Модельные исследования действия солей кадмия и стронция на фотосинтетические показатели пшеницы

Автор: Пиняскина Александра Владимировна, Пиняскина Елена Владимировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 5-1 т.17, 2015 года.
Бесплатный доступ
Изучали влияние солей кадмия и стронция на флуоресцентные характеристики фотосинтетических пигментных комплексов изогенных линий пшеницы с. «Мироновская 808». Показано, что инкубация проростков пшеницы в растворах CdCl 2 и SrCl 2 подавляла ростовые процессы, снижала интенсивность фотосинтеза. В постстрессовый период фотосинтез усиливался. Анализ индукционных кривых флуоресценции хлорофилла всех сортов (Rht 9, 13, 17) показал, что на ранних стадиях токсическое действие Cd и Sr в исследованных концентрациях проявляется не только в подавлении электронного транспорта ФСII, но и в уменьшении степени энергизации фотосинтетических мембран. Начальные процессы развития токсического действия Cd и Sr связаны со снижением эффективности световых реакций фотосинтеза, что и определяет в дальнейшем ухудшение продукционных свойств.
Растение, фотосинтез, флуоресценция, тяжелые металлы
Короткий адрес: https://sciup.org/148204008
IDR: 148204008 | УДК: 577.355.4:
Model researches of cadmium and strontium salts impact on the wheat photosynthetic indicators
It was studied the impact of cadmium and strontium salts on fluorescent characteristics of photosynthetic pigmentary complexes of wheat isogene lines sort "Mironovskaya 808". It is shown that the incubation of wheat sprouts in CdCl 2 and SrCl 2 solutions suppressed growth processes, reduced intensity of photosynthesis. During the post-stressful period photosynthesis amplified. The analysis of induction curves of fluorescence of chlorophyll of all grades (Rht 9, 13, 17) showed that at early stages toxic impact of Cd and Sr in the studied concentration is shown not only in suppression of electronic transport of FSII, but also in reduction of degree of energization the photosynthetic membranes. Initial developments of toxic impact of Cd and Sr are connected with decrease in efficiency of light reactions of photosynthesis, as defines further deterioration of productional properties.
Текст научной статьи Модельные исследования действия солей кадмия и стронция на фотосинтетические показатели пшеницы
постоянная аэрация раствора). На 3 сутки после проклевывания проростки переносили в контейнеры с керамзитом. Семена были выращены в растворах хлорида кадмия с концентрациями 1*10-4 Моль/л, 1*10-5 Моль/л, 1*10-6 Моль/л и в растворах хлорида стронция в тех же концентрациях. Контрольные образцы были выращены на воде. Измерения проростков пшеницы производились на 5, 10 и15 сутки после проклевывания. Учитывали темпы прорастания, общую всхожесть, прирост побегов, количество корней и длину наибольшего корня. Количество повторностей – 10.
Параметры флуоресценции хлорофилла а листьев измеряли с помощью флуориметра MINI-PAM (Pulse Amplitude Modulation) Yeinz Walz GmbH, (Германия) методом амплитудной импульсной модуляции света. Перед измерением побеги были адаптированы к темноте в течение 10 мин. В ходе экспериментов регистрировали следующие параметры флуоресценции: F 0 – интенсивность флуоресценции хлорофилла в адаптированных к темноте образцах при действии зондирующих импульсов возбуждающего света; Fm — интенсивность флуоресценции хлорофилла во время действия 0,8 с насыщающей вспышки света, восстанавливающей первичные хинонные акцепторы Q A до Q А * ; Y - эффективность процессов фотосинтеза; qP и qN - коэффициенты химического и нехимического фототушения флуоресценции, NPQ - параметр нехимического фототушения флуоресценции.
Для возбуждения флуоресценции хлорофилла используется импульсная модуляция красного света, источником которого служит «красный» светодиод (LED), максимум излучения при длине волны 650 нм. Продолжительность импульсов 3 мкс модуляционная частота 0,6 кГц, режиме укороченной вспышки (burst-mode) вспышки продолжительностью 0,2 сек. чередуются с периодами темноты 0,8 сек. Свет светодиода проходит через фильтр (Balzers DT Cyan, special), с максимумом пропускания 650 нм и оставляющий небольшой участок спектра более 700 нм. Флуоресценция регистрировалась с помощью трехслойного полупроводникового фотодиода (PIN-фотодиод) при длине волны более 700 нм, благодаря наличию длинноволнового фильтра (RG 9, Schott). Интенсивность 0,15 мкмоль м-2с-1 ФАР; Источником насыщающих световых импульсов и постоянного актиничного освещения служит галогенная лампа 8 Вт /20 Вт (Bellaphot. Osram). Статистическую обработку данных проводили с помощью пакетов стандартных программ Microsoft Excel 2007 и Statistica 6. В таблицах и на рисунках представлены средние значения и стандартные отклонения. Достоверность различий между парными значениями оценивали по t –критерию Стьюдента при P<0.05.
