Моделирование среды, как экологический способ решения актуальных проблем аквакультуры

Автор: Пономарева Елена Николаевна, Металлов Геннадий Федорович, Левина Ольга Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1-1 т.16, 2014 года.
Бесплатный доступ
В статье приводятся результаты исследований по улучшению качества рыбоводной продукции при выращивании молоди осетровых в установках замкнутого водоснабжения. Анализируется эффективность применение солевой среды, как способа решения проблем пастбищной и товарной аквакультуры. Оцениваются рыбоводно-биологические и физиолого-биохимические параметры молоди русского осетра в соленой и пресной воде.
Аквакультура, русский осетр, установка замкнутого водоснабжения, рыбоводные показатели, физиология, соленость
Короткий адрес: https://sciup.org/148202699
IDR: 148202699 | УДК: 639.371.2.07.053.1:556.531.4.001.57
Environment modelling, as ecological way of the decision actual problems of the aquaculture
Results of researches on improvement of quality of fish-breeding production are given in article at cultivation sturgeon’s fry in recircular system. Efficiency application of the salt environment, as way of the solution of problems of a pasturable and market aquaculture is analyzed. Piscicultural, physiological and biochemical parameters are estimated for the Russian sturgeon fry in saltwater and freshwater.
Текст научной статьи Моделирование среды, как экологический способ решения актуальных проблем аквакультуры
Одним из важнейших рыбохозяйственных водоемов России до недавнего времени, отличающегося высокой биологической продуктивностью и уникальным видовым составом ихтиофауны являлось Каспийское море. Особый статус, среди водных биологических ресурсов этого водоёма принадлежит осетровым рыбам [1]. Однако в связи с ощутимым антропогенным воздействием на водные экосистемы крайне актуальной становится проблема сохранения биологического разнообразия рыб этого водоёма. Ежегодно снижаются запасы практически всех экологических групп рыб, в том числе и осетровых.
Между тем осетровые рыбы – интересны не только как предмет исследования эволюционных преобразований животного мира, но и как исключительно ценный объект промысла, объем вылова, которых значительно сократился за последние десятилетия. В результате сильного антропогенного воздействия (браконьерство на путях нерестовых миграций, нарушение гидрологического режима на нерестилищах и т.д.) естественное воспроизводство осетровых на основных реках Каспийского бассейна находится на критическом уровне. В условиях снижения эффективности естественного воспроизводства этих рыб ведущую роль в восстановлении их запасов в настоящее время играет искусственное воспроизводство, масштабы которого сокращаются [2].
Основными причинами сокращения искусственного воспроизводства являются: снижение уловов осетровых рыб, плохое качество производителей естественной генерации и низкая эффективность работы рыбоводных заводов за счёт устаревшей технологии. Для совершенствования
методов искусственного воспроизводства с целью сохранения и восстановления запасов этих рыб необходимо проведение комплексных физиологических и рыбоводно-биологических исследований.
В связи с этим важнейшим условием получения качественной рыбоводной продукции при выращивании молоди осетровых в установках замкнутого водоснабжения (УЗВ) явилось создание таких условий, при которых параметры искусственной среды соответствовали бы естественным. Это предполагало, выращивание молоди в 5‰ солевой среде, более эффективное регулирование других гидрохимических показателей, очистку от метаболитов и постоянный физиологический контроль за состоянием рыб. Выращивание молоди в среде максимально приближённой к естественным условиям позволяло ожидать более высокую выживаемость молоди осетровых рыб после её интродукции в Северный Каспий [3].
Многолетние исследования солеустойчивости осетра, белуги, севрюги на ранних этапах онтогенеза свидетельствуют об их широкой эвригалинности. Ранняя эвригалинность формируется за счет органов, участвующих в водно-солевом обмене (хлоридсекретирующие клетки жабр, почки, интерреналовая и щитовидные железы, гипотала-мо-гипофизарный комплекс) и расценивается, как адаптация, направленная на снижение смертности потомства. Изменение солености вызывает у молоди активные поведенческие реакции, наблюдается избирание оптимальных солевых зон, изменение в двигательной и пищевой активности [410]. Однако, подобные исследования ориентировались в первую очередь на изучение выживаемости молоди в различных градиентах солености. При этом не всегда учитывалась динамика осмоляльности крови, а так же изменения её основных биохимических показателей, характеризующих белковый, жировой и окислительный обмены.
