Многолетняя динамика сообществ макрозообентоса Северного Каспия

Автор: Малиновская Л.В., Зинченко Т.Д.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1-1 т.12, 2010 года.
Бесплатный доступ
Приводится материал многолетних исследований (1935-2002 гг.) количественного развития и распространения донных организмов в Северном Каспии в различные периоды формирования экосистемы моря.
Макрозообентос, распределение, северный каспий
Короткий адрес: https://sciup.org/148205476
IDR: 148205476 | УДК: 574.587(262.81)
Long-term dynamics of macrozoobenthos communities in the North Caspian Sea
There is given the data of perennial researches (1935-2002) of quantitative development and distribution of bottom organisms in the North Caspian Sea within various periods of the sea ecosystem formation.
Текст научной статьи Многолетняя динамика сообществ макрозообентоса Северного Каспия
В течение многовековой истории Каспийского моря происходили многократные смены трансгрессивных и регрессивных фаз положения его уровня. Непостоянство уровенного режима сказывалось на изменениях гидрологогидрохимических характеристик моря и влияло на целый ряд структурных перестроек биосистемы [5, 11].
Анализ многолетней динамики качественных и количественных характеристик донных сообществ позволяет установить основные закономерности их развития в условиях изменения экологических процессов в море, а также прогнозировать состояние кормовой базы промысловых рыб при воздействии комплекса антропогенных факторов, из которых определяющим в современный период является загрязнение водной среды нефтепродуктами. Исследование многолетних изменений донных сообществ Северного Каспия является одной из основных задач экологического мониторинга.
Изучению состава и распределения донных беспозвоночных в Северном Каспии в различные периоды формирования экосистемы моря посвящены исследования Н.Л. Чугунова [30], А.Я. Бирштейна [2], А.А. Шорыгина [31], Н.Н. Спасского [28], Л.Г. Виноградова [4], А.К. Са-енковой [26], В.Ф. Осадчих [19, 20], Яблонской Е.А. [32], Л.В. Малиновской [15] и упоминания в обзорах [6, 9, 22]. Авторы этих работ отмечают, что биомасса и состав бентоса Северного Каспия испытывают межгодовые колебания, которые являются отражением изменений в гидрологическом режиме водоема.
Целью работы является анализ многолетней динамики (1935-2002 гг.) сообществ макрозообентоса, развивающихся в различных природно-экологических условиях Северного Каспия.
МАТЕРИАЛ И МЕТОДИКА
В работе использованы архивные материалы мониторинговых съемок (июнь), проводимых сотрудниками лаборатории кормовой базы и питания рыб Каспийского научноисследовательского института в северной части Каспийского моря (1935-1978 гг.) и результаты собственных исследований (1979 г. по 2002 гг.). Исследовалась акватория моря с глубинами от 1,2 до 30 м.
Пробы бентоса отбирали дночерпателем «Океан-50», с площадью захвата 0,1 м 2 . Содержимое дночерпателя промывали через сито из газа № 14 и фиксировали 4% раствором формальдегида. Обработку материала в лаборатории проводили по общепринятой методике [25].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Состояние бентоса Северного Каспия до падения уровня моря характеризуют данные 1935 г. Средняя биомасса макрозообентоса (табл. 1), основу которой формировали солоноватоводные двустворчатые моллюски родов Dreissena, Adacna, Monodacna , составляла 40,3 г/м 2
Понижение уровня моря привело к сокращению акватории Северного Каспия и его осо-лонению, что особенно резко проявилось в 1938-1940 гг. Было зарегистрировано сокращение, по отношению к 1935 г. биомассы донных беспозвоночных, примерно в 4 раза. Доля слабосолоноватоводных и солоноватоводных моллюсков сократилась в 10 раз.
Одной из основных причин падения биомассы донных сообществ (9,3 г/м 2 ) в указанный период, по мнению Я.А. Бирштейна [2], являлся своеобразный ход половодья р. Волги в 1936 г., который вызвал резкую солевую стратификацию в прилегающей к дельте части моря, сопровождающуюся дефицитом кислорода. Средняя соленость Северного Каспия возросла до 11-12%о. В восточной половине водоема образовались очаги повышенной солености до 15-17^ [3, 13].
