Мобильные элементы Oryza sativa L. (обзор)
")
Автор: Илюшко М.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 1 т.61, 2026 года.
Бесплатный доступ
Ожидается, что глобальные изменения окружающей среды уже в ближайшие 50-70 лет приведут к созданию качественно новой среды обитания для многих организмов, затрагивая агроценозы, в силу чего адаптация сельскохозяйственных культур к климатическим условиям становится сложной селекционной задачей. Рис (Oryzasativa L.) входит в число лидирующих культур в мировом земледелии и служит модельным биологическим объектом (N. Kurata с соавт., 2002). Действие мобильных элементов, или транспозонов (transposable elements, TEs), повышая изменчивость доместицированных видов, способствует фенотипическому разнообразию и эффективности искусственного отбора (В.И. Глазко с соавт., 2022). Представленный обзор систематизирует знания о мобильных элементах риса и их применимости в селекции растений. Приводится характеристика TEs, свойственных родуOryza L. и культурному видуO. sativa(A.F. Tufan с соавт., 2020; S.Y. Jiang с соавт., 2013). Рассматривается вклад мобильных элементов в эволюцию рода. Выявлено, что LTR (long terminal repeat) ретротранспозоны, будучи главными компонентами геномов Oryza, при полиплоидизации распространяются в первую очередь, влияя на их размер вне зависимости от типа субгенома. Ретротранспозоны суперсемействаGypsy существенным образом коррелируют с размером геномов (A. Fornasiero с соавт., 2025). Полиморфизм инсерций мобильных элементов позволил сделать заключение о полифилетичном происхождении подвидов риса japonica и indica и группы Aus/boro в отличие от прежних представлений об их монофилетичной эволюции, основанных на данных о ключевом локусе доместикации Sh4 (локус осыпаемости) (M. Carpentier с соавт., 2019). С помощью TEs получены практические результаты по повышению продуктивности растений O. sativa. При облучении семян риса пучками тяжелых ионов высокой энергии созданы три мутантные формы с транспозициями Dasheng, mPing, Osr13 и RIRE2рядом с некоторыми генами, в частности OsSPCH2 и OsCPS2. В результате возросли размеры и масса семян, сократился период до цветения растений (X. Wen с соавт., 2023). В ряде исследований выявлены гены, активация которых TE не приводит к фенотипическим изменениям или сопровождается нежелательными последствиями для хозяйственно ценных признаков (R. Akakpo с соавт., 2020; X. Wen с соавт., 2023). Мутации, вызванные транспозиционной активностью, наследуются в ряду поколений до M5 (M. Komatsu с соавт., 2003). Число копий мобильных элементов играет решающую роль в проявлении агрономически важных признаков. При сравнении двух почти изогенных линий риса с одной и тремя копиями HUO доказано снижение продуктивности метелки (число и масса семян) и устойчивости к бактериальному ожогу (Xanthomonasoryzae) у трехкопийной линии (Y. Peng с соавт., 2019). Предлагается детектировать стрессовую толерантность O. sativa, используя IRAP маркеры мобильности Tos17. С помощью технологии CRISPR/Cas9 в культуре in vitro получен регенерант риса с целевой мутацией, вызвавшей делецию ретротранспозона Tos17 на 7-й хромосоме, что открывает перспективы геномного редактирования хозяйственно ценных генов, поврежденных TEs (H. Saika с соавт., 2019, Y. Luo с соавт., 2020). Таким образом, на рисе O. sativa показаны пути практического применения знаний о мобильных генетических элементах в селекции сельскохозяйственных культур. Расширение методов создания исходного материала и объединение с уже имеющимися возможностями позволит селекции более гибко реагировать на динамично меняющийся климат.