Успех адаптации к солёности во многом зависит от способности рыб перестраивать свой водно-солевой обмен. У русского осетра в Южном Каспии осмоляльность сыворотки крови находится в пределах 264-368 мосм/кг Н 2 О. При миграции рыб из более солёного Южного Каспия в опреснённый Северный Каспий наблюдается постепенное снижение осмоляльности сыворотки крови. Если в более южных участках моря этот показатель незначительно превышает осмолярность морской воды или почти равен ей, то в Северном Каспии наблюдается гипертония, осмолярность сыворотки крови превышает её уровень в среде обитания и колеблется в пределах 219-322 мосм/кг Н 2 О [10].
Проведенные ранее исследования на молоди гибридных форм осетровых (русско-ленский осетр, стербел) так же свидетельствовали об их высоком уровне адаптивности к оптимальной солевой среде [3,11]. Выявлено стимулирующее влияние солёности на пластический обмен и уменьшение последствий воздействия искусственных условий выращивания в УЗВ на физиологическое состояние молоди.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование влияния солевой среды на молодь осетровых было выполнено в 2013 году в лаборатории водных биоресурсов и аквакультуры на базе экспериментального аквариального комплекса «Кагальник» Южного научного центра РАН. В качестве объекта исследований использовали молодь русского осетра (Acipenser gueldens-taedttii Brandt, 1896).
Экспериментальная молодь содержалась в 2-х пластиковых бассейнах (ИЦА-1) с пресной (контроль) и солёной (опыт) водой в диапазоне оптимальных параметров водной среды. Солевая среда – это раствор поваренной соли (NaCl) солёностью 5 ‰ (5 г/л). Концентрация соли контролировалась рефрактометром Klilong RHS-10ATC. Ежедневно 5% объёма экспериментальной среды обновлялось.
В течение всего периода исследования проводили наблюдения за термическим и гидрохимическим режимами. Температуру, кислород и рН измеряли три раза в сутки. Для этого использовали специальные приборы: термооксиметр Сyber Scan DO 300, рН-метр HANNA. Исследуемые гидрохимические показатели находились в пределах, допустимых для выращивания молоди осетровых рыб. Температурный режим поддерживался на уровне 20-22 °С, кислородный режим изменялся в пределах 90-100%, значения рН составляло 7-8 ед.
Для контроля и измерения содержания биогенных элементов в водной среде использовали экспресс-метод фирмы Tetra. Исследование ос- новных показателей качества воды выявило некоторое увеличение уровня нитритов (до 0,3 мг/л) и аммиака (до 0,5 мг/л). Систематически осуществлялся контроль за темпом роста рыбы. Взвешивание и измерение рыбы проводили согласно разработанным рекомендациям [12].
Состояние исследуемых рыб оценивали на основании морфологических показателей, выживаемости, коэффициента массонакопления [13], коэффициента упитанности по-Фультону. Среднесуточную скорость роста вычисляли по формуле сложных процентов [14]. Наблюдения за темпом роста осуществляли по мере выращивания рыб в опытах.
Физиологическое состояние рыб оценивали по концентрации гемоглобина в крови, СОЭ, уровню холестерина и общих сывороточных липидов, концентрации общего сывороточного белка. Взятие крови для исследований осуществляли прижизненно путем пункции хвостовой вены. Количество гемоглобина определяли колориметрическим способом, концентрацию холестерина в сыворотке крови энзиматическим колориметрическим методом, общий сывороточный белок определяли на рефрактометре ИРФ-22 [15], СОЭ определяли методом Панченкова. Общие липиды в сыворотке крови определяли с помощью набора реактивов фирмы PLIVA – Lachema [16,17]. Осмоляльность сыворотки крови определяли криоскопическим методом на осмометре ОСКР-1.
