Модифицирующее действие экстрактов кожицы винограда и красного вина на чувствительность бактерий Escherichia coli к различным антибиотикам

Автор: Безматерных К.В., Смирнова Г.В., Октябрьский О.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
Экстракты красного вина и кожицы винограда содержат значительное количество полифенолов, включая активные антиоксиданты кверцетин и ресвератрол, и проявляют высокую антирадикальную и хелатирующую способность. Внесение экстрактов в культуру E. coli вызывает 30%-ное ин-гибирование скорости роста, индукцию антиоксидантных генов katG и sodA и повышение устойчивости бактерий к перекиси водорода. Предобработка клеток E. coli каждым из экстрактов снижает бактерицидную активность ципрофлоксацина и цефотаксима и, наоборот, усиливает чувствительность бактерий к канамицину и стрептомицину. Воздействие экстрактов осуществляется, по-видимому, путем их влияния на скорость роста бактерий, редокс-статус клеток и уровень экспрессии антиоксидантных генов. В случае ципрофлоксацина, предобработка экстрактами может снижать степень повреждения ДНК и уровень индукции SOS-ответа. Модулирующий эффект экстрактов должен учитываться при антибиотикотерапии.
Растительные экстракты, полифенолы, антиоксидантные гены, антибиотики
Короткий адрес: https://sciup.org/147204790
IDR: 147204790 | УДК: 579.22
Modifying effect of grape skin and red wine extracts on Escherichia coli susceptibility to various antibiotics
Extracts from red wine and grape skin contain a significant amount of polyphenols, including quercetin and resveratrol, and possess high antiradical and chelating ability. Addition of these extracts into E. coli culture causes a 30% inhibition of the growth rate, an induction of antioxidant genes katG and sodA and a rise in H2O2 resistance. Pretreatment of E. coli with the extracts attenuates bactericidal activity of ciprofloxacin and cefotaxim and, in contrast, augments E. coli susceptibility to kanamycin and streptomycin. The extracts may exert their action by influence on the growth rate, redox state of cells and expression of antioxidant genes. In the case of ciprofloxacin, pretreatment with the extracts can decrease DNA damage and the level of the SOS response. The modulating effect of extracts should be taken into consideration in antibiotic therapy.
Текст научной статьи Модифицирующее действие экстрактов кожицы винограда и красного вина на чувствительность бактерий Escherichia coli к различным антибиотикам
Полифенолы, являющиеся обычными компонентами продуктов растительного происхождения и входящие в состав медицинских и косметических препаратов, привлекают растущий интерес в связи с их положительным влиянием на здоровье чело века [Crozier, Jaganath. Clifford. 2009]. Эти соединения обладают антиоксидантными свойствами благодаря способности связывать свободные радикалы и хелатировать ионы железа [Rice-Evans et al. 1995; Perron. Brumaghim. 2009]. Вместе с тем, в аэробных условиях полифснолы могут подвергаться аутоокиелению и генерировать активные
(С Безматерных К. В., Смирнова Г. В., Октябрьский О. Н., 2016
формы кислорода, выступая в качестве прооксидантов [Tang, Halliwell, 2010], При действии на живые клетки полифенолы модулируют передачу7 внутриклеточных сигналов и влияют на регуляцию экспрессии генов и активности ферментов* участвующих в ответе на окислительный и другие стрессы [Eberhardt, Jeffery', 2006; Smirnova et aL 2009]*