Oryzasativa, мобильные генетические элементы, эволюция, культура клеток in vitro, селекция
Короткий адрес: https://sciup.org/142247330
IDR: 142247330 | УДК: 633.18:631.524.85.02:577.2 | DOI: 10.15389/agrobiology.2026.1.57rus
Oryza sativa L. transposable elements (review)
It is expected that global environmental changes in the next 50-70 years will lead to the creation of a qualitatively new habitat for many organisms, affecting agrocenoses, due to which the adaptation of agricultural crops to climate change becomes a complex breeding task. Rice Oryza sativa L. is one of the leading crops in world agriculture and a model biological object (N. Kurata et al., 2022). The action of mobile elements or transposons (TEs), increasing the variability of domesticated species, contributes to phenotypic diversity and the effectiveness of artificial selection (V.I. Glazko et al., 2022). The presented review systematizes knowledge about mobile elements of rice and their applicability in plant breeding. The paper provides a characteristic of TEs inherent in the genus Oryza L. and the cultivated species O. sativa (A.F. Tufan et al., 2020; S.Y. Jiang et al., 2013). The mobile elements contribution to the evolution of the genus is considered. It was revealed that LTR retrotransposons, being the main components of the Oryza genomes, are distributed primarily during polyploidization, affecting their size regardless of the subgenome type. In particular, Gypsy superfamily retrotransposons significantly correlate with the genome size (A. Fornasiero et al., 2025). Polymorphism of mobile element insertions allowed to conclude that the rice subspecies japonica and indica and the Aus/boro group are polyphyletic in origin, in contrast to previous ideas about their monophyletic evolution based on data on the key domestication locus Sh4 (shedding locus) (M. Carpentier et al., 2019). Using TEs, practical results were obtained to increase the productivity of O. sativa plants. Namely, three mutant forms with transpositions of Dasheng, mPing, Osr13, and RIRE2 near some genes, in particular, OsSPCH2 and OsCPS2, were created by irradiating rice seeds with high-energy heavy ion beams. As a result, the size and weight of seeds increased, and the period before plant flowering decreased (X. Wen et al., 2023). A number of studies have identified genes whose activation by TEs does not lead to phenotypic changes or leads to undesirable consequences for economically valuable traits (R. Akakpo et al., 2020; X. Wen et al., 2023). Mutations caused by transposition activity are inherited in a generations number up to M5 (M. Komatsu et al., 2003). The copy number of mobile elements plays a decisive role in the manifestation of agronomically important traits. When comparing two near-isogenic rice lines with one and three copies of HUO, a decrease in panicle productivity (seeds number and weight) and resistance to bacterial blight (Xanthomonas oryzae) was proven in the three-copy line (Y. Peng et al., 2019). It is proposed to detect O. sativa stress tolerance using IRAP markers of Tos17 mobility. Using the CRISPR/Cas9 technology in vitro culture, a rice regenerant with a target mutation causing deletion of theTos17 retrotransposon on the seventh chromosome was obtained, which opens up prospects for genomic editing of economically valuable genes damaged by TEs (H. Saika et al., 2019; Y. Luo et al., 2020). Thus, in the recent period, ways of practical application of knowledge about mobile genetic elements in crop breeding have been shown on O. sativa rice. Expanding the methods for prebreeding and combining it with existing capabilities will allow breeding to respond more flexibly to a dynamically changing climate.
Текст научной статьи Мобильные элементы Oryza sativa L. (обзор)
На геном растений в значительной степени влияет наличие и активность мобильных генетических элементов, известных как транспозоны (transposable elements, TEs). Они способны перемещаться и реплицироваться внутри генома хозяина, внося вклад в генетическую изменчивость и эволюционные процессы (1-3).
Мобильные генетические элементы эукариот разделяют на два основных класса: ретротранспозоны и ДНК-транспозоны. Ретротранспозоны (класс I) распространяются посредством копирования и вставки в процессе транскрипции с последующим использованием обратной транскриптазы, а ДНК-транспозоны (класс II) — посредством «вырезания и вставки» при помощи транспозаз. Класс I представлен автономными эндогенными ретровирусами (EVR), содержащими длинные концевые повторы (long terminal repeat, LTR), длинными диспергированными ядерными элементами (long interspersed nuclear elements, LINE), а также неавтономными элементами — короткими диспергированными ядерными элементами (short interspersed nuclear elements, SINE) и комплексными ретротранспозонами (composite retrotransposon SVA). Класс II включает автономные элементы, кодирующие DDE-аминокислотный мотив, типичный для транспозазы/интегразы большинства семейств автономных ДНК-транспозонов, а также неавтономный элемент — миниатюрный инвертированный повтор транспозируемого элемента (miniature inverted repeat transposable element, MITE) (1, 2, 4). У растений больше распространены эндогенные ретровирусы, а у животных выше частота встречаемости SINE (1, 5). В геномах растений наблюдается наибольшая представленность LTR ретротранспозонов (2, 6, 7), относящихся к суперсемействам Copia и Gypsy (7).
При воздействии внешних стрессовых факторов активность TEs часто заметно увеличивается, особенно в генеративных клетках растений (2, 4, 7). Для предотвращения разрушительного действия ретротранспозонов геном растений предусматривает стратегию сдерживания экспансии мобильными элементами. Процесс инактивации мобильных элементов происходит посредством механизмов транскрипционного и посттранскрипционного сайленсинга. В случае активации возможна различная судьба TEs в геноме с эволюционно генетическими последствиями (7).
Рис ( Oryza sativa L.) — уникальный вид двойного назначения. Во-первых, это одна из самых востребованных сельскохозяйственных культур, рисом питается половина населения планеты. Во-вторых, это признанный модельный биологический объект с небольшим геномом ( ∼ 389-430 Mbp), который был полностью секвенирован сначала для сортов Nipponbare (subsp. japonica ) и 93-11 (subsp. indica ) (референсные геномы) (8, 9). Впоследствии исследования расширились до 3000 сортов с применением технологии быстрого прочтения (в частности, коротких Illumina-сиквенсов) (10).
Настоящий обзор систематизирует знания о мобильных элементах риса и их применимости в селекции культурного вида Oryza sativa.
Мобильные элементы — широко распространенная и наиболее динамичная часть генома (11). У растений с маленьким размером генома, к которым относится O. sativa , содержание TEs относительно невысокое и, по разным оценкам, составляет от 19,6 до 40,0 % (11-13). Локализованы мобильные элементы во всех частях хромосом. У риса выявлено обилие центромерных ретротранспозонов во всех хромосомах, что не характерно для других культур (14, 12). Половина мобильных элементов располагается примерно в 8 % открытых (доступных) хроматиновых регионах (15).