В 40-е годы с увеличением водного стока р. Волги (135 м3) и снижением солености вод Северного Каспия (7,63%о), период относительной стабильности уровня моря характеризовался повышением биомассы зообентоса (24,3 г/м2), обусловленной в значительной степени моллюсками слабосолоноватоводного и солоноватоводного комплексов (10,3 г/м2). В этот, по мнению Н.Н. Спасского [28], восстановительный период (1941-1956 гг.), биомасса и запасы организмов макрозообентоса, включая и солоноватоводных моллюсков, оставались примерно в 1,5-2 раза ниже, чем в 1935 г. Уменьшилась плотность заселенных биотопов двустворчатыми моллюсками родов Hypanis, Didacna, Dreissena.
Таблица 1. Динамика биомассы зообентоса, солености воды и некоторых гидрологических характери- стик Северного Каспия в различные периоды 1935-2002 гг.
|
Годы |
Биомасса, г/м 2 |
Уровень моря, м |
Водный сток (апрель-июнь), м 3 |
Соленость, % |
|
1935 |
40,3 |
- 26,65 |
102,6 |
8,74 |
|
1937-1940 |
9,3 |
- 27,51 |
112,8 |
10,31 |
|
1941-1956 |
24,3 |
- 28,05 |
135,5 |
7,63 |
|
1957-1962 |
71,5 |
- 28,27 |
116,6 |
7,52 |
|
1963-1977 |
53,4 |
- 28,41 |
93,4 |
8,32 |
|
1978-2002 |
70,6 |
- 27,51 |
111,7 |
7,13 |
В первые годы после зарегулирования Волги (1957-1962 гг.) в Северном Каспии развитие донных беспозвоночных проходило в условиях уменьшения объ ё ма стока в период паводка, увеличения поступления растворенных органических соединений и взвешенных веществ до величин, достигающих 9 млн. т [10]. Период 1956-1960 гг. характеризовался высокой биомассой фитопланктона Северного Каспия — 2,3 г/м 3 [14]. Из донных организмов было зарегистрировано широкое расселение интродуцированных в 40-е годы из Азовского моря двустворчатых моллюсков Abra ovata и многощетинковых червей Hediste diversicolor . В составе общей биомассы бентоса значительную долю составил «стихийный вселенец» моллюск Myti-laster lineatus, биомасса которого возросла более чем в 5 раз. Обусловленная этими изменениями общая биомасса зообентоса возросла в 3 раза (71,5 г/м 2 ), по сравнению с 1941-1956 гг., варьируя в различные годы от 52,0 г/м 2 (1957 г.) до 89,7 г/м 2 (1961 г.).
В последующий период (1963-1977 гг.), который характеризовался маловодностью, попуски воды в половодье уменьшились до 56,9 км3 (1975). Началось медленное (1963-1971 гг.), а затем интенсивное (1972-1977 гг.) снижение уровня Каспийского моря до отметки примерно - 29,0 БС и повышение средней солености вод Северного Каспия до 11% (1976 г.). Значительно сократился вынос биогенных элементов и увеличилось поступление растворенных органических соединений, что обусловило снижение продукции фитопланктона, в составе которого преобладали крупные водоросли (Spiro-gyra sp., Zygnema sp., Rhizosolenia calcar avis), недоступные для организмов-фильтраторов, в том числе и моллюсков. Осолонение вод в этот период способствовало интенсивному развитию видов морского комплекса, из которых преобладали средиземноморские вселенцы: дву створчатые моллюски — Abra ovata, Cerasto-derma lamarcki, Mytilaster lineatus; многощетин-ковый червь Hediste diversicolor и усоногие ракообразные Balanus improvisus. Начались процессы вытеснения коренной каспийской фауны видами инвазионного комплекса. Так, максимальная биомасса перечисленных видов двустворчатых моллюсков, в отдельные годы достигающая 78% от биомассы всех моллюсков, была зарегистрирована в районах Северного Каспия, находящихся под влиянием среднекаспийских вод с более устойчивой высокой соленостью. Среднемноголетняя биомасса представителей слабосолоноватоводного и солоноватоводного комплексов (моллюски Dreissena poly-morpha, Adacna vitrea, A. angusticostata) сократилась с 16,7 г/м2 (1957—1962 гг.) до 8,5 г/м2 [19]. Что касается биомассы и запасов ракообразных, каспийских полихет и личинок хиро-номид, то эти величины также снизились, по сравнению с 1957-1962 гг., но были выше, чем до зарегулирования стока Волги, что связано с эвригалинностью массовых видов и обитанием их в прибрежном мелководье Северного Каспия, где они обитают в условиях обеспеченнно-сти пищей.