Высокая антиоксидантная и антипролифера-тивная активность ряда полифенолов (кверцетин, ресвератрол и др.) обусловливает их широкое использование в составе биологически активных добавок и лекарственных препаратов при профилактике и лечении заболеваний, связанных с окислительным стрессом (рак, диабек сердечнососудистые и другие). Кроме того* полифенолы являются компонентами экстрактов лекарственных растений, широко применяемых в традиционной медицине. В последние годы показано, что действие антибиотиков на бактериальные клетки приводит к накоплению окислительных повреждений во всех типах макромолекул (ДНК, белки. липиды), что вносит вклад Б бактерицидный эффект [Belenky el at* 2015]. В связи с этим можно ожидать, что при одновременном применении, воздействие на бактерии полифенолов, обладающих антиоксидантными свойствами, может интерферировать с действием антибиотиков, влияя тем самым на эффективность ан-тибиотикотерапии. Ранее мы и другие исследователи показали, что зеленый и черный чай и экстракты некоторых лекарственных растений изменяют чувствительность бактерий Е. cob к антибиотикам разных классов [Smirnova el aL 2012; Maralhe el al., 2013; Samoilova el aL 2014],
Целью настоящей работы является изучение влияния экстрактов вина и кожицы красного винограда, которые ЯВЛЯЮТСЯ богатыми источниками полифенолов, включая кверцетин и ресвератрол, на чувствительность бактерий Е. сой к антибиотикам с разным механизмом действия (ципрофлоксацин, канамицин, стрептомицин и цефотаксим),
Материалы и методы исследования
Приготовление экстрактов. Красное виноградное вино “PRIOS” (Испания) упаривали на роторном испарителе IKA RV10 (Германия) и лио-фильно высушивали. Сухой экстракт разводили в ДМСО. Конечная концентрация экстракта вина в экспериментах с бактериями составляла 3,6 мг/мл. Сухую кожицу7 красного винограда (10 г) размалывали и экстрагировали 3 раза по 30 мин, раствором Этиловый спирт : вода в соотношении 4 : 1 (v/v) на ультразвуковой водяной бане (El masonic S10 Н, Elma, 37 kHz, 30W) при температуре 60°С, Объединенный экстракт упаривали на роторном испа рителе и лиофильно высушивали, Конечная концентрация экстракта кожицы в экспериментах с бактериями составляла 3,8 мг/мл.
Общее содержание полифеполов в экстрак тах измеряли модифицированным методом Folin-Ciocalteu [Wu et aL 2006]. Экстракты (10 мкл) смешивали с 40 мкл реагента Fol in-Cioca lieu, встряхивали в течение 15 сек. и инкубировали 3 мин. при комнатной температуре. Затем добавляли 100 мкл 7%-ного карбоната натрия, и смесь доводили деионизированной водой до 3 мл. Через 90 мин. инкубации при комнатной температуре измеряли поглощение при 760 нм, используя спектрофотометр Shimadzu UV-VIS. Полученные значения сравнивали со стандартной кривой, построенной для растворов галловой кислоты, и выражали как эквиваленты галловой кислоты (мкг GAE/мг сухого экстракта).
Содержание отдельных поли фенолов в экстрактах определяли методом ВЭЖХ на жидкостном хроматографе Shimadzu LC-20AD. Разделение проводили на колонке С18 с использованием растворителей: А - ацетонитрил: В - смесь бидистил-лированной воды и уксусной кислоты в соотношении 40 : 1. Скорость потока - 1 мл/мин. Режим подачи растворителей: 0-15 мин., А - 14%, В - 86%; 16—15 мин.. А - 35%. В - 65%; 46-48 мин.. А -100%. Анализ пиков осуществляли при длинах волн 220-400 нм.
Хелатирующую активность экстрактов оп ределяли модифицированным методом [Kim et al„ 2005]. Реакционную смесь, содержащую 50 мкл образца и 10 мкл 1 мМ FeS04, активировали добавлением 6.7 мкл 5 мМ феррозина в ацетатном буфере, Раствор перемешивали и инкубировали при комнатной температуре в течение 10 мин,, после чего определяли поглощение при длине волны 562 нм. Хелатирующую активность рассчитывали, используя уравнение
Хелатирующий эффект (%) = ((Поглощение контроля - Поглощение образца) / Поглощение контроля) х 100,
График зависимости хелатирующего эффекта от концентрации исследуемого образца использовали для расчета ЕС?о как концентрации, при которой Fe2^ ионы связываются на 50%.
Анти радикальную активность полифенолов и экстрактов определяли по их способности связывать стабильный DPPH радикал [Shyur et aL, 2005]. Определение проводили в реакционной смеси, содержащей 3 мл 0.3 мМ DPPH'-этанольного раствора, 1 мл 50 мМ Tris-HCl буфера (pH 7.4) и 5-20 мкл экстракта. После 30 мин. инкубации при комнатной температуре измеряли поглощение при
517 нм. Ингибирующий эффект на уровень DPPH* рассчитывали согласно формуле
Ингибирующий эффект (%) = ((Поглощение контроля - Поглощение образца) / Поглощение контроля) х 100.