У O. sativa в рамках проекта «3000 геномов риса» идентифицированы 60743 локуса мобильных элементов, которые принадлежат 496 семействам TEs (16). В исследованиях представителей рода Oryza в большей степени фигурируют LTR ретротранспозоны суперсемейств Gypsy и Copia (10, 17, 18). Их доля составляет 5,24-44,54 % на геном вида (18).
O. sativa содержит как минимум 109 Мб LTR ретротранспозонов, что составляет 29 % генома (12), то есть большую часть всех мобильных элементов. В целом их насчитывают более 13000 (12, 19). У O. sativa LTR представлены 340 семействами и 3663 локусами, полностью покрывающими LTR ретротранспозоны в геноме. Лидируют по числу копий семейства Das-heng (450 копий) и Hopi (430 копий), остальные семейства представлены относительно небольшим, до 180, числом копий (6) (рис. 1). Из LINE мобильных элементов у риса встречаются TEs семейств Karma и Lullaby (5, 10, 20), активными ДНК-транспозонами считаются dTok, nDart и mPing/Pong
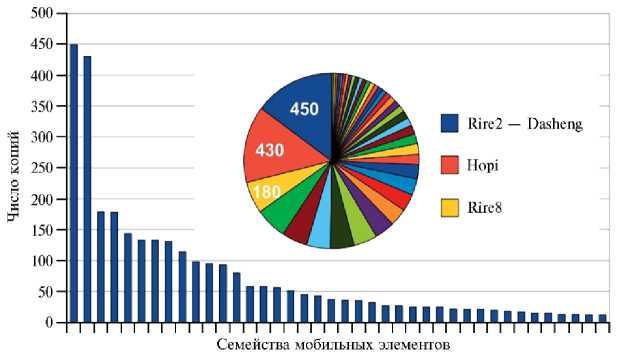
Рис. 1. Распределение числа копий по семействам LTR-транспозонов в геноме Oryza sativa L. Круговая диаграмма демонстрирует долю всех семейств LTR-транспозонов в геноме, указано число копий трех наиболее распространенных семейств. Всего представлены первые 40 семейств (6).
О. malampuzhaensis ВВ I
О. minuta ВВ I
O. iatifolia DD I----
O. australiensis EE I
O. coarctata LL Г
О. schlechteri LL[HH\ E
0. tongiglumis НН E
O. rid ley i НН E
O. longiglumis J J E
0. ridleyi J J E
O. meyeriana GG Г
0. coarctata KK I
0. schlechteri KK I
0. malampuzhaensis CC
O. minuta CC
O. alia CC
O. grandiglumis CC
O. Iatifolia CC
O. alta DD I O. grandiglumis DD I
□ Non-ТЕ content □ hAT
■ САСТА
■ Harbinger
□ Mutator ■ Mariner ■ LRT Copia В LRT Gypsy В LRT unknown □ Unspecified
Размер, Мб
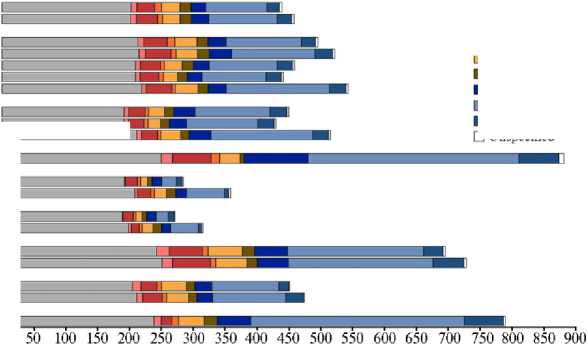
Рис. 2. Количество основных классов мобильных элементов (в мегабазах) рода Oryza L. Содержимое, не относящееся к транспозонам, обозначено серым цветом. Субгеномы вида упорядочены по типу генома ( BB , CC , DD , EE , KK , LL , HH , JJ , GG ) (26).
Число копий различных ретротранспозонов в геноме риса сортоспецифично: от одной до нескольких сот (21). Так, сорт Nipponbare несет две копии TEs Tos17 (4114 п.н.) на гаплоидный геном (22).
При изучении активности мобильных элементов O. sativa применяют несколько подходов: позиционное клонирование генов, измененных инсерциями TEs (например, hAT или Houba); изучение полиморфизмов TEs-инсерций с использованием транспозонов-кандидатов TEs (mPing и Pong); транскрипционные исследования кандидатов TEs с использованием праймеров, таргетирующих консервативные домены (Tos17); полногеномный транскриптомный анализ (Lullaby); полногеномное секвенирование с дальнейшим анализом различными биоинформатическими методами (23).
LRT ретротранспозоны риса представлены в базе данных RetrOryza (7). RTRIP (Rice Transposon Insertion Polymorphism) — база данных, содержащая информацию обо всех мобильных элементах в 3000 сортов риса из 89 стран, которые классифицированы в пять сортовых групп: indica, aus/boro, basmati/sans, tropical japonica, temperate japonica (16).