Результаты исследований. По всхожести наиболее устойчивым к ТМ является Rht 9. Концентрация хлорида кадмия 10-4 наиболее ингибирующая для всех линий. Rht 13 и Rht 17 схоже реагировали на соли кадмия. В то же время стронций ингибирует всхожесть Rht 13 во всех концентрациях на 45-50%. Наиболее устойчивыми к действию стронция оказались линии Rht 9 и Rht 17. На Rht 17 все концентрации влияют практически одинаково, и всхожесть составляет 75-85% от контроля (рис. 1).
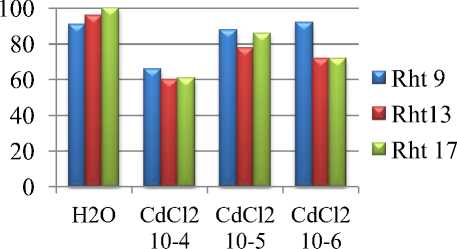
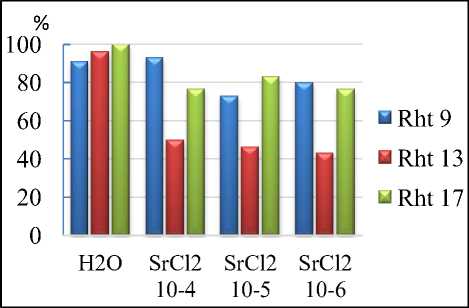
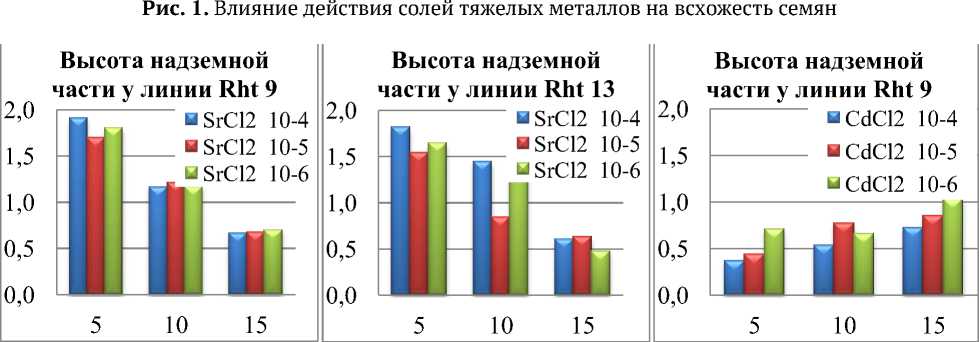
Рис. 2. Изменение надземной части линий Rht 9 и Rht 13 в зависимости от концентрации солей кадмия и стронция
У линии Rht 9 под действием солей стронция наблюдается ингибирующий эффект по всем показателям, который усиливается к 15 суткам. Изменение концентрации никак не влияет на отдельные измеряемые морфологические показатели и оказывает общее ингибирующее действие (рис. 2а, б). При действии же различных концентраций хлорида кадмия на начальных этапах развития наблюдается замедление роста; по видимому к 15 суткам происходит адаптация к данному виду стресса, в результате чего показатели длины гипокотиля близки к контрольным и составляют 73-103%.
У линии Rht 17 на 10 сутки наблюдается резкий скачок ростовых процессов как корневой, так и надземной части при воздействии солями стронция всех концентраций и солями кадмия 105-10-6 М. Концентрации хлорида стронция, ингибирующие на 10 сутки корневую систему, стимулируют ростовые процессы гипокотиля. Необходимо отметить, что концентрации ингибирующие рост гипокотиля стимулируют рост корня в длину. Таким образом, наблюдается обратная корреляция между длиной корневой системой и длиной гипокотиля. В случае же с хлоридом кадмия наблюдается прямо пропорциональная зависимость этих показателей. В растворах с солями стронция у линий Rht 9 и Rht 13 изменения числа корней по отношению к контролю практически не происходит, а у сорта 17 наблюдается прирост как при действии солей стронция, так и при действии хлорида кадмия в концентрациях 10-5-10-6 М.
