Модель ингибирования автогенной сукцессии восстановления растительности на месте заброшенных деревень в горно-лесной зоне Южного Урала

Автор: Миркин Б.М., Сайфуллина Н.М., Ямалов С.М.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Лазерная техника и технология
Статья в выпуске: 2 т.8, 2006 года.
Бесплатный доступ
В статье рассматривается восстановительная сукцессия растительности на месте заброшенных деревень, как пример модели ингибирования. Ингибирование осуществляется нитрофильными рудеральными видами класса Galio-Utricetea Passarge 1967. Приводятся результаты дисперсионного анализа диагностических видов пяти классов растительности. Показана сила влияния факторов “сукцессионное время” и “характер использования”, а также генеральные средние распределения видов по классам этих градиентов.
Короткий адрес: https://sciup.org/148197831
IDR: 148197831
Текст научной статьи Модель ингибирования автогенной сукцессии восстановления растительности на месте заброшенных деревень в горно-лесной зоне Южного Урала
-
1 Башкирский государственный университет, г. Уфа
-
2 Государственный природный заповедник “Шульган-Таш”, с. Иргизлы
В статье рассматривается восстановительная сукцессия растительности на месте заброшенных деревень, как пример модели ингибирования. Ингибирование осуществляется нитрофильными рудеральными видами класса Galio-Utricetea Passarge 1967. Приводятся результаты дисперсионного анализа диагностических видов пяти классов растительности. Показана сила влияния факторов “сукцессионное время” и “характер использования”, а также генеральные средние распределения видов по классам этих градиентов.
В 70-80-е годы прошлого столетия намечался “динамический бум” в фитоценологии, что отражало “… социальный заказ в связи с резко усилившимся влиянием человека на растительность – понять суть происходящих изменений, чтобы прогнозировать их и поставить под контроль. Кроме того, изучая динамику растительности, исследователь получал ценнейшую информацию о динамике экосис-тем…Таким образом, “динамический бум” в фитоценологии был отражением общего “динамического бума” в экологии” [4].
Одним из главных достижений “динамического бума” было развитие представлений Ф. Клементса об автогенных экологических сукцессиях. Они стали рассматриваться как стохастический процесс, соответствующий не одной модели (повышение продуктивности, биоразнообразия, степени развития почв и т.д. – т.е. развития в направлении моноклимакса), а трем моделям – благоприятствования (“модель Клементса”), толерантности (ухудшение условий среды со сменой видов по градиенту повышения патиентности) и ингибирования (остановки сукцессии на стадии серийных сообществ вследствие её блокирования одним видом или их группой) [10].
Модель ингибирования иллюстрировалась на примере сукцессий, которые блокировали мхи – кукушкин лен на пожарищах [12] и зеленые мхи и водоросли, которые формируют пленку карахарсанг [2], “запирающую” пустынные грунты от поселения сосудистых растений.
В дальнейшем было показано, что блокирующими видами могут быть рудералы: однолетние костры препятствовали восстановлению североамериканских прерий [3], что стало стимулом для разработки Дж. Кертисом метода восстановления прерий с высевом смеси семян, собранных в естественной прерии (затем этот метод усовершенствовал Д. Дзыбов в своих “агростепях”, когда высевались не семена, на сбор которых затрачивается много времени, а скошенное и размельченное сено). О блокировании сукцессий рудералами писали А.А. Тишков [6; 7] и Е.А. Шварц [8].
Авторами описана автогенная восстановительная сукцессия, соответствующая модели ингибирования на Южном Урале, при восстановлении растительности на месте заброшенных деревень. В горно-лесной зоне Республики Башкортостан в период с 1918 по 1988 гг. было заброшено множество мелких населенных пунктов. Причинами этого были голод, разруха послевоенных лет, и, в особенности, программа ликвидации “неперспективных деревень”, сократившая число населенных пунктов на 43% [1; 9]. Большая часть постселитебных территорий, размер которых был невелик (0,04-2 км2), в дальнейшем использовалась для выпаса и сенокошения. Меньшая их часть, расположенная в зоне заповедования государствен- ного природного заповедника “Шульган-Таш” и национального парка “Башкирия”, восстанавливалась в режиме залежи.
Для изучения этого процесса было выполнено 602 геоботанических описания сообществ с сукцессионным возрастом от 12 до 92 лет. Материал был обработан в соответствии с принципами системы Браун-Бланке, причем, поскольку большинство сообществ представляло “фитосоциологические смеси” видов разных классов, то использовался не классический синтаксономический анализ, а так называемый дедуктивный метод классификации растительности [11]. На основании этого метода выделяются базальные и (или) дериватные сообщества, которые подчиняются непосредственно классу, порядку или союзу на основе представленности диагностических видов высших единиц [5].