Общая биомасса донных беспозвоночных в 1963-1977 гг., по отношению к предыдущему периоду (табл. 1), снизилась до 53,4 г/м 2 , с колебаниями от 30,1 г/м 2 (1967 г.) до 60,9 г/м 2 (1963 г.)
По нашим данным, например, биомасса личинок хирономид (Diptera, Chironomidae) в течение всего маловодного периода мало изменялась и не превышала 0,5 г/м2, тогда как в многоводные годы наблюдается тенденция к ее увеличению.
Подъ ё м уровня (рис. 1) Каспийского моря регистрируется начиная с 1978 г., что обусловило существенные изменения в развитии и распределении сообществ макрозообентоса. В
1978-2002 гг. водность р. Волги в весеннее по- среднем составила 111,7 км 3 , т [11]. Повышение ловодье колебалась от 61,6 до 159,9 км 3 и в стока волжских вод способствовало уменьше
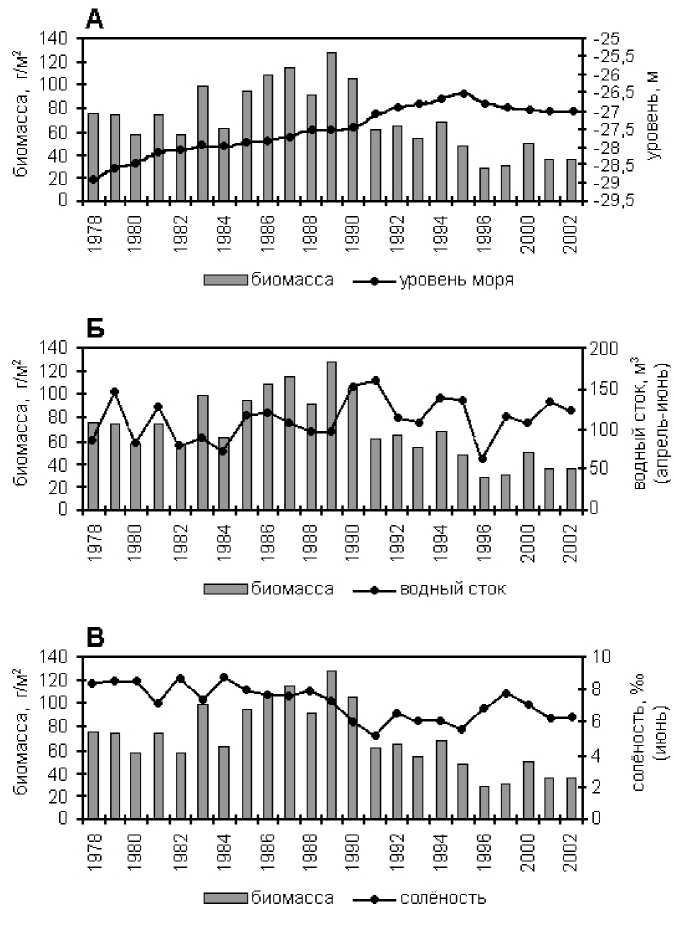
Рис. 1. Многолетняя динамика биомассы макрозообентоса Северного Каспия в периоды изменения уровня моря, водного стока, солености
нию средней солености, в отдельные годы до 5,25%о и расширению опресненных площадей в Северном Каспии, что происходило на фоне регистрируемого периодического увеличения выноса в море загрязняющих веществ [22-24] и возросших концентраций биогенных веществ, преимущественно минеральных форм. Среднегодовые концентрации минерального фосфора в отдельные годы достигали 90 мкг/л. Среди минеральных форм азота преобладали нитраты (до 92%) с максимальной концентрацией до 920-1280 мкг/л, что свидетельствует о значительном антропогенном воздействии [12].