Строили график зависимости ингибирующего эффекта от концентрации исследуемых соединений и определяли величину7 1С5о как концентрацию, при которой связывается 50% свободных радикалов DPPH*.
Штаммы бактерий и условия культивирования. В качестве объекта исследований использовали штамм Е. coh BW25113 ^(araD-araB^CI, MacZ47^3(; ;гтВ-3\ V, rphA, ^(rhaD-rhaB)56^, hsdRSW, полученный из Е. coii Genetic Stock Center (CGSC). Штаммы NM3001 и NM3011, несущие слияния промоторов so^Zd и sail (sfb4) генов со структурным геном lacZ. кодирующим 0-галактозидазу. были сконструированы путем трансдукции фагом PI слияний sodA.dacZ и 5mlA(sfiA)\lacZ из Е, coli DM4000 (дар проф. М. Volkert) и QC772 (дар проф. D. Touati) в BW25113. Штамм NM3021, несущий транскрипционное генное слияние katG::lacZs был получен путем трансформации клеток К colt BW25113 плазмидой рКТЮЗЗ [Tao et al, 1989]. Штаммы NM3031 и NM3041 со слияниями katE.:lacZ к rpoS(katF)\dacZ были созданы путем трансформации клеток Е. сок BW25113 плазмидами pRS KatE 16 (дар проф. М. Volkert) и pRS 415 KatF5 (дар проф. A. Eisenstark).
Бактерии выращивали на минимальной среде М9 (Na2HPO^12H2O -15.13 г/л; КН2РО, - 3 г/л; NH.C1 - 1 г/л; NaCl - 0.5 г/л; MgSO^7H2O - 0.246 г/л; СаС12 - 0.011 г/л) с добавлением 0.15% глюкозы, 0.2% казаминовых кислот и тиамина (10 мкг/мл). За ростом следили путем измерения оптической плотности при длине волны 600 нм (ODy * ).
Удельную скорость роста культуры (ц) рассчитывали по формуле где OD^otti и ООб(*)№) - оптическая плотность культуры, измеренная при длине волны 600 нм, во время Г2 и 6,1 - время в часах.
Чувствительность Е* coll к H2Oi и антибиотикам определяли микропланшетным методом. Клетки из ночной культуры центрифугировали и переносили в колбы объемом 250 мл, содержащие 100 мл среды М9, до начальной OD^ = 0Л и выращивали на качалках при скорости вращения 140 об/мин и температуре 37°С до ОО6См> = 0.6, Далее культуру центрифугировали и ресуспендировали в 8 мл среды М9, В ячейки планшета добавляли по 5
мкл исследуемых экстрактов, 5 мкл концентрированных клеток и среду7 М9 до общего объема 200 мкл. Планшеты инкубировали на качалках (140 об/мин, температура 37°С) 20 мин,, измеряли OD^io и в опытные ячейки вносили Н2О2 (4 мМ) или антибиотики ципрофлоксацин (0.03, 0,3 и 3 мкг/мл), стрептомицин (10 и 40 мкг/мл), канами-цин (10 и 40 мкг/мл) и цефотаксим (5 и 10 мкг/мл). Отбор проб и измерение OD^ на микропланшет-ном спектрофотометре BioRad xMark, проводили до и через 30 и 70 мин, после внесения антибиотиков, Индекс антиоксидантной активности (АОА) рассчитывали как отношение удельной скорости роста бактерий, предобработанных экстрактами, к удельной скорости роста без предобработки через 30 мин, после добавления Н2О2.
Колопнеобразующую способность определяли в образцах из контрольной культуры и культур, обработанных экстрактами и антибиотиками. Пробы отмывали, разводили 0.9%-ным NaCl, капли полученной суспензии (10 мкл) помещали на чашки с LB-агаром и инкубировали в термостате при 37°С. Количество образовавшихся колоний (КОЕ) подсчитывали через 24 ч. инкубации.
Статиста чес кую обработку экс пер име1 ■-тальпых данных осуществляли с помощью пакета программ Microsoft Excel и Statistica 6,0. Каждый результат показан как среднее значение по меньшей мере трех-пяти независимых экспериментов ± стандартная ошибка среднего, Достоверность различий была определена с помощью t-критерия Стьюдента, статистически значимыми считались различия данных при уровне значимости р <0,05.