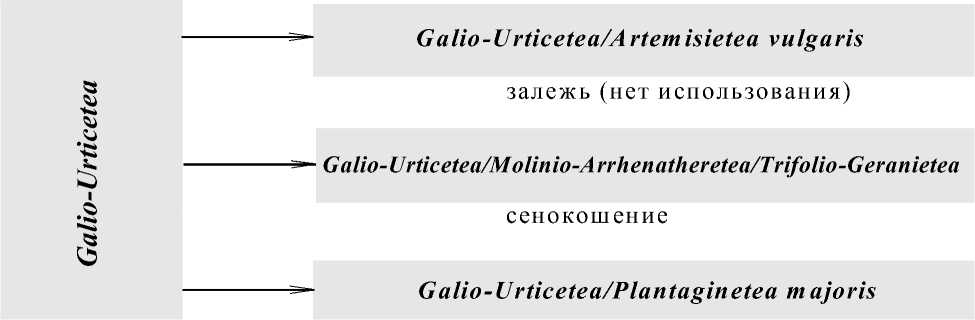
Характеристике всех типов выделенных сообществ будет посвящена специальная статья. В этом сообщении мы ограничимся приведением общей сукцессионной схемы на уровне классов системы Браун-Бланке и обсуждением результатов градиентного (дисперсионного) анализа. Анализ был выполнен по факторам “сукцессионное время” (три класса: I – 12-30 лет; II – 30-45 лет; III – свыше 45 лет) и характер использования территории заброшенных деревень (три градации: залежь, сенокос, пастбище). В последние годы процесс забрасывания деревень прекратился, и потому в распоряжении авторов не было материала о первых годах сукцессии, когда процессы восстановления растительности протекают особенно интенсивно. Классовый интервал первой стадии сукцессии в 12-30 лет слишком широк, что не позволяет описать первые этапы восстановления растительности. По этой причине на схеме сукцессии характер начальной стадии указан гипотетически (рис.).
Из рисунка очевидно, что при полном отсутствии использования, когда из экосистемы заброшенного поселения практически не происходит оттока веществ, эффект ингибирования проявляется наиболее полно, и за 50 лет проанализированной сукцессии практически сохраняются рудеральные сообщества класса Galio-Utricetea Passarge 1967, характерные для богатых и хорошо увлажненных местообитаний. В их составе появляются также рудеральные виды Artemisietea vulgaris Lohmeyer et al. in R. Tx. 1950, которые представляют сообщества дву-многолет-них рудералов. При пастбищном использовании происходит смена сообществ более сложными, в состав которых, кроме сохраняющихся видов класса Galio-Utricetea, входят виды сообществ луговых пастбищ – класс Plantaginetea majoris R. Tx. et Preising in R. Tx. 1950. И, наконец, при сенокосном использовании формируются наиболее богатые видами сообщества, в состав которых входят, наряду с рудеральными видами класса Galio-Utricetea , луговые виды класса Molinio-Arrhenatheretea R. Tx. 1937 em. R. Tx. 1970 и виды сообществ ксеротермных опушек класса Trifolio-Geranietea T. Mьller 1961.
В таблице показаны результаты дисперсионного анализа диагностических видов
выпас
Рис. Схема динамики растительности в процессе восстановительных сукцессий на месте заброшенных деревень при разном режиме использования.
Таблица. Влияние факторов использования и стадии сукцессии на постоянство диагностических видов основных классов
|
Виды |
Об щее чис ло встр еч |
Сила влияния фактора |
Генеральные средние |
|||||||
|
A |
B |
AB |
A1 |
A2 |
A3 |
B1 |
B2 |
B3 |
||
|
Диагностические виды класса Molinio-Arrhenatheretea |
||||||||||
|
Ranunculus polyanthemos |
87 |
0,23 |
0,00* |
0,03 |
0,65 |
0,06 |
0,21 |
0,10 |
0,26 |
0,27 |
|
Alopecurus pratensis |
21 |
0,08 |
0,02 |
0,07 |
0,20 |
0,02 |
0,03 |
0,02 |
0,08 |
0,05 |
|
Festuca pratensis |
89 |
0,08 |
0,00* |
0,01* |
0,48 |
0,12 |
0,23 |
0,17 |
0,24 |
0,23 |
|
Lathyrus pratensis |
110 |
0,12 |
0,01* |
0,05 |
0,58 |
0,14 |
0,29 |
0,11 |
0,40 |
0,29 |
|
Phleum pratense |
129 |
0,12 |
0,00* |
0,02 |
0,62 |
0,18 |
0,35 |
0,29 |
0,34 |
0,30 |
|
Poa trivialis |
61 |
0,05 |
0,00* |
0,02 |
0,31 |
0,11 |
0,13 |
0,09 |
0,19 |
0,17 |
|
Vicia cracca |
62 |
0,14 |
0,00* |
0,01* |