Измерение длины надземной части у всех линий, выращенных в различных концентрациях стронция, показало прирост по отношению к контролю на 5 сутки и его дальнейшее постепенное ингибирование у Rht 9 и Rht 13. С другой стороны, почти во всех опытных образцах при воздействии солями кадмия максимальное ингибирование происходит именно к 5 суткам. Из исследуемых концентраций кадмия наибольшим ингибирующим эффектом для всех морфологических показателей обладает CdCl2 10-4 М. Слабым ингибитором практически по всем показателям является хлорид кадмия в концентрации 10-6 М. Наиболее устойчивой к действию солей кадмия оказалась линия Rht 9. Хлорид стронция для всех линий и во всех концентрациях показал себя сильным ингибитором, замедляя рост по отношению к контролю до 70%. Необходимо отметить факт коррелятивной связи между количеством и длиной корней при действии солей стронция. Увеличение длины корней сопряжено со снижением их числа, что особенно заметно у Rht 13 и Rht 17.
Исследуя действие солей металлов на проростки пшеницы разных линий мы работали в основном с биометрическими параметрам этих растений. Так как биометрические показатели являются результатом изменений на молекулярном уровне, мы изучали действие ТМ на процессы фотосинтеза. В результате исследований нами было показано, что квантовый выход флуоресценции ( F ) как и максимальной флуоресценции хлорофилла ( Fm *) у всех линий пшеницы при концентрации Sr 10-6 моль/л совпадают с контрольными (по воде) или являются максимальным (рис. 3 а, б, в). Снижение квантового выхода ( F ) у сорта Rht 13 указывает на фотоповреждение фотосинтезирующих систем.
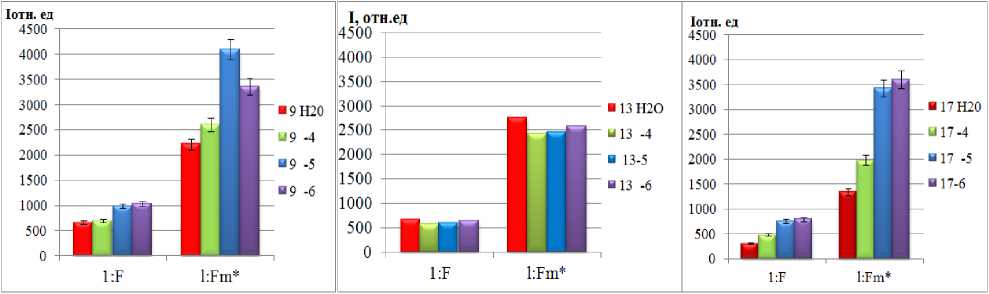
а б в
Рис. 3. Квантовые выходы флуоресценции (F) и максимальной флуоресценции (Fm) линий Rht 9,13,17 в зависимости от концентрации солей Sr
Выход максимальный флуоресценции (Fm*), измеренный после полной темновой адаптации (когда все реакционные центры открыты, и все первичные акцепторы электронов окислены) у линий Rht 9 и 17 максимален, при минимальном использовании энергии на фотохимические реакции и тепловое излучение. Уменьшение Fm* у линии Rht 13 (при всех разведениях) свидетельствует об увеличении безызлучательных потерь световой энергии – тепловая диссипация. Анализ кривых тушения флуоресценции (NPQ) показывает увеличение значений, приближаясь к контрольным (относительно воды), что свидетельствует об интенсификации процессов нефотохимического тушения, связанных с рассеиванием энергии возбуждения антенного хлорофилла в виде тепла. В целом, по рис. 3 видно, что наиболее чувствительной к ТМ является линия Rht 13, даже слабая концентрация растворов металлов у которого оказывает негативное влияние на фотосинтетическую активность: уменьшение F и Fm*, рост qP и qN. Что же касается высоких концентраций ТМ, то изучение индукционных кривых флуоресценции указывает на индукцию процессов нефотохимического тушения (см. рис.4 б), концентрация 10-6 моль/л вы- звала снижение фотосинтетической активности ниже исходного уровня.