Результаты и их обсуждение
Для понимания биологических эффектов экстрактов красного вина и кожицы винограда был предварительно изучен их состав, а также хелатирующая и антиоксидантная активность. Состав и свойства исследуемых экстрактов представлены в табл. 1. Общий уровень полифенолов в экстракте вина был в 2 раза выше, чем в экстракте кожицы винограда. Оба экстракта содержали примерно одинаковое количество ресвератрола, но существенно отличались по уровню кверцетина, которого в экстракте кожицы было в 2 раза больше, чем в экстракте красного вина.
Кверцетин обладает высокой хелатирующей способностью (ЕСso = 0*004 мкг), и его повышенное содержание в экстракте кожицы может быть причиной в 2 раза более высокой хелатирующей активности этого экстракта* Напротив, значение IC50 было ниже, то есть антирадикальная активность выше у экстракта вина* чем у экстракта кожицы. Отсутствие параллелизма между хелатирующей и антпрадикальной активностями может
Содержание полифенолов, хелатирующая спосо объясняться тем* что отдельные компоненты экстрактов, например ресвератрол, проявляют выраженную антирадпкальную активность, но при этом не хелатируют железо. В целом способность экстрактов связывать радикалы DPPH* была в 20-30 раз ниже по сравнению с чистым препаратом кверцетина и в 6-9 раз ниже по сравнению с ресвератролом (не показано).
Таблица 1 ость и анти радикальная активность экстрактов красного вина и кожицы винограда
|
Состав и свойства |
Экстракт вина |
Экстракт кожицы винограда |
|
Общие полифенолы, мг GAE/r экстракта |
111 ±2 |
54 ± 1 |
|
Ресвератрол, мкг/г экстракта |
81.5 |
75.9 |
|
Кверцетин, мкг/г экстракта |
55.1 |
115.6 |
|
ЕСзи, мг экстракта |
0.169 |
0.082 |
|
ТС50, мг экстракта |
0.3X6 |
0.536 |
Внесение в культуральную среду экстрактов вина и кожицы винограда приводило к снижению удельной скорости роста К. сой на 28 и 30% по Сравнению С контролем. Соответственно, в Присутствии экстрактов замедлялся прирост числа колониеобразующих единиц (КОЕ). Таким образом, оба экстракта демонстрировали умеренное бактериостатическое действие па клетки К. coli. Бактериостатический эффект был, по-видимому, обусловлен Суммарным ДеЙСТВИеМ ИНГреДИеНТОВ, ВХО ДЯЩИХ в состав экстрактов. Кверцетин и ресвератрол в концентрациях, присутствующих в культуральной среде с добавлением экстрактов вина (1.10 мкг/мл и 163 мкг/мл) и кожицы винограда (2.31 мкг/мл и 1.52 мкг/мл)* не оказывали статистически значимого влияния на ц и КОЕ.
А нтиба ктериальна я а ктивность ра стительных экстрактов и отдельных полифенолов была отмечена ранее [Scalbert, 1991: Smith* Imlay. Mackie* 2003: Cushnie, Lamb* 2005; Obeidat et al., 2012: Subramanian et al.* 2014]. В ряде случаев было установлено, что механизм бактериостатического и бактерицидного действия полифенолов связан с продукцией активных форм кислорода (АФК) или окислительным повреждением мембран [Smith* Imlay. Mackie* 200.3: Subramanian et al., 2014]. АФК могут образовываться в результате аутоокисления полифенолов в аэробных условиях. В частности* ранее мы продемонстрировали, что богатые полифенолами растительные экстракты* а также танин и кверцетин, продуцируют перекись водорода и индуцируют антиоксидантные гены katG и sodA, кодирующие, соответственно, каталазу HPI и Мп-супероксиддисмутазу [Smirnova et al., 2009: Samoilova et al.. 2014]. Индукция антиоксидантных генов и возрастание активности ферментов, осуществляющих деструкцию АФК, способствуют повышению устойчивости к последующему окислительному стрессу [Smirnova et al.* 2009: Samoilova et al.? 2014]
Измерение активности ^-галактозидазы в штаммах* несущих слияние с геном lacZ, показало, что экстракты вина и кожицы винограда индуцируют экспрессию генов katG и sod А (табл. 2). Как отмечалось ранее, индукция этих генов может быть связана с генерацией активных форм кислорода при аутоокислении полифенолов и с активацией Fur регул она* в состав которого входит ген sodA, вследствие дефицита железа в результате его хелатирования [Smirnova et а!., 2009]. Экспрессия гена rpoS. кодирующего транскрипционный регулятор общего стрессового ответа* не изменялась в присутствии экстракта вина и достоверно снижалась экстрактом кожицы винограда (табл. 2). Аналогичным образом изменялась экспрессия подконтрольного регулятору RpoS гена katE, кодирующего каталазу НРП. Экспрессия гена sidAT входящего в состав SOS-регулона, который активируется в ответ на повреждение ДНК, не зависела от наличия экстрактов в среде.