В фитопланктоне доминировали виды пресноводного и слабосолоновато-водного ком плекса, в составе которого, начиная с 2000 г. преобладали мелкоклеточные водоросли (8085% от общей биомассы альгофлоры) [1]. В этих условиях наблюдался широкий размах колебаний средней биомассы бентоса Северного Каспия — от 27,3 г/м2 в 1996 г. - до 128,2 г/м2 (1989 г.), что обусловлено, в значительной степени, динамикой стока пресных вод.
За 25 летний период исследований регистрировались годы малой и большой водности (рис. 1). В маловодные годы регистрируется сокращение общей биомассы бентоса и отдельных его ценозов, примерно в 1,6 раза, преимущественно за счет организмов слабо - и солоноватоводного комплексов. В многоводные годы, при низкой солености и благоприятно складывающихся, за счет роста выноса биогенных элементов, трофических условий количественные показатели зообентоса, в большинстве случаев, возрастают (рис. 1), что подтверждается ранними исследованиями Л.Г. Виноградова [4], В.Ф. Осадчих [21], Е.А. Яблонской [32]. В период повышения уровня моря (19781990 гг.) интенсивность развития бентоса была в 1,6 раза выше, чем в период понижения уровня моря [17, 33]. Средняя биомасса хиро-номид в 1981-1984 гг. возросла и составила в Северном Каспии 1,32 г/м2. Доминирующими, среди хирономид, были эндемики Каспия Chi-ronomus albidus, а субдоминантами, с учетом численности, частоты встречаемости и численности личинок, зарегистрированы представители пресноводного и слабосолоновато-водного комплекса, распространенные в опресненных районах западного побережья Северного Каспия — Cryprochironomua gr. defectus, Polypedilum convictum, P. nubeculosujm, Glyptotendipes gr. gripekoveni, Cladopelma lateralis, Paratanytarsus confusus, Dicrotendipes gr. tritomus [7]. По данным Л.В. Смирновой [27], биомасса общего бентоса в Северном Каспии (июль) увеличилась от 25,9 (1986г.) до 64,2 г/м2 (1990), на западе — от 34,6 (1988 г.) до 84,7 (1990 г.), на востоке от 12,3 (1986 г.) до 37,1 (1990 г.). Лидирующее положение занимали олигохеты, среди ракообразных — амфиподы. Руководящими видами среди моллюсков были мити-лястр и абра.
По результатам исследований за 1986-1997 гг. наблюдалась тенденция количественного развития важных в кормовом отношении организмов бентоса, что положительно отразилось на нагуле рыб-бентофагов. Исключение составил 1996 г., который характеризовался снижением уровня моря, экстремально малой водностью (61,6 км 3 ) и повышением солености (6,72%о) [10], что отразилось на развитии зообентоса, средняя биомасса которого, по отношению к 1994-1995 гг. (многоводным годам), сократилась в 2,4 раза, не превышая 27,4 г/м 2 . Отмеченное снижение уровня моря является, вероятно, кратковременной осцилляцией на фоне общего повышения уровня моря [29].
Межгодовые изменения биомассы бентоса в период трансгрессии моря определяли моллюски (до 80% биомассы всего бентоса). В их составе до 1990 г. доминировали инвазийные двустворчатые моллюски Mytilaster lineatus (до 40% биомассы и запаса моллюсков); в 1980, 1982, 1990 и 1991 гг. преобладали Abra ovata, а в 1992-2002 гг. преимущественное развитие имели Adacna angusticostata. Средняя биомасса двустворчатых моллюсков в многолетнем ряду измерений варьировала от 15,5 г/м2 (1996 г.) до 105,0 г/м2 (1989 г), а численность изменялась в диапазоне 231 экз2/м (2002 г.) -12,5 тыс. экз./м2 (1989 г.).