При изучении кинетики восстановления фотосинтетического аппарата наблюдалась та же картина: максимально быстро восстанавливались сорта Rth 17 (1мин.) и 13 (1,5 мин.), тогда как Rth 9 потребовался более продолжительный период времени до 4 минут. При изучении влияния разных концентраций кадмия на проростки, отметим, что не все образцы проросли: на 15 сутки зафиксировали хлороз листьев, соответственно, говорить об эффективность фотосинтетических процессов не приходится. Квантовые выходы флуоресценции (F) и максимальной флуоресценции (Fm) линий Rht 9, 13, выращенных в растворе Cd при концентрации 10-6 моль/л снижались и были меньше контрольных ( по воде) (рис. 5).
Для того, чтобы определить же какие реакции идут, мы сняли характеристические кривые NPQ, qP и qN отражающие ту часть нефотохимического тушения, которая связана с рассеиванием энергии возбуждения антенного хлорофилла в виде тепла. В контрольных образцах Rht 9(Н 2 О) было отмечено резкое увеличение qN, связанное с нарушением утилизации фотосинтетической энергии.
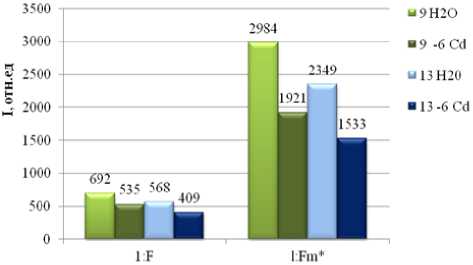
Рис. 5. Квантовый выход флуоресценции (F)и максимальной флуоресценции (Fm) линий Rht 9, 13, выращенных в растворе Cd
Rhtl3 Н2О
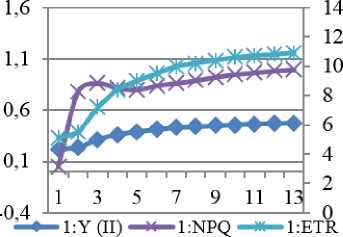
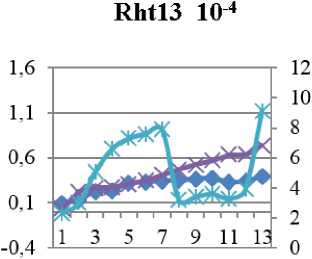
Рис. 4. Индукционные кривые пшеницы линии Rht 13 - контроль (Н 2 О) и опыт (концентрации Sr10-4 и 10-6М). По левой вертикальной оси – интенсивность флуоресценции (I, отн. ед.), по правой вертикальной – скорость электронов по ETR (µmol electrons/(m2·s).
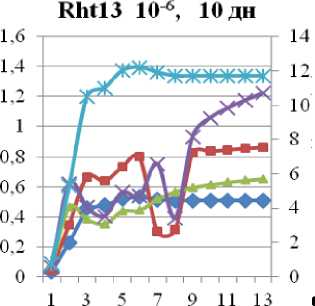
Уменьшение же qP в опытном образце мы связываем с нарушениями в цепи транспорта электронов. Отмечено резкое падение тока электронов и снижение тепловой диссипации на фоне падения эффективности фотосинтеза почти в два раза от контрольной. Гораздо хуже дело обстоит у сорта Rth 13: (см.рис. 6 в) видно, что поток электронов не стабилен, более того, он дважды падает практически до 0.
Используя кривые индукции флуоресценции, мы разделили различные типы флуоресценции, приводящие к снижению квантового выхода ФС II. Быстрое восстановление, происходящее в первые 30-60 секунд, отражает перераспределение энергии связанное с нефотохимическим тушением флуоресценции. Более медленные процессы нефотохимического тушения, протекающие в первые 10-30 минут связаны с перераспределением энергии возбуждения в пользу ФС II.
Исходя из анализа формы, кинетики индукционной кривой (её медленной - восстановительной фазы), можем говорить о восстановлении различных компонентов нефотохимического тушения флуоресценции, степени фотоингибирования, электронном транспорте между стромой (или цитоплазмой) и переносчиками электронов в тилакоидных мембранах. Эффективность преобразования энергии в процессе фотосинтеза зафиксирована у сорта Rth 9 на уровне 0,758, несколько ниже у сорта Rth 13, в то время, как в контроле Y=0,77 и 0,758, соответственно. Теоретически максимальное значение Y имеют растения, растущие в благоприятных условиях 0,83, однако на практике этот показатель несколько ниже - 0,8 (рис. 7).