Ранее было показано, что ресвератрол вызывает фрагментацию ДНК и дозозависимое повышение экспрессии гена smIA [Hwang. Lim* 2015]. В наших экспериментах концентрация ресвератрола в составе вина и кожицы винограда, а также соответствующая доза чистого препарата были недостаточны для того, чтобы вызвать SOS-ответ.
Повышение экспрессии антиоксидантных генов katG и sodA способствовало возрастанию устойчивости бактерий к последующему пероксидному стрессу. Индекс антиоксидантной активности (отношение удельной скорости роста бактерий, пре-добработанных экстрактом, к необработанным клеткам через 30 мин. после добавления Н2О2) составлял 5.3 и 5.5 для экстрактов вина и кожицы винограда, соответственно (табл. 2). Таким образом, исследуемые экстракты демонстрировали высокую антиоксидантную активность как в химиче- ском. так и в биологическом тесте*
Ранее мы и другие исследователи показали, что экстракты некоторых лекарственных растений способны модифицировать действие на Е. coil антибиотиков* имеющих различные внутриклеточные мишени (синтез ДНК, клеточной стенки и белка)* Направление и сила эффекта в значительной мере зависели от природы антибиотика и вида экстракта и были связаны с экспрессией антиокси дантных генов и хелатированием железа [Smirnova et aL 2012; Marathe et al** 2013; Samoilova et al** 2014]. В данной работе исследовалось влияние экстрактов вина и кожицы винограда на чувствительность Е. colt к антибиотикам ципрофлоксацину (синтез ДНК)* цефотаксим}7 (синтез клеточной стенки) и аминогликозидам канам ицину и стрептомицин}7 (синтез белка).
Таблица 2
Влияние экстрактов красного вина и кожицы винограда на экспрессию исследуемых генов и индекс АОА
|
Активность р-галактозидазы, ед. Миллера |
Контроль (ДМСО) |
Экстракт вина |
Экстракт кожицы винограда |
|
katGvJacZ |
7O±3 |
121±8*(1.73) |
139±11* (1.99) |
|
katEvJacZ |
417±31 |
409±28 (0.98) |
358±21 (0.86) |
|
rpoSvJacZ |
262±16 |
226±14 (0.86) |
202±9* (0.77) |
|
sodAOacZ |
55±2 |
94±5*(1.71) |
100±8* (1.82) |
|
stdAvJacZ |
56±5 |
47^2 (0*84) |
56^2(1.0) |
|
Индекс AOA |
1.0 |
5*3 |
5.5 |
Представлены данные через 50 мин, экспозиции с экстрактами, В скобках указан коэффициент относительно контроля, * Статистически достоверная разница с контролем (р<0.05).
Низкие дозы ципрофлоксацина (0.03 и 0.3 мкг/мл) не ингибировали рост в течение 30 мин. после воздействия. В этих условиях предобработка бактерий экстрактами вина и кожицы винограда снижала удельную скорость роста на 30-40%. Высокая доза ципрофлоксацина (3 мкг/мл), напротив, быстро ингибировала рост. При этом в культурах, предобработанных экстрактами, ц было на 30% выше, чем в тех, которые были обработаны только антибиотиком. В случае канамицина и стрептомицина экстракты вина и кожицы винограда усиливали ингибирование роста, вызванное антибиотиками (10 и 40 мкг/мл). Наиболее выраженный эффект предобработки экстрактами на рост бактерий наблюдался при экспозиции к цефотаксиму. Цефотаксим вызывал лизис клеток через 23 и 18 мин. после внесения в среду’ при дозе 5 и 10 мкг/мл, соответственно. Предобработка экстрактами предотвращала лизис в течение 70 мин. при концентрации цефотаксима 5 мкг/мл и существенно замедляла его при более высокой дозе антибиотика. Эти результаты хорошо согласуются с полученными нами ранее данными о влиянии богатых полифенолами экстрактов (зеленый и черный чай, толокнянка, брусника и др.) на рост Е. coh в присутствии антибиотиков с разным механизмом действия [Samoilova et al., 2014].