Вклад мобильных элементов в эволюцию Oryza L. Мобильные элементы имеют свойства перемещаться, самовоспроизводиться и утрачиваться внутри генома, что ведет к его диверсификации и адаптации. Показано, что изобилие мобильных элементов приводит к расширению генома, они также участвуют в регуляции экспрессии генов (1, 4). TEs способны преодолевать межвидовые барьеры посредством горизонтального переноса, приводя к соответствующим эволюционным последствиям (11). Эти свойства мобильных элементов легли в основу изучения родства между различными группами растений. O. sativa вовлекался в эволюционные исследования в качестве представителя рода и вида.
Транспозиционные события четырех специфичных для O. sativa LTR-ретротранспозонов (Hopi, Houba, Osr30 и RIREI) изучены с использованием IRAP-маркеров у представителей четырех родов Poaceae : O. sativa , Brachypo-dium distrachyun (L.) P. Beauv., Hordeum vulgare L., Triticum aestivum L. Полиморфизм отсутствовал у H. vulgare и T. aestivum . Небольшой полиморфизм встречался среди растений B. distrachyun по трем TEs. Исследованные ретротранспозоны еще активны и обусловливают геномный полиморфизм между растениями O. sativa , в то время как у других видов существуют различные механизмы их сайленсинга. Несмотря на то, что у четырех видов был общий предок, рис отделился от него более 40 млн лет назад. Представленный результат может означать, что механизмы, репрессирующие транспозиционные события у ретротранспозонов, возникли после дивергенции риса (11).
Род Oryza состоит, по разным оценкам, из 23-27 видов, два из которых культурные (17, 24, 25). Среди видов Oryza есть диплоидные и полиплоидные, размер геномов которых различается более чем в 3 раза (18, 24). LTR-ретротранспозоны одного семейства распределяются неодинаковым образом у 12 различных по плоидности представителей рода ( O. alta , O. Aus-traliensis , O. brachyanta , O. coarctata , O. glaberrima , O. granulate , O. minuta , O. nivara , O. officinalis , O. punctate , O. ridleyi , O. rufipogon ). Пул LTR мобильных элементов в значительной степени сохранен и распространен в Oryza , являясь древним предшественником событий видообразования, которые привели к происхождению рода. В целом, именно LTR-ретротранспозоны, будучи главными компонентами геномов Oryza , при полиплоидизации распространяются в первую очередь, влияя на их размер (рис. 2). Ретротранспозоны суперсемейства Gypsy существенным образом коррелируют с размером геномов (18). Наибольшую долю из элементов Gypsy представляют два семейства — RIRE2 и Atlantys (24).
За время дивергенции предков аллополиплоидов в их геномах происходила взрывная амплификация транспозонов одних семейств и исчезновение всех или большей части последовательностей из семейств транспозонов общего предка. Так, из 33 семейств ретротранспозонов в геноме O. punctata шесть семейств, занимающих 25 млн п.н. (6,3 % генома), не найдены в геномах других видов риса. Сравнительная характеристика LTR-ретротранспозонов восьми наиболее близкородственных видов из группы родства O. sativa показала, что последовательности теряются из генома со средней скоростью 3620 п.н./млн лет на транспозон. Экстраполируя эти данные, можно предсказать уменьшение генома O. sativa subsp. japonica на четверть без следующего взрыва транспозиций в течение 3-4 млн лет, что в полной мере касается всех видов с субгеномом AA в пределах Oryza (25, 27). Тем не менее время инсерций и накопление LTR ретротранспозиций не зависит от типа субгенома, весь комплекс мобильных элементов участвует в изменении размеров гомеологичных геномов рода Oryza (26). Кроме того, не все TEs ведут себя одинаково: миниатюрные терминальные ретротранспозоны (TRIMs), локализованные в основном около генов, высококонсервативны в геномах 13 видов риса (25).
Происхождение O. sativa остается дискуссионным вопросом, несмотря на значительную изученность вида современными методами. Ключевой локус доместикации Sh4 , ответственный за прочность крепления зерновки риса, то есть отсутствие осыпаемости (28, 29), обнаружен у подвидов japonica и indica , что свидетельствует в пользу монофилетичного происхождения культурного риса (10). Полиморфизм инсерций мобильных элементов лег в основу изучения внутривидовой изменчивости O. sativa . Сиквенсы 3000 геномов риса проанализированы по 32 семействам ретротранспозонов. В результате выдвинуто предположение, что происхождение культурного вида O. sativa связано с тремя событиями доместикации. В позднем неолите примерно 300-800 тыс. лет назад от дикого вида O. rufipogon еще до одомашнивания риса отделились два подвида — japonica и indica . Несколько позже, примерно 230-540 тыс. лет назад, от подвида indica отделилась еще одна группа — Aus/boro (10). То есть в этой логике вид O. sativa какое-то время существовал в природе, распространялся и лишь потом был окультурен человеком. Частота инсерций мобильных элементов различается. Некоторые семейства, например Houba, демонстрируют низкую частоту инсерций, что предполагает недавнюю транспозиционную активность; частота инсерций TEs семейства Hopi выше из-за продолжающихся транспозиций после доместикации; в семействах TEs с большинством инсерций, например Dash-eng, допускают более продолжительную активность. При этом агрономическая история O. sativa привела к активации и увеличению числа копий некоторых мобильных элементов семейства Tos17 (20).