Rht 9 Н2О
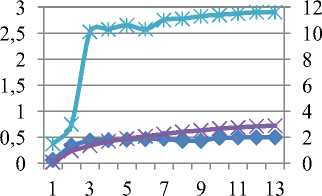
1:Y (II) 1:NPQ 1:ETR
а
Rht 9 10-6 Cd
2,5
1,5
0,5
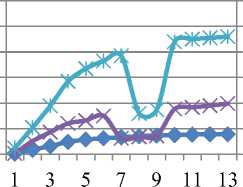
б
Rht 13 10-6 Cd
2,5
1,5
0,5
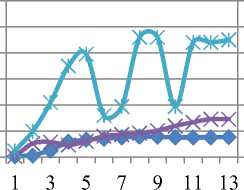
в
Рис. 6. Индукционные кривые сортов Rth 9 и Rth 13. По левой вертикальной оси – интенсивность флуоресценции (I, отн. ед.), по правой вертикальной – ETR (µmol electrons/(m2·s)
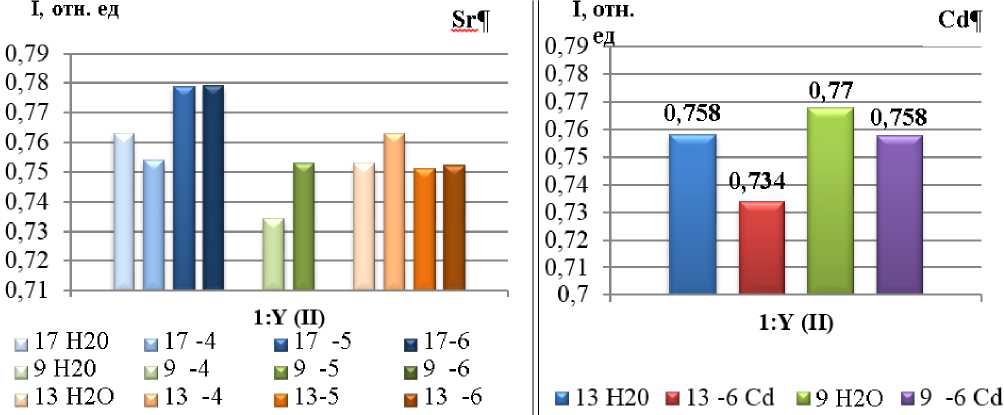
Рис. 7. Эффективность фотосинтеза у сортов Rh 9, 13, 17, выращенных при различных концентрациях ТМ
Выводы: соли кадмия и стронция по-разному влияют на морфофизиологические показатели и фотосинтез растений. Степень ингибирования фотосинтеза во многом зависит от концентрации металла в субстрате. Высокие концентрации металлов влияют на ультраструктуру хлоропластов, замедляют скорость электронного транспорта, уменьшая содержание фотосинтетических пигментов и активность ферментов цикла Кальвина как за счет их прямого действия на отдельные реакции фотосинтеза, так и в результате опосредованного влияния на другие физиологические процессы.
Список литературы Модельные исследования действия солей кадмия и стронция на фотосинтетические показатели пшеницы
- Лукин, С.В. Содержание меди и свинца в почвах Белогородской области/С.В. Лукин, С.В. Меленцева, П.М. Авраменко//Агрохимия. 2009. №2. С. 71-74.
- Минкина, Т.М. Влияние различных мелиорантов на подвижность цинка и свинца в загрязненном черноземе/Т.М. Минкина, Г.В. Мотузова, О.Г. Назаренко и др.//Агрохимия. 2007. № 10. С. 67-75.
- Минеев, В.Г. Химизация земледелия и природная среда. -М.: Агропромиздат, 990. С. 287.
- Прохорова, Н.В. К вопросу о фоновой концентрации в почвах Самарской области//Самарская Лука: Бюлл. -Самара, 2002. №2. С. 145-149.
- Sinha, P. A core-extraction distributed ad hoc routing algorithm/P. Sinha, R. Sivakumar, V. Bharghavan//Selected Areas in Communications, IEEE Journal. 1999. №17 (8). P. 1454-1465.