Особенностью современного периода (начиная с 2001 г.) является значительное снижение численности моллюсков, обусловленное сокращением популяционной доли молоди в сообществах бентоса [16]. В структуре зоопланк-тонных сообществ, по данным за июнь 20012002 гг., численность велигеров моллюсков также сократилось более чем в 4,5 раза [1].
Одной из основных причин снижения молоди Bivalvia является расселение с 2000 г. в водах Каспийского моря гребневика Mnemiopsis leidyi [8], который, активно потребляя меропланктон, выедает велигеров моллюсков. Загрязнение вод Северного Каспия различными поллютантами также оказывает негативное воздействие на донные сообщества, приводя к снижению его продуктивности [22].
Динамика количественных показателей донных ракообразных в 1978-2002 гг. носила флуктуирующий характер, который был обусловлен гидролого-гидрохимичесим режимом водоема, антропогенным воздействием и трофической нагрузкой со стороны бентофагов, особенно молоди урожайных поколений карповых рыб. Средняя биомасса ракообразных была достаточно высока — от 2,9 г/м 2 до 9,9 г/м 2 (1998-1980 гг.). За годы трансгрессии моря среди ракообразных по биомассе доминировали корофииды (57%) и гаммариды (26%). Кумо-вые рачки преобладали в 1982, 1992, 1995 и 1996 гг.
Биомасса Oligochaeta + Polychaeta колебалась от 2,8 (1997 г.) до 15,1 г/м 2 (1989 г.). Высокие показатели биомассы обусловлены в основном развитием олигохет (80% от общей биомассы бентоса), что служит косвенным показателем процесса эвтрофирования в зоне устьевого волжского пространства Северного Каспия [12]. Их численность на заиленных биотопах в отдельные годы достигала 8 тыс. экз./м 2 .
В распределении общего бентоса между западными и восточными пастбищами в течение всего периода наблюдений приоритет имел западный район. Так, в годы низкого уровня моря (1973-1977 гг.) биомасса донных беспозвоночных на западе Северного Каспия была почти в 2 раза выше, чем на востоке. В многоводный период (1980-1997 гг.) различия еще более увеличились (в 3,3 раза), что обусловлено резким снижением в восточном районе биомассы галофильных видов, особенно моллюска Di-dacna trigonoides, вызванного распреснением моря. Исключение составляет маловодный 1996 г., когда средняя биомасса бентонтов в обоих районах моря имела сравнимые величины (в западном районе — 30,9 г/м2, в восточном — 24,3 г/м2). Начиная с 2001 г. вновь регистриру- ется резкий спад количественных показателей моллюсков в западном районе, обусловившее снижение, примерно в 2 раза, общей биомассы бентоса.
Таким образом, донные сообщества Северного Каспия в многолетнем ряду наблюдений (1935-2002 гг.) претерпевали существенные качественные и количественные изменения, обусловленные динамикой объема половодья, уровенного режима моря, солености и трофического статуса моря. Каждый из этих факторов в определенный период времени может играть существенную роль. Были установлена положительные корреляционные связи между биомассой бентоса (1978-2002 гг.) и такими параметрами, как величина весеннего половодья (R = 0,663 при Р = 0,001), уровень моря в пределах его колебаний от 26,54 до 28, 88 (R = 0,658 при P = 0,001) и соленость воды (R = 0,800 при P=0).
Из общей закономерности выпадают 20002002 гг., когда формирование зообентоса проходило в новых экологических условиях стабилизации уровня Каспийского моря, сопровождающейся распреснением северокаспийских вод, высоким антропогенным воздействием, связанным с загрязнением вод поллютантами различной природы, процессом эвтрофирова-ния, а также продолжающимся «биологическим загрязнением» экосистемы моря, оказывающим влияние на взаимоотношения между гидробионтами всех трофических звеньев (рис. 1).
Несомненно, что результаты исследований многолетних количественных изменений донных сообществ позволят дать адекватную оценку запасов кормовых ресурсов и прогнозирования их состояния, что особенно актуально в связи с возрастающей хозяйственной деятельностью и поиском эффективных, экономически и экологически обоснованных способов эксплуатации Северного Каспия, прогнозирования направленных возможных изменений его экосистемы при различных формах антропогенного воздействия, сопровождаемых, в том числе, и неизбежными инвазионными процессами.