Рыб кормили гранулированным кормом «Bio-Mar» (1-2мм) по стандартным нормам с корректировками по поедаемости.
В исследованиях использовано 90 одновозрастных особей русского осетра, с обработкой 68 гематологических и биохимических проб. Выполнено 270 взвешиваний и измерений молоди русского осетра. Результаты исследований обработаны с применением общепринятых методов биологической статистики и программы Microsoft Exel. Уровень различий оценивали с помощью критерия достоверности Стьюдента.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
До начала эксперимента вся рыба находилась в пресной воде. Средняя масса молоди русского осетра, используемой в эксперименте, составляла 5,19±0,19 г (контрольный бассейн) и 5,52±0,21 г (опытный бассейн). Коэффициент упитанности по-Фультону находился на одном уровне и составлял 0,36±0,01 и 0,39±0,03 ед. в контрольном и опытном бассейнах соответственно. Исследование влияния солевой среды на темп роста молоди русского осетра проводили в несколько этапов. За весь период эксперимента наблюдалось стабильное увеличение массы, при любых условиях эксперимента (таблица 1).
В течение первых 10-ти дней исследования (1й этап) масса рыб, выращиваемых в пресной воде, увеличилась примерно на 21%. При выращивание в солености 5-‰ отмечено увеличение массы на 11%. Такие удельные массовые характеристики, как среднесуточный прирост, среднесуточная скорость роста, коэффициент массонакопления в контрольном бассейне были примерно в два раза выше, чем у рыб в бассейне с экспериментальной солевой средой.
Таблица. 1. Рыбоводно-биологические показатели молоди русского осетра в соленой (числитель) и пресной (знаменатель) воде
|
Рыбоводно-биологические показатели |
1 этап |
2 этап |
3 этап |
|
Средняя масса, г |
6,3 ± 0,3 6,1 ± 0,31 |
±0,40 ±0,40 |
8,7 ± 0,50 8,5 + 0,50 |
|
Абсолютный прирост, г |
1,11 0^58 |
1,8 15 |
0.6 06 |
|
Среднесуточный прирост, г |
0,11 Об |
0,2 0^ |
0,1 од |
|
Среднесуточная скорость роста, % |
106 |
2,6 |
0,7 0.8 |
|
Коэффициент массонакопления, ед. |
0,05 05 |
0,01 0^02 |
|
|
Коэффициент упитанности по Фультону, ед. |
0,31 ± 0,01 0,30 + 0,01 |
0,27 + 0,01 0,28 + 0,01 |
0,27 ± 0,01 0,29+ 0,01 |
|
Продолжительность выращивания, сут. |
10 |
10 |
7 |
Однако, наблюдалось некоторое снижение показателя упитанности рыб в обоих вариантах эксперимента. Такая динамика массовых характеристик в контроле и опыте, вероятно, была связана с адаптацией молоди не только к новым условиям содержания (температурный и кислородный режим, солёность среды обитания), но и кормлению.
На втором этапе исследования наблюдался более интенсивный темп роста молоди русского осетра в обоих вариантах. Однако в солевой среде такие удельные массовые характеристики, как среднесуточная скорость роста, среднесуточный прирост увеличились на 60 и 70%, соответственно, тогда как в пресной воде увеличение этих показателей произошло на 45 и 23%, соответственно.
На завершающем этапе исследования (3-й этап) темп роста массы значительно снизился, как при выращивании в пресной, так и в солевой среде. Вероятной причиной тотального снижения темпа роста рыбы в обоих вариантах на данном этапе экспериментальных работ могло быть некоторое ухудшение экологической ситуации в бассейнах за счёт не достаточно эффективной работы фильтров нейтрализующих азотистые соединения в условиях замкнутого цикла.