0,46 |
0,06 |
0,13 |
0,09 |
0,20 |
0,16 |
|
Pimpinella saxifraga |
108 |
0,12 |
0,02 |
0,03 |
0,18 |
0,13 |
0,45 |
0,34 |
0,26 |
0,17 |
|
Cerastium holosteoides |
50 |
0,09 |
0,01 |
0,04 |
0,08 |
0,02 |
0,27 |
0,10 |
0,13 |
0,13 |
|
Prunella vulgaris |
64 |
0,13 |
0,02 |
0,00* |
0,09 |
0,04 |
0,32 |
0,12 |
0,19 |
0,15 |
|
Ranunculus acris |
66 |
0,10 |
0,04 |
0,04 |
0,32 |
0,04 |
0,25 |
0,11 |
0,26 |
0,10 |
|
Achillea millefolium |
264 |
0,24 |
0,00* |
0,00* |
0,92 |
0,39 |
0,83 |
0,55 |
0,66 |
0,70 |
|
Geranium pratense |
133 |
0,02 |
0,01 |
0,03 |
0,38 |
0,24 |
0,40 |
0,31 |
0,37 |
0,28 |
|
Stellaria graminea |
182 |
0,24 |
0,00* |
0,01* |
0,82 |
0,19 |
0,60 |
0,29 |
0,49 |
0,54 |
|
Trifolium pratense |
119 |
0,11 |
0,00* |
0,06 |
0,37 |
0,12 |
0,47 |
0,22 |
0,29 |
0,36 |
|
Dactylis glomerata |
293 |
0,02 |
0,02 |
0,01* |
0,82 |
0,70 |
0,67 |
0,69 |
0,80 |
0,62 |
|
Диагностические виды кла сса Trifolio-G eranietea |
||||||||||
|
Veronica teucrium |
114 |
0,19 |
0,02 |
0,02 |
0,68 |
0,11 |
0,31 |
0,17 |
0,40 |
0,25 |
|
Fragaria viridis |
221 |
0,19 |
0,00* |
0,01* |
0,91 |
0,32 |
0,64 |
0,42 |
0,56 |
0,62 |
|
Agrimonia asiatica |
122 |
0,10 |
0,00* |
0,02 |
0,14 |
0,20 |
0,47 |
0,33 |
0,27 |
0,28 |
|
Hypericum perforatum |
73 |
0,06 |
0,01 |
0,04 |
0,29 |
0,06 |
0,28 |
0,11 |
0,24 |
0,17 |
|
Origanum vulgare |
140 |
0,05 |
0,02 |
0,03 |
0,49 |
0,21 |
0,44 |
0,29 |
0,42 |
0,30 |
|
Trifolium medium |
119 |
0,08 |
0,09 |
0,02 |
0,46 |
0,16 |
0,38 |
0,23 |
0,47 |
0,15 |
|
Диагностические виды класса Artemisietea vulgaris |
||||||||||
|
Artemisia absinthium |
187 |
0,15 |
0,08 |
0,01* |
0,25 |
0,31 |
0,71 |
0,39 |
0,30 |
0,67 |
|
Carduus acanthoides |
180 |
0,23 |
0,05 |
0,03 |
0,08 |
0,33 |
0,71 |
0,48 |
0,22 |
0,61 |
|
Arctium tomentosum |
29 |
0,02 |
0,06 |
0,03 |
0,12 |
0,02 |
0,12 |
0,03 |
0,03 |
0,16 |
|
Artemisia vulgaris |
102 |
0,03 |
0,01* |
0,05 |
0,35 |
0,27 |
0,17 |
0,20 |
0,32 |
0,22 |
|
Leonurus quinquelobatus |
284 |
0,15 |
0,02 |
0,01* |
0,29 |
0,81 |
0,69 |
0,72 |
0,59 |
0,74 |
|
Carduus crispus |
21 |
0,01* |
0,01* |
0,02* |
0,03 |
0,08 |
0,03 |
0,09 |
0,01 |
0,05 |
|
Диагностические виды класса Galio-Urticetea |
||||||||||
|
Galium aparine |
135 |
0,09 |
0,01* |
0,01* |
0,14 |
0,47 |
0,22 |
0,34 |
0,31 |
0,33 |
|
Urtica dioica |
299 |
0,27 |
0,00* |
0,01* |
0,28 |
0,93 |
0,64 |
0,82 |
0,65 |
0,69 |
|
Geum urbanum |
169 |
0,09 |
0,05 |
0,02* |
0,25 |
0,33 |
0,58 |
0,25 |
0,49 |
0,48 |
|
Glechoma hederacea |
193 |
0,08 |
0,00* |
0,01* |
0,25 |
0,41 |
0,63 |
0,47 |
0,45 |
0,48 |
|
Chaerophyllum prescottii |
206 |
0,15 |
0,00* |
0,01* |
0,83 |
0,55 |
0,29 |
0,42 |
0,59 |
0,48 |
|
Диагностические виды класса Plantaginetea majoris |
||||||||||
|
Plantago major |
86 |
0,29 |
0,01* |
0,01* |
0,06 |
0,49 |
0,03 |
0,23 |
0,13 |
0,27 |
|
Taraxacum officinale |
170 |
0,24 |
0,01 |
0,07 |
0,57 |
0,69 |
0,14 |
0,31 |
0,41 |
0,52 |
|
Amoria repens |
112 |
0,18 |
0,03 |
0,03 |
0,29 |
0,53 |
0,06 |
0,26 |
0,21 |
0,34 |
Примечание: фактор А - характер использования (А1- сенокос, А2 - пастбище, А3 - залежь);
фактор В - сукцессионный возраст сообществ (В1 - 12-30 лет; В2 - 30-45 лет; В3 - свыше 45 лет);
* - влияние фактора недостоверно.