В основу заключений об эволюционных событиях и выявления филогенетического родства видов берется активность мобильных элементов. Сформировано представление, что их взрывное действие ведет к увеличению размеров геномов, играя важную структурную и функциональную роль. Транспозиционные события приводят к мутациям, обусловливая формирование новых аллелей генов с последующим видообразованием. Каждое транспозиционное событие может увеличить геномный полиморфизм индивидуумов вида в ответ на изменения окружающих условий, после чего наступает период относительного покоя, приводящий к исчезновению большой части мобильных элементов генома. Так, у риса более 190 Мб LTR-ретротранспозонов утрачены около 8 млн лет назад (30). По другим оценкам, это произошло в более поздний период (19). В обоих случаях использованы данные полногеномного секвенирования O. sativa сорта Nipponbare.
Мобильные элементы вовлечены в доместикацию риса и процессы его адаптации. По одной из версий, шел негативный отбор против TEs HUO, который обнаруживается среди видов рода Oryza с AA геномом и в половине археологических семян риса, редуцирован в ландрасах, отсутствует в 115 современных сортах, идентифицирован только в одном китайском сорте. Увеличение числа копий HUO способно запускать геномную нестабильность посредством обширного метилирования ДНК и биогенеза малых РНК, глобально изменяя экспрессию генов. В результате снижается устойчивость к болезням и урожайность. Следствием этого, вероятно, стала элиминация TEs HUO в природе и в процессе селекции (31).
Таким образом, для природной эволюции необходима активация мобильных элементов, а для селекции — стабильность генома и транспози- ционное спокойствие в культивируемых сортах.
Исследования культивируемого риса O. sativa. Мобильные элементы O. sativa изучали в комплексных исследованиях динамики генома риса.
Используя данные полногеномного секвенирования, S.Y. Jiang с со-авт. (12) провели биоинформатический поиск генов, вовлекаемых в транспозиции мобильными элементами. У риса было идентифицировано 7043 LTR-ретротранспозона. Ими перенесено 672 гена, из которых только 36 % экспрессируются. У риса высокий процент псевдогенов (отсутствие интронов, полиА-последовательность на 3´-конце), транспортируемых ERV.
Расположение мобильных элементов относительно генов имеет решающее значение для гипометилирования. Метилирование ДНК тесно связано с ростом, развитием, яровизацией и цветением растений. Мутантную линию osmet1 сорта Kitaake (subsp. japonica ), характеризующуюся пониженным уровнем метилирования сравнили с растениями, выращенными на питательной среде с ингибитором метилирования Aza-D. Мобильные элементы SINE у риса демонстрировали наивысший уровень метилирования среди ретротранспозонов, а также его зависимость от удаленного или близкого расположения к белок-кодирующим генам (32). Возможно, в селекционной практике целесообразно учитывать транспозиции, влияющие на проявление хозяйственно ценных признаков.
Рис возделывается при создании слоя воды, что ведет к повышению концентрации ионов двухвалентного железа (Fe2+), токсичного для растений культурного вида. В проростках риса сорта Nipponbare в ответ на избыток железа активируются гены, вовлекаемые в гомеостаз по железу, и множество мобильных элементов участвуют в транскрипционном ответе на стресс и могут давать адаптивное преимущество растениям (9).
Хлорофилл-дефектная мутантная линия риса GR-7895 (subsp. japon-ica ) содержит транспозон nDart1-0 (pyl-v). Традиционным методом (бек-кроссированием) транспозон был перенесен в сорт риса Basmati-370 для описания гена, ответственного за GTP-связывающий белок, и его воздействия на активацию фитогормонов, влияющих на толерантность риса к биотическим и абиотическим стрессорам. Количество салициловой и гиббереллиновой кислот, антиоксидантов и малонового диальдегида увеличилось, в то время как содержание цитокининов, аскорбатпероксидазы, каталазы и флавоноидов сократилось. Таким образом, GTP-связывающий белок влияет на процессы, лежащие в основе формирования хлоропластов. Тем не менее исследователи пришли к заключению, что в условиях стресса мутанты с транспозоном nDart1-0 могут иметь преимущество, поэтому его необходимо переносить в другие сельскохозяйственные растения, например кукурузу, для изучения регуляции фотосинтеза у разных культур (33).
Полиморфизм LTR-ретротранспозона Tos17 изучен с помощью IRAP маркеров (три праймера) в двух сортах риса Osmancik-97 и Mevlutbey. Второй сорт характеризуется повышенной устойчивостью к экстремальным условиям среды. Загрязнение никелем и бором вызвало различную реакцию сортов: геномные изменения оказались более существенными у растений сорта Osmancik-97. Мобильность Tos17 может служить молекулярным маркером стрессовой толерантности O. sativa (34).
Предпринята попытка направленного мутагенеза по 17 генам риса, ответственных за стрессоустойчивость. В промоторах пяти генов (29 % таргетированных) удалось получить инсерции мобильного элемента mPing, вызвавшие изменения в генной экспрессии в условиях холодового и солевого стрессов (35). Ключевая роль в условиях дефицита влаги у риса принадлежит 62
гену DRO1 . Индуцированные условия засухи привели к транспозиции IN-DITTO2 (MITE), которая действует как автономный ауксин-отзывчивый промотор, усиливающий физиологическую адаптацию при дефиците влаги (36). Следовательно, мобильные элементы вовлекаются в гормональную регуляцию процессов адаптации.