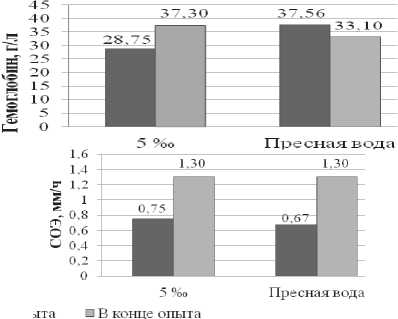
Рис. 1. Динамика гемоглобина и СОЭ у русского осетра в экспериментальных условиях (n=18)

5 9'Оо Пресная вода ■ В начале опыта □ В конце опыта
Рис. 2. Динамика общих сывороточных белков (г/л) у русского осетра в экспериментальных условиях (n=18)
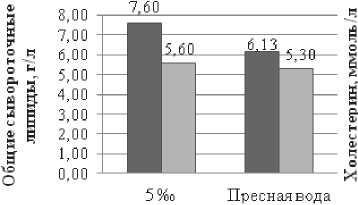
С целью выявления изменений в обменных процессах изучалось физиологическое состояние выращиваемых объектов. Молодь, поступившая в эксперимент была изначально физиологически гетерогенна и по ряду показателей отличалась не только между собой, но и от рыб, исследованных в Северном Каспии. В частности выявлено достоверное различие (р<0,05) по концентрации гемоглобина в крови и сывороточных липидов. Концентрация холестерина и общих сывороточных липидов у рыб в обоих вариантах эксперимента превышала значения, характерные для осетровых рыб Северного Каспия. Повышенный уровень показателей липидного обмена характерен для рыб питающихся искусственными кормами.
В тоже время низкие показатели скорости оседания эритроцитов и гемоглобина, у экспериментальных рыб содержащихся изначально в пресной воде в сравнении с рыбами из естественной среды, свидетельствовали о недостаточном уровне гидратации крови и низкой активности окислительного обмена. При обитании разновозрастных осетровых в естественной среде нормой принято считать следующие значения этих показателей: гемоглобин – 50–80 г/л, сывороточный белок – 28–40 г/л, сывороточные липиды – 3–4 г/л, холестерин – 1,0–2,8 ммоль/л, СОЭ – 2–4 мм/час [18].
На заключительном этапе эксперимента молодь осетра, содержащаяся 27 дней в солёной воде, по исследуемым физиологическим параметрам достоверно не отличалась от рыб, содержащихся аналогичный период в пресной воде.
Однако, если сравнивать функциональное состояние рыб выдержанных в солевой среде с их исходным состоянием (пресная вода), то отмечено, что за весь период выращивания в условиях эксперимента наблюдается достоверное повышение концентрации гемоглобина и существенное снижение концентрации сывороточных липидов (р≤0,001). Уровень холестерина изменился незначительно и находился в пределах, характерных для осетровых в Каспийском море. Выявленную динамику физиологических показателей у рыб выращенных в солёной воде можно рассматривать, как положительную тенденцию стабилизации их функционального состояния. Что под- тверждается динамикой массовых характеристик у опытных рыб.
У молоди осетра выращенной в пресной воде, наоборот, по завершении эксперимента от исходных значений снизился уровень окислительного обмена и, практически не изменилась достаточно высокая концентрация сывороточных липидов. К тому же у всей экспериментальной молоди в пресной воде достоверно (р≤0,001) снизился уровень холестерина, который является предшественником гормонов и входит в состав оболочек клеток. Выращивание в пресной воде не приводит к положительным сдвигам в физиологическом состоянии этих рыб.