Работа выполнена при финансовой поддержке программы фундаментальных исследований Президиума РАН «Биологическое разнообразие»: раздел «Особенности экологии и динамики чужеродных видов гидробионтов (зоопланктон, зообентос, рыбы, паразиты рыб) в водоемах Средней и Нижней Волги».
Список литературы Многолетняя динамика сообществ макрозообентоса Северного Каспия
- Ардабьева А.Г., Тарасова Л.И., Малиновская Л.В. Кормовая база Северного Каспия в 2003 г.//Рыбохозяйственные исследования на Каспии. Результаты НИР за 2003 г. Астрахань, 2004. С. 112-123.
- Бирштейн А.Я. Годовые изменения бентоса Северного Каспия//Зоол. журн. 1945. Т. 24, вып. 3. С. 133-147.
- Винецкая Н.И. Многолетние и сезонные изменения гидрохимического режима Северного Каспия до зарегулирования стока Волги//Тр. КаспНИРО, 1962. Т. 18. С. 4-90.
- Виноградов Л.Г. Многолетние изменения северокаспийского бентоса//Тр. ВНИРО. 1959. Т. 38. Вып. 1. С. 241-274.
- Гурский Ю.Н., Сузюмов А.Е., Артамонов В.И., Золотых Е.Б. Некоторые результаты комплексного геолого-геохимического изучения Каспийского мелководья//Комплексные исследования Каспийского моря. М.: Наука, 1974. Вып. 4. С. 81-92.
- Даирова Д.С., Зинченко Т.Д. История исследований макрозообентоса Нижней Волги и ее дельты//Изв. Самар. НЦ РАН. 2003. Спец. вып. 1. С. 91-101.
- Зинченко Т.Д., Алексевнина М.С. Изменения хирономидофауны (Diptera, Chironomidae) в низовьях дельты Волги и в Каспийском море в связи с подъемом его уровня//Экология, эволюция и систематика хирономид/ред. Н.А. Шобанов и Т.Д. Зинченко. Борок, Тольятти: ИБВВ и ИЭВБ РАН, 1996. С. 154-165.
- Камакин А.М., Студиникина Ю.Б. Распределение вселенца Mnemiopsis leidyi в Каспийском море.//Рыбохозяйственная наука на Каспии: задачи и перспективы: междунар. конф., посвящ. 40-летию ГУДП «Дагестанское отделение КаспНИРХ». Астрахань: КаспНИРХ, 2003. С. 81 -84.
- Карпинский М.Г. Экология бентоса Среднего и Южного Каспия. М.: Изд-во ВНИРО, 2000. 283 с.
- Катунин Д.Н. Северный Каспий. Гидрохимические условия//Каспийское море. Гидрология и гидрохимия. М.: Наука, 1986. С. 128-142.
- Катунин Д.Н., Егоров С.Н., Кашин Д.В., Хрипунов И.А., Беспарточный Н.П. Кравченко Е.А., Никотина Л.Н., Железцова Е.Г., Фесенко В.И., Азаренко А.В. Основные черты гидролого-гидрохимического режима нижнего течения р. Волги и Каспийского моря.//Вопросы промысловой океанологии. М.: Изд-во ВНИРО, 2004. Вып. 1. С. 69-96.
- Катунин Д.Н., Егоров С.Н., Хрипунов И.А., Кашин Д.В., Галушкина Н.В., Кравченко Е.А. Основные особенности гидролого-гидрохимического режима р. Волга и Каспийского моря в трансгрессивный период.//Журн. «Рыбное хозяйство», 2007. № 3. С. 75-79.
- Катунин Д.Н., Хрипунов И.А. Многолетнее распределение температуры, солености и прозрачности вод Северного Каспия//Тр. КаспНИРХ. М.: Пищевая промышленность, 1976. С. 128-142.
- Левшакова В.Д. Кормовая база рыб бетофагов//Труды ВНИРО. 1978. Т. 108. С. 81-98.