МикроРНК — часть некодирующей РНК, тесно связанная по своему происхождению с транспозонами (1), — участвует в РНК-сайленсинге и посттранскрипционной регуляции. Длинные некодирующие РНК (lncRNAs) регулируют генную экспрессию и другие процессы, кольцевая circRNAs ковалентно связывает концы и вовлекается в транскрипционную и посттранскрипционную регуляцию, влияя, в конечном счете, на продукцию белка. У риса subsp. indica ∼ 53 % lncRNAs и ∼ 15 % circRNAs ассоциировано с мобильными элементами, в особенности с MITE. Высокая степень метилирования ДНК мобильных элементов затрагивает экспрессию lncRNAs и circRNAs. (37). Число копий мобильных элементов играет решающую роль в проявлении агрономически важных признаков. При сравнении двух почти изогенных линий риса с одной и тремя копиями HUO доказано снижение продуктивности метелки (число и масса семян) и устойчивости к бактериальному ожогу ( Xanthomonas oryzae ) у трехкопийной линии. Однако устойчивость к хлоридному засолению у этой линии была выше (31).
Практические результаты получены при облучении семян риса сорта Jijing 809 (subsp. japonica ) пучками тяжелых ионов высокой энергии. Созданы три мутантные формы с транспозициями Dasheng, mPing, Osr13 и RIRE2, расположенными рядом с некоторыми генами, например OsSPCH2 и OsCPS2 . В результате возросли размеры и масса семян, сократился период до цветения растений (38). Воздействие на некоторые гены не приводило к фенотипическим изменениям или отрицательно сказывалось на агрономически важных признаках (13, 38). Повреждения ДНК физическим мутагеном вызывают активацию мобильных элементов, приводящую к существенным однонуклеотидным заменам (SNPs) и инсерциям/делециям (InDels) (38).
Сельскохозяйственное производство в значительной степени подвержено воздействию меняющихся погодных условий, в силу чего адаптация культурных растений к климатическим изменениям становится сложной селекционной задачей (39). Стрессоустойчивость сельскохозяйственных растений — ключевая проблема при получении урожаев. Только пластичные сорта, способные реализовать свой потенциал вне зависимости от колебаний условий окружающей среды, представляют интерес для товаропроизводителя. Поскольку активация транспозиционных событий происходит под действием стресса, это способ расширения нормы реакции генотипов и изменения биохимических процессов у растений.
Рис используют в космических исследовательских программах (40). Искусственное воздействие тяжелых ионов высокой энергии на транспозиционную активность O. sativa доказано на Земле (38). В условиях космического полета происходит естественная обработка семян таким физическим фактором, снижающим степень метилирования мобильных элементов группы LINE. Степень метилирования SINE, LTR, RC (rolling-circle) зависит от особенностей механизма метилирования (CpG, CHH или CHG). В любом случае результат наследуется в ряду поколений мутантов до M 5 (40). Результаты исследований доказывают неизбежность эволюционных преобразований живых организмов в космосе, и этот фактор следует учитывать при разработке программ освоения дальнего космоса.
Исследования в культуре тканей in vitro . Культуры in vitro рассматриваются как фактор стрессового воздействия наряду с другими биотическими и абиотическими стрессами для растений, такими как жара, холод, засуха, солевой стресс, воздействие патогенов, повреждение, радиация, полиплоидизационные и гибридизационные события (7). В культурах in vitro возникает геномная нестабильность, которая может быть связана с активацией мобильных элементов с последующими мутационными изменениями, что наблюдается у ряда культур, в том числе у O. sativa (41).
Из многообразия мобильных элементов у риса по крайней мере 13 TEs демонстрируют транскрипционную активность in planta в каллусной культуре мутантной линии AB156365, полученной из сорта Nipponbare. Из них LTR-ретротранспозоны — Tos17, osr37, osr10, BAJIE, RIRE2, RIRE3, RN363, RN216; MITE мобильные элементы — mPing, Mite#1, Mite#2, Mite#3, Tami2 (42). N. Picault с соавт (20), исследуя каллусную культуру немутантных растений этого же сорта, дополнили список LTR-ретротранспозоном Lullaby. Показано отсутствие корреляции между экспрессией и транспозиционной активностью, в частности у Tos17, что предполагает ретротранспозиции в каллусе риса независимо от транскрипционного и посттранскрипционного контроля (42). Наличие транскриптов считается плохим предиктором транспозиционного потенциала вследствие посттранскрипционного контроля (43).
Активация LTR-ретротранспозонов в каллусе риса сортоспецифична: Tos17 активен в референсном геноме сорта Nipponbare, интактен в сортах Moritawase и Dongjin (subsp. japonica ), мобильный элемент семейства Oryco, наоборот, показывает высокий транспозиционный потенциал в корейском сорте Dongjin, но интактен в сорте Nipponbare (44). Также выявлена подвидовая специфичность LTR-ретротранспозонов суперсемейства Copia. У трансформированных растений риса сорта IR64 (subsp. indica ) мобильный элемент Go-on активируется высокими температурами в период роста каллуса, чего не наблюдается у растений сорта Nipponbare (subsp. japonica ) (43). Существуют универсальные мобильные элементы, проявляющие в стрессовых условиях активность у современных видов рода Oryza , в ландрасах, сортах и культуре тканей in vitro O. sativa . К таким относится близкородственный Tos17 и Lullaby мобильный элемент HUO, отсутствующий в референсных геномах сортов Nipponbare и 93-10 (31).