■ В начале опыта
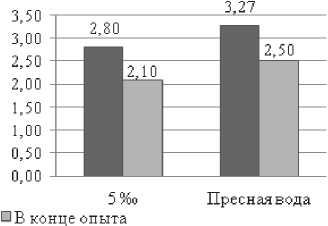
Рис. 3. Динамика биохимических показателей крови русского осетра в экспериментальных условиях (n=18)
Сравнительная оценка физиологического статуса выращиваемой молоди по концентрации осмотически активных веществ в сыворотке крови выявила гипертоничность особей по отношению к внешней среде, что характерно для разновозрастных осетровых в опреснённой зоне Северного Каспия (230-270 ммоль/кг H 2 O). В частности осмоляльность сыворотки крови в исходном состоянии (пресная вода) находилась на одном уровне у рыб в контрольном и опытном бассейнах, и составила соответственно 221,0±7,8 ммоль/кг H 2 O и 226,0±4,2 ммоль/кг H 2 O (р>0,05). В течение эксперимента осмоляльность сыворотки крови достоверно не изменилась как в опытном 228,0±1,41 ммоль/кг H 2 O, так и в контрольном вариантах 239,0±9,2ммоль/кгH 2 O (р>0,05). Осмоляльность солевой среды составила 134 ммоль/кг H 2 O, пресной воды – 20,0 ммоль/кг H 2 O. Гипертоничность среде обитания естественное состояние водно-солевого обмена у каспийских осетровых поддерживать которую в слабосолёной среде значительно легче, чем в пресной по причине излишней гидратации организма.
Таким образом, ранее установленный факт более высокого темпа роста у гибридных форм осетровых в солёной воде [11,19] подтвердился и при выращивании молоди русского осетра. Однако, наблюдаемое торможение прироста массы при выращивании в пресной и в солевой среде, возможно связано с более высокой чувствительностью русского осетра и его гибридов с сибирским осетром к параметрам окружающей среды и в частности к условиям выращивания в УЗВ.
Это свидетельствует о том, что своевременное вселение чистых линий проходных осетровых рыб в опреснённые участки моря становится объективным условием высокой выживаемости этих рыб и эффективного решения проблем пастбищной аквакультуры. Стимулирующее влияние ионов натрия на интенсивность всасывания пищевых компонентов в кишечнике рыб в данном случае рассматривается, как дополнительный аргумент в пользу ранней интродукции проходных видов осетровых в море [20,21].
При длительном выращивании особенно чистых линий осетровых в УЗВ искусственно создаваемые параметры водной среды должны приближаться к оптимальным значениям, что предполагает более эффективное регулирование и очистку от метаболитов. Необходимым условием выращивания качественной продукции в УЗВ является использование посадочного материала только с хорошим исходным физиологическим состоянием.
Список литературы Моделирование среды, как экологический способ решения актуальных проблем аквакультуры
- Ходоревская, Р.П. Современное состояние запасов осетровых каспийского бассейна и меры по их сохранению/Ходоревская Р.П., Калмыков В.А., Жилкин А.А.//Вестник АГТУ. Серия: Рыбное хозяйство. 2012. № 1. С. 99-106.
- Распопов В.М. Естественное воспроизводство рыбных ресурсов и техногенное общество (на примере осетровых)/Распопов В.М., Сергеева Ю.В.//Вестник АГТУ: Серия Рыбное хозяйство. 2009. № 2. С. 14-16.
- Матишов Г.Г. Практическая аквакультура (Разработки ЮНЦ РАН и ММБИ КНЦ РАН)/Матишов Г.Г., Е.Н.Пономарева, Н.Г.Журавлева. -Ростов н/Д.: ЮНЦ РАН. 284 с.
- Гербильский Н.Л. Теория биологического прогресса осетровых и её применение в практике осетрового хозяйства/Гербильский Н.Л.//Учёные записки Ленинградского ун-та. Сер. биол. наук. 1962. Вып. 48. Часть 2. № 311. С. 5-18.
- Дюбин В.П. Эвригалинность молоди севрюги на ранних этапах онтогенеза/Дюбин В.П.//Тезисы отчет. сессии ЦНИОРХ. 1972. С. 50-51.
- Краюшкина Л.С. Каспийские осетровые и эволюция механизмов осмотической и ионной регуляции у Acipenseridae/Краюшкина Л.С.//Первый конгресс ихтиологов России: Тезисы докладов. М.: ВНИРО, 1997. С. 432.
- Краюшкина Л.С. Особенности осмотической и ионной регуляции у морских проходных осетров -коротконосого Acipenser brevirostrum и острорылого A. Oxyrhynchus (Acipenseridae)/Краюшкина Л.С.//Вопросы ихтиологии. 1998. Т. 38. № 5. С. 684-692.
