доля видов, снижающих гнездовое обилие, относится к периферийной и пограничноареальной группам. Разнонаправленность тенденций изменения гнездового обилия птиц на отдельных выделах указывает на возможность перераспределения населения видов между соседними высотно-поясными участками или речными долинами.