Транспозиция Tos17 активируется в культуре ткани растений сорта Nipponbare и не проявляется в регенерированных растениях. Для растений-регенерантов характерно присутствие 5-30 копий Tos17, тогда как в культуре ткани число копий коррелирует с продолжительностью периода культивирования. В референсном геноме риса выявлено две почти идентичные копии Tos17 на 7-й и 10-й хромосомах. Только одна из них, расположенная на 7-й хромосоме, транспозиционно активна, по крайней мере в культуре in vitro (22). Неактивная копия на 10-й хромосоме сильно метилирована, содержит несколько стоп-кодонов и InDels в кодирующей области (21).
Регуляция мобильного элемента Karma (LINE) прослежена на 54 клеточных линиях, которые культивировали от 5 мес до 4 лет, а также на регенерантах из 18 независимых трансгенных каллусных линий риса. Обнаружено отсутствие ретротранспозиций Karma в культурах клеток и растениях R0. Увеличение числа копий мобильных элементов начинается с первого поколения и продолжается до шестого поколения регенерантов. Все же Karma-транскрипты активируются в культуре тканей и ретротранспозиции имеют продолжение в растениях-регенерантах. В отличие от других ретротранспозонов, у Karma транскрипция после активации не прекращается и гипометилирование (эпигенетическая регуляция у транспозонов) идет в нескольких поколениях регенерантов (5).
Используя in vitro культуру тканей, у сортов Ilmibyeo и Baegjinju1ho (subsp. japonica ) удалось получить соответственно 1672 и 843 мутантные формы. Проанализировано 1533 гена на наличие инсерций Tos17, мутации выявлены в 830 из них (45). Однонуклеотидные замены (SNPs), инделы и Tos17 транспозиции — главные причины сомаклональной изменчивости in vitro (46). Индуцированный мутагенез вызвал минимум в десяти растениях положительные агрономически важные изменения: существенно увеличились размер и масса зерна (прибавка достигала 56 %) (45).
У растений риса были изучены TEs, ассоциированные с «открытым» хроматином (ТEs-derived accessible chromatin regions, ACRs). Использовались культуры тканей от шести типов эксплантов: молодой лист, флаговый лист, корень, тычинки и пестики, верхняя и нижняя чешуя цветка, молодая метелка (15). Оказалось, что мобильные элементы и TEs-ACRs, в том числе тканеспецифичные, больше распространены в дистальных межгенных областях или за генами. На TEs-ACRs приходилось 8,6 % ACRs, из них тканеспецифичных было 3,5 %. В составе TEs-ACRs хроматиновые области демонстрировали различные уровни доступности и консервативности по сравнению с ACRs без TEs. Некоторые тканеспецифичные TEs-ACRs расположены выше группы функциональных тканеспецифичных генов. Например, хроматиновый регион, несущий MULE, локализовался в промоторе гена LOC_Os03g44180 гидрохинон-глюкозилтрансферазы, в основном экспрессирующегося в корнях. Таким образом, TEs-ACRs могут играть важную роль в регуляции экспрессии тканеспецифичных генов, тем самым вовлекаясь в развитие тканей у риса (15).
У O. sativa известно большое число генов, ответственных за хозяйственно полезные признаки. Создавая стрессовые условия in vitro для растений, удается получить мутантные формы с перемещением мобильных элементов. Индуцированные транспозиции у риса наследуются (31). При этом во многих случаях затронуты агрономически важные характеристики (15, 31, 38). Направленный мутагенез растений с вставками мобильных элементов в целевых генах при молекулярно-генетическом контроле с последующим отбором — это новое окно возможностей для пребридинга в селекции O. sativa .
Предлагаются протоколы геномного редактирования генов, захваченных мобильными элементами. Так, с помощью технологии CRISPR/Cas9 в культуре in vitro сорта Nipponbare получен регенерант риса с целевой мутацией, вызвавшей делецию ретротранспозона Tos17 на 7-й хромосоме. Предполагается, что в обозримой перспективе станет возможным редактировать хозяйственно ценные гены, поврежденные TEs (47). Рекультивирование in vitro семян отредактированной линии показало сохранность изменений на 7-й хромосоме, что свидетельствует о стабильности и наследуемости инактивированной делеции (48).
Исследования в андрогенезе in vitro. Гаплоидные технологии представляют часть методов культуры клеток (андрогенез in vitro) и направлены на создание гаплоидных растений или гомозиготных удвоенных гаплоидов (49, 50). У большинства видов для этой цели используется культура микроспор, из которых напрямую регенерируют растения (50). Регенеранты риса получают из пыльников через каллусообразование (51, 52), в процессе которого возможны мутации разного типа (51).