При экспозиции Е. coh ко всем изученным антибиотикам наблюдалось дозозависимое снижение числа КОЕ (рисунок). Предобработка бактерий экстрактами вина и кожицы винограда повышала значение КОЕ при действии низкой дозы ципрофлоксацина (0.03 мкг/мл) в 138 и 88 раз, соответственно. Концентрации кверцетина и ресвератрола, близкие к тем, которые содержатся в экстрактах, оказывали противоположное действие, повышая чувствительность бактерий к низкой дозе ципрофлоксацина. При более высоких концентрациях ципрофлоксацина протекторный эффект предобработок экстрактами был выражен значительно слабее.
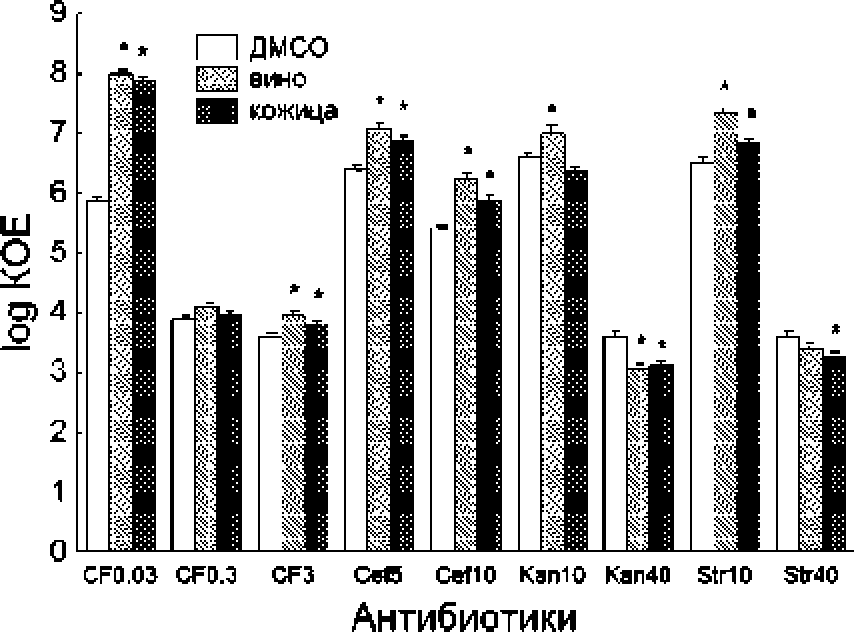
Влияние экстрактов вина и кожицы винограда на выживаемость Е. coh BW25113 при экспозиции к антибиотикам с разным механизмом действия,
CF0.03, CF0.3, CF3 - 0.03, 0.3 и 3 мкг/мл ципрофлоксацина, Се15, СеПО - 5 и 10 мкг/мл цефотакси-ма; Kani 0, Кап40 - 10 и 40 мкг/мл канамицина;
Str 10, Str40 - 10 и 40 мкг/мл стрептомицина. Представлены данные через 70 мин. после добавления антибиотика. ^Статистически достоверная разница по сравнению с действием антибиотика па клетки, не обработанные экстрактами (р<0.05)
Предобработка экстрактами в 3-6 раз повышала КОЕ при действии обеих доз цефотаксима, что хорошо согласуется со способностью экстрактов ингибировать лизис, индуцированный этим анти- биотиком. Напротив, экстракты вина и кожицы винограда в 2-4 раза усиливали бактерицидную активность высоких доз канамицина и стрептомицина (рисунок).
Аналогичным образом на чувствительность к канамицину и стрептомицину" влияли кверцетин и ресвератрол. Данные, полученные в настоящей работе, хорошо согласуются с результатами наших предшествующих исследований, показавших, что экстракты зеленого и черного чая. толокнянки и брусники ослабляют бактерицидный эффект ципрофлоксацина и ампициллина, но повышают восприимчивость к канамицину. Эффекты, вызываемые экстрактами, были тесно связаны с суммарным уровнем полифено лов и могли быть обусловлены влиянием на степень окислительного стресса в присутствии антибиотиков. Сходство модифицирующего действия различных экстрактов на чувствительность к антибиотикам может свидетельствовать об общности факторов и механизмов, ведущих к такой модификации.