- Малиновская Л.В. Особенности развития зообентоса Северного Каспия в условиях продолжающегося подъема уровня моря//Тез. докл. Х Междунар. конф. по промысловой океанологии. (Санкт-Петербург, 20-23 мая 1997 г.) М.: ВНИРО, 1997. С. 82-83.
- Малиновская Л.В. Многолетняя динамика развития моллюсков Северного Каспия//Междунар. конф. «Рыбохоз. наука на Каспии: задачи и перспективы», посвящ. 40-летию ГУДП «Дагестанское отделение КаспНИРХ». Астрахань: Изд-во КаспНИРХ, 2003. С. 118-121.
- Малиновская Л.В. Зообентос Северного Каспия в период подъема уровня моря.//Атореф. диссерт. на соиск. учен. степени канд. биол. наук. Тольятти, 2007. 20 с.
- Научные основы устойчивого рыболовства и регионального распределения промысловых объектов Каспийского моря/ред. В.Н.Беляева, В.П. Иванов, В.К. Зиланов. М.: ВНИРО, 1998. 167с.
- Осадчих В.Ф. Бентос Северного Каспия в условиях зарегулирования стока р. Волги//Зоол. журн. Т. 42. Вып. 2. 1963. С. 184-196.
- Осадчих В.Ф. Динамика биомассы зообентоса Северного Каспия в 1966-1972 гг.//Труды ВНИРО. 1974. Т. 101. С. 36-47.
- Осадчих В.Ф. Влияние режима весеннего половодья на развитие зообентоса Северного Каспия//Гидробиол. журн. 1980. Т. XVI. Вып. 6. С. 17-25.
- Панин Г.Н., Мамедов Р.М., Митрофанов И.В. Современное состояние Каспийского моря/отв. ред. М.Г.Хубларян//Ин-т Вод проблем РАН. М.: Наука, 2005. 356 с.
- Попова О.В., Попова Э.С. Многолетняя динамика выноса фенолов в составе волжского стока и распределение по акватории Северного Каспия // Современные проблемы водной токсикологии // Тез. межд. конф. Борок: ИБВВ РАН 2005. С. 109.
- Попова О.В., Чуйко Е.В. Многолетнее изменение содержания тяжелых металлов в поверхностных водах Северного Каспия и факторы их определяющие//Тез. межд. конф. Борок: ИБВВ РАН, 2005. С. 110.
- Романова Н.Н. Методические указания к изучению бентоса южных морей СССР. М.: ВНИРО, 1983. 13 с.
- Саенкова А.К. Сезонная динамика бентоса Северного Каспия//Тр. КаспНИРО. 1959. Т. 15. С. 56-104.
- Смирнова Л.В. Состояние зообентоса Северного Каспия в конце 80-х годов//Морские гидробиологические исследования. М.: ВНИРО, 2000. С. 103-110.
- Спасский Н.Н. Состояние и изменение бентоса Северного Каспия в период с 1940-1945 гг.//Зоол. журн. 1948. Т. 27, Вып. 3. С. 193-204.
- Ходоревская Р.П., Рубан Г.И., Павлов Д.С. Поведение, миграции, распределение и запасы осетровых рыб Волго-Каспийского бассейна. М.: Тов. научн. изданий КМК, 2007. 242 с.
- Чугунов Н.Л. Опыт количественного исследования продуктивности донной фауны в Северном Каспии и типичных водоемах дельты р. Волги//Труды Астраханской ихтиол. лаб. 1923. Т. 5. Вып. 1. С. 107-192.
- Шорыгин А.А. Изменение количества и состава бентоса Северного Каспия в 1935-1940 гг.//Зоол. Журн. 1945. Т. 24. Вып. 3. С. 148-159.
- Яблонская Е.А., Осадчих В.Ф. Влияние окаенологических условий на формирование бентоса//Гидрометеорология и гидрохимия морей. Каспийское море. С-П.: «Гидромедиздат», 1996. Т. VI, Вып. 2. С. 263-278.
- Malinovskaja L.V., Filippov A.A., Osadchiky V.F., Aladin N.V. Benthic macroinvertebrates of the nortkern Caspian Sea During recent rises in water -level//International Journal of Salt Lake Research. 1998. V. 7. P. 211-233.