Известно, что в андрогенных каллусах риса сорта Nipponbare происходит активация мобильных элементов семейства mPing (MITE), которые могут быть одним из источников появления мутантов (53). Возникновение мутаций, связанных с Tos17, описано в каллусах риса, полученных из соматических клеток (22). В регенерантах риса андрогенного происхождения из сорта Jiahua No.1 (subsp. japonica) продемонстрированы активные транспозиции еще одного MITE мобильного элемента — mGing (54). H. Yang с соавт. (55) предполагают, что в подобных исследованиях транспозиционные события могли случиться в соматических клетках in vitro, из которых регенерировали гетерозиготные мутантные растения. Поскольку в андрогенезе in vitro возникновение гетерозиготных растений возможно разными путями (50, 56), для актуализации выводов о TEs активности необходима доказательная база происхождения регенерантов из клетки микроспоры (55).
На сорте риса Nipponbare проведено сравнительное изучение активности видоспецифичного транспозона nDaiZ (семейство hAT) на интактных растениях и регенерантах R 0 -R 1 , полученных в культуре тканей и андроге-незе in vitro (57). Активация мобильного элемента происходила исключительно в условиях in vitro. Частота транспозиций была выше в культуре тканей и составляла 29,31 %, в андрогенезе in vitro — 8,60 %. В обычных условиях происходил эпигенетический сайленсинг метилированием ДНК, в процессе культивирования тканей in vitro наблюдалось деметилирование (57).
Несмотря на то, что гаплоидные технологии используются для быстрого перевода гибридов в гомозиготное состояние и ускорения селекционного процесса (50, 52), предлагается индуцируемый транспозиционный мутагенез. В андрогенезе in vitro на 100 регенерантов удалось получить шесть гомозиготных мутантов с синтетическим транспозоном COKC, правда, без фенотипических проявлений (55). Такие мутанты могут использоваться в селекционной работе в качестве исходного материала при должной полевой оценке.
Широкомасштабные исследования мобильных элементов в андроге-незе in vitro O. sativa в научной литературе нам не встречались. Между тем предсказать их поведение отчасти возможно. Известно, что гибридизационные события увеличивают гетерозиготность и ведут к активации TEs (58). Следовательно, гомозиготные удвоенные гаплоиды, вероятно, будут характеризоваться меньшей активностью TEs. Транспозоны андрогенных регенерантов менее активны даже в сравнении с культурой соматических тканей in vitro (57). Гаплоидные растения, будучи носителями одинарного набора хромосом, также не могут позволить себе активных перемещений мобильных элементов в геноме. В противном случае последствия для гаплоидных индивидуумов будут губительными. В андрогенезе in vitro отмечается большая доля (около 15-20 %) нежизнеспособных зеленых регенерантов, погибающих на стадии укоренения в условиях ex vitro или несколько позже (59). Считается, что они несут летальные и полулетальные мутации (60), вполне возможно вызванные активацией TEs. Большая доля альбинизма в андро-генезе in vitro (от 5 до 100 % регенерантов) ограничивает общий выход продуктивных растений (51, 52). Природа мутаций нежизнеспособных и хло-рофилл-дефектных растений глубоко не изучалась. Именно такие растения в первую очередь могут стать источником информации о поведении мобильных элементов у гаплоидных генотипов. Поиск активных мобильных элементов затруднен и трудно прогнозируем. Обнаруженные у риса транспозиции выявлены на диплоидных растениях (20). Малочисленность исследований TEs O. sativa в андрогенезе in vitro в эру биотехнологий и наличие полностью секвенированного генома вида скорее свидетельствуют о транспозиционном спокойствии гаплоидов и удвоенных гаплоидов риса.
Итак, на заре генетики были сформированы представления о спонтанном и индуцированном мутагенезе. Индуцированные мутации можно 66
получить, воздействуя ограниченным набором физических или химических мутагенов, что успешно использовалось в селекционной практике. Позднее список мутагенов значительно расширился. В настоящее время очевидно, что любой биотический или абиотический стрессовый фактор может вызвать активацию мобильных элементов, что ведет к изменению генотипа. Для пребридинга культуры необходимо большое разнообразие растений, из которых в конечном счете будет отобран сорт. Это разнообразие можно получить за счет транспозиций. Исследования Oryza sativa продемонстрировали, что мутации, вызванные мобильными элементами, часто положительные. Селекция неизбежно воспользуется возможностями мобильных элементов, которые участвуют в перестройке генома растений с агрономически позитивными отклонениями, наследуемыми в поколениях. Проблема состоит лишь в том, чтобы научиться получать направленные изменения в фенотипе риса, используя транспозиции, для повышения продуктивности растений и качества зерна. Сейчас появились возможности для обратной коррекции генов, поврежденных мобильными элементами, с помощью геномного редактирования. Стал возможным молекулярно-генетический контроль стрессовой толерантности O. sativa на основе мобильного элемента Tos17 . В целом, знания о мобильных генетических элементах и вызываемых ими изменениях уже начали приносить первые результаты, связанные с повышением продуктивности модельного объекта O. sativa , и могут стать основой для изучения мобилома множества сельскохозяйственных культур.
Автор выражает глубокую признательность академику РАН Ю.Н. Журавлеву за идею обобщения информации о мобильных элементах риса.
ФРБНУ ФНЦ агробиотехнологий Дальнего Бостока им. А.К. Чайки,