пяти перечисленных классов растительности. Показана сила влияния факторов “сукцессионное время” и “характер использования”, а также генеральные средние распределения видов по классам этих градиентов.
Из таблицы очевидно, что виды, входящие в диагностическую комбинацию разных классов существенно различаются по отношению к характеру использования и фактору сукцессионного времени. Обращает на себя внимание то, что для всех видов характер использования является более значимым, чем сукцессионное время, распределение по градиенту которого ингибируется рудеральными видами.
Нетрудно видеть, что виды вторичных послелесных лугов класса Molinio-Arrhenatheretea в основном тяготеют к сенокосному использованию и 2-3 стадиям сукцессии. Некоторые виды этого класса ( Cerastium holosteoides, Pimpinella saxifrage, Prunella vulgaris, Trifolium pretense) повышают постоянство при залежном варианте. Сходную тенденцию проявляют и виды класса ксеротермных опушек Trifolio-Geranietea. Эту закономерность нетрудно объяснить полу-естественной природой растительности данных классов.
Виды класса Artemisietea vulgaris показывают четкое тяготение к залежному варианту, причем их постоянство в ходе сукцессии возрастает. Главные “ингибиторы” сукцессии – виды класса Galio-Utricetea представлены во всех вариантах сукцессии и во всех возрастных стадиях, значения силы влияния факторов для них минимальны. И, наконец, пастбищные виды класса Plantaginetea majoris встречаются во всех возрастных стадиях, причем, естественно, что их максимальное постоянство отмечается в пастбищном варианте.
Весьма характерно, что даже при двух вариантах сукцессии, которые протекают на фоне использования (что способствует выносу элементов минерального питания из почвы) сохраняется роль рудеральных видов. Кроме того, сукцессия не идет в направлении формирования сообществ типичных лугов горно-лесной зоны – порядка Carici macrourae-Сrepidetalia sibiricae Ermakov et al. 1999, которые могут сменяться лесами. Таким образом, ингибирующий эффект видов ценных стадий сукцессии наблюдается и в этих сериях.
С чем связано ингибирующее влияние эксплерентов из класса Galio-Utricetea? Авторы полагают, что причиной замедленного характера сукцессий является высокий трофический статус почв на заброшенных территориях поселений, что создает условия эко-лого-фитоценотического оптимума для этих видов и усиливает их конкурентную способность. Это позволяет рудералам прочно удерживать свои фитоценотические позиции и ингибировать процесс сукцессии.
Работа поддержана грантом РФФИ (проект № 04-04-49296а).
Список литературы Модель ингибирования автогенной сукцессии восстановления растительности на месте заброшенных деревень в горно-лесной зоне Южного Урала
- Acфандияров А.З. История сел и деревень Башкирской АССР. Кн. 1. Уфа: Башкирское книжное издательство, 1990.
- Нечаева Н.Т., Мухаммедов Г.М. Мониторинг природной и улучшенной растительности центральных Каракумов. Ашхабад, 1991.
- Миркин Б.М., Наумова Л.Г. Адвентивизация растительности: инвазивные виды и инвазибельность сообществ//Успехи современной биологии. 2001. Т.121. №6.
- Миркин Б. М. Наумова Л. Г. Наука о растительности (история и современное состояние основных концепций). Уфа: Гилем, 1998.
- Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности. М.: Логос, 2000.
- Тишков А.А. Динамика заповедных экосистем и проблемы сохранения ценофонда//Заповедники СССР: их настоящее и будущее. Ч 1. М., 1990.
- Тишков А.А., Масляков В.Ю., Царевская Н.Г. Антропогенная трансформация биоразнообразия в процессе непреднамеренной интродукции организмов (биогеографические последствия)//Изв. РАН. Сер. географич. 1995. №4.