Недавно мы показали [Смирнова и др.. 2016]. что скорость роста бактерий может быть основным фактором, определяющим их чувствительность к ципрофлоксацину и ампициллину, независимо от условий, влияющих на рост. Экстракты вина и кожицы винограда вызывали 30%-ное ингибирование скорости роста, которое могло быть следствием ирооксидантной и хелатирующей активности полифенолов, входящих в их состав. Такое снижение скорости роста может быть причиной протекторного действия экстрактов при экспозиции бактерий к ципрофлоксацину и цефотаксиму, который. как и ампициллин. ОТНОСИТСЯ к классу" 0-лактамов. Кроме того, показано, что действие антибиотиков на уровне первичных мишеней влечет за собой неспецифические метаболические изменения. способствующие окислительному повреждению всех типов макромолекул в клетке (ДНК. белки, липиды) [Belenky el al.. 2015]. Предобработка экстрактами изменяет редокс-ситуацию внутри клеток и стимулирует экспрессию антиоксидантных генов kaiG и sodA. что также может вносить вклад в модуляцию восприимчивости к антибиотикам. Поскольку исследуемые антибиотики имеют различные первичные внутриклеточные мишени, модулирующий эффект, производимый экстрактами, может сильно варьировать в зависимости от типа антибиотика, ВПЛОТЬ до противоположного воздействия у аминогликозидов по сравнению с хинолонами и 0-лактамами.
Внутриклеточной мишенью фторхинолона ципрофлоксацина являются бактериальные топоизомеразы. Хинолоны связываются с комплексом ДНК-фермент, препятствуя восстановлению разрывов в молекуле ДНК, что приводит к фрагментации хромосом и. в конечном итоге, к гибели кле ток [Drlica et al., 2008]. Экспозиция к ципрофлоксацину7, как и к другим агентам, повреждающим ДНК, индуцирует SOS-ответ, контролирующий репарацию ДНК и филаментацию клеток [Piddock. Wise, 1987]. В наших экспериментах ципрофлоксацин индуцировал экспрессию принадлежащего к SOS-регулону гена suLA* Максимальный уровень экспрессии наблюдался при концентрации ципрофлоксацина 0.3 мкг/мл. Предобработка экстрактами вина и кожицы винограда значительно ингибировала экспрессию sulA при этой концентрации ципрофлоксацина, свидетельствуя о снижении степени повреждения ДНК. Необходимы дальнейшие исследования для точного определения сайтов, на которые влияют компоненты экстрактов при действии других антибиотиков.
Заключение
Полученные в работе данные показывают, что предобработка клеток Е. сой экстрактами вина и кожицы винограда снижает чувствительность бактерий к ципрофлоксацину и цефотаксиму, но повышает восприимчивость к высоким дозам канамицина и стрептомицина. Модифицирующий эффект может быть связан с суммарным действием полифенолов в составе экстрактов. Воздействие экстрактов на клетки осуществляется, по-видимому. путем их влияния на скорость роста бактерий, редокс-сшуацию и уровень индукции антиоксидантных генов. В случае ципрофлокса цина. предобра ботка экстракта м и может снижать степень повреждения ДНК Модулирующее влияние продуктов, богатых полифенолами, на бактерицидную активность различных антибиотиков должно учитываться при антибактериальной терапии.
Исследования выполнены при финансовой поддержке грантов РФФИ №14-04-9603 L 16-04-00762 и Программы УрО РАН №15-4-4-16.