- Гершанович А.Д. Экология и физиология молоди осетровых/Гершанович А.Д., Пегасов В.А., Шатунов М.И. М.: Агропромиздат, 1987. 215 с.
- Семенова О.Г. Адаптация пресноводных и солоноватоводных видов осетрообразных (Acipenseriformes) к гиперосмотической среде: Автореф. дисс. биол.наук. Санкт-Петербург, 2002. 24 с.
- Металлов Г.Ф. Физиолого-биохимические механизмы эколого-адаптационной пластичности осморегулирующей системы осетровых рыб/Металлов Г.Ф., Пономарев С.В., Аксенов В.П., Гераскин П.П. Астрахань: АГТУ, 2010. 192 с.
- Пономарёва Е.Н. Динамика функционального состояния молоди гибрида русско-ленского осетра при моделировании условий выращивания в установке замкнутого водоснабжения/Пономарёва Е.Н., Металлов Г.Ф., Григорьев В.А., Ковалёва А.В., Пономарёв С.В., Левина О.А.//Известия вузов. Северо-Кавказский регион. Серия: Естественные науки. 2012. № 5. С. 72-76.
- Правдин И.Ф. Руководство по изучению рыб/И.Ф. Правдин. М.: Пищевая промышленность, 1966. 376 с.
- Купинский С.В. Радужная форель -предварительные параметры стандартной модели массонакопления/C.В. Купинский, С.А. Баранов, В.Ф. Резников//Сборник научных трудов: Индустриальное рыбоводство в замкнутых системах. М.: ВНИИПРХ, 1985. Вып. 46. С. 109-115.
- Castell, J.D. Report of the EIFAC, IUNS and ICES Working Group on the standartization of the methodology in fish nutrition research. EIFAC Tech/Castell J.D., K.Tiews//Pap. Hamburg, 1979. P. 1-24.
- Филиппович Ю.Б. Практикум по общей биохимии/Ю.Б. Филиппович, Т.А. Егорова, Г.А. Севастьянова. -М.: Просвещение, 1975. 318 с.
- Zolner, N. Uber die quantitave Bestimmung von Lipoiden (micromethode mittels die vieles naturlischen Lipoiden allen Bekannten plasmolipoiden) gemeinsamen sulfophosphovanilin-reaction/N.Zolner, K. Z.Kirch//Zeitschrift fur die gesamte experimentelle Medicin. 1962. Vol. 135. № 6. Р. 545-561.
- Knight J. Chemical basis of the sulfo-phospho-vanillin reaction for estimating total serum lipids/J.Knight, S.Anderson, J.Rawle//Clin Chem, 1972. Vol. 18. Р. 199-202.
- Металлов Г.Ф. Биохимические и морфофизиологические показатели русского осетра в современных экологических условиях Волго-Каспия/Металлов Г.Ф., Распопов В.М., Аксенов В.П., Чипинов В.Г.//Сборник материалов и докладов международного симпозиума «Тепловодная аквакультура и биологическая продуктивность водоемов аридного климата» (Астрахань 16-18 апреля 2007 г.). Астрахань, 2007. С. 484-486.
- Григорьев В.А. Влияние солености воды на рост и развитие гибридных форм осетровых рыб/Григорьев В.А., Ковалева А.В., Корчунов А.А.//Международная конференция «Осетровые рыбы и их будущее» (7-10 июня 2011 г.)/-Бердянск, 2011. С. 109-112.
- Sastry K.V. Effect of inhibitors on Na-dependent D-glucose transport in the small intestine of two teleost fishes/Sastry K.V., Garg V.K., Agraval V.P.//Indian. J. exp. Biol. 1977. Vol. 15. № 8. P. 661-662.
- Boge G. The use of intestinal brush border membrane vesicles for comparative studies of glucose and amino isobutyric acid transport by pour species of marine teleost/Boge G., Rigal A., Peres G.//Comp. Biochem. Physiol. 1982. Vol. 72. № l. P. 85-89.