Список литературы Модифицирующее действие экстрактов кожицы винограда и красного вина на чувствительность бактерий Escherichia coli к различным антибиотикам
- Смирнова Г.В. и др. Роль тиоловых редокс-систем при ответе бактерий Escherichia coli на стрессорные воздействия температур и антибиотиков//Микробиология. 2016. Т. 85(1). С. 1-11
- Belenky P. et al. Bactericidal antibiotics induce toxic metabolic perturbations that lead to cellular damage//Cell Rep. 2015. Vol. 13. P. 968-980
- Crozier A., Jaganath I.B., Clifford M.N. Dietary phenolics: chemistry, bioavailability and effects on health//Nat. Prod. Rep. 2009. Vol. 26. P. 10011043
- Cushnie T.P.T., Lamb A.J. Antimicrobial activity of flavonoids//Int. J. Antimicrob. Agents 2005. Vol. 26. P. 343-356
- Drlica K. et al. Quinolone-mediated bacterial death//Antimicrob. Agents Chemother. 2008. Vol. 52. P. 385-392.
- Eberhardt M.V., Jeffery E.H. Perspective. When dietary antioxidants perturb the thiol redox//J. Sci. Food Agric. 2006. Vol. 86. P. 1996-1998.
- Hwang D., Lim Y.H. Resveratrol antibacterial activity against Escherichia coli is mediated by Z-ring formation inhibition via suppression of FtsZ expression//Sci. Rep. 2015. Vol. 5. P. 10029
- Kim H-J. et al. Evalution of antioxidant activity of Vetiver (Vetiveria zizanioides L.) oil and identification of its antioxidant constituents//J. Agric. Food Chem. 2005. Vol. 53. P. 7691-7695
- Marathe S.A. et al. Curcumin reduces the antimicrobial activity of ciprofloxacin against Salmonella typhimurium and Salmonella typhi//J. Antimicrob. Chemother. 2013. Vol. 68. P. 139-152
- Miller J.H. Experiments in molecular genetics. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. 1972
- Obeidat M. et al. Antimicrobial activity of crude extracts of some plant leaves//Res. J. Microbiol. 2012. Vol. 7. P. 59-67
- Perron N.R., Brumaghim J.L. A review of the anti-oxidant mechanisms of polyphenol compounds related to iron binding//Cell Biochem. Biophys. 2009. Vol. 53. P. 75-100
- Piddock L.J.V., Wise R. Induction of the SOS response in Escherichia coli by 4-quinolone antimicrobial agents//FEMS Microbiol. Lett. 1987. Vol. 41. P. 289-294
- Rice-Evans C.A. et al. The relative antioxidant activities of plant-derived polyphenolic flavonoids//Free Radic. Res. 1995. Vol. 22. P. 375-383
- Samoilova Z. et al. Medicinal plant extracts variously modulate susceptibility of Escherichia coli to different antibiotics//Microb. Res. 2014. Vol. 169. P. 307-317
- Scalbert A. Antimicrobial properties of tannins//Phytochemistry. 1991. Vol. 30. P. 3875-3883
- Shyur L-F. et al. Antioxidant properties of extracts from medicinal plants popularly used in Taiwan//Inter. J. Appl. Sci. Eng. Technol. 2005. Vol. 3. P. 195-202
- Smirnova G. V. et al Influence of polyphenols on Es-cherichia coli resistance to oxidative stress.//Free Radic. Biol. Med. 2009. Vol. 46. P. 759768
- Smirnova G. et al. Influence of plant polyphenols and medicinal plant extracts on antibiotic susceptibility of Escherichia coli//J. Appl. Microbiol. 2012. Vol. 113. P. 192-199
- Smith A.H., Imlay J.A., Mackie R.I. Increasing the oxidative stress response allows Escherichia coli to overcome inhibitory effects of condensed tannins//Appl. Environ. Microbiol. 2003. Vol. 69. P. 3406-3411
- Subramanian M. et al. Resveratrol induced inhibition of Escherichia coli proceeds via membrane oxidation and independent of diffusible reactive oxygen species generation//Redox biology. 2014. Vol. 2. P. 865-872
- Tang S.Y., Halliwell B. Medicinal plants and antioxi-dants: What do we learn from cell culture and Caenorhabditis elegans studies?//Biochem. Biophys. Res. Commun. 2010. Vol. 394. P. 1-5
- Tao K. et al. Molecular cloning and nucleotide sequencing of oxyR, the positive regulatory gene of a regulon for an adaptive response to oxidative stress in Escherichia coli: homologies between OxyR protein and a family of bacterial activator proteins//Mol. Gen. Genet. 1989. Vol. 218. P. 371-376
- Wu L-C. et al. Antioxidant and antiproliferative activities of red pitaya//Food Chem. 2006. Vol. 95. P. 319-327
