Модулирующее влияние ГАМК и габазина на респираторные реакции в условиях электростимуляции большого ядра срединного шва

Автор: Орлова Анастасия Олеговна, Инюшкин Алексей Николаевич
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Физиология
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
Цель работы. Изучить реакции паттерна дыхания и биоэлектрической активности инспира-торных мышц у крыс при электростимуляции рострального отдела большого ядра до и на фоне микроинъекции гамма-аминомасляной кислоты (ГАМК) (в первой серии эксперимента) и габази-на (во второй серии). Материалы и методы. Эксперименты проведены на 24 взрослых нелинейных крысах. Животных наркотизировали уретаном (1,5 г/кг внутрибрюшинно). Паттерн дыхания регистрировали методом спирографии. Одновременно с этим записывали биоэлектрическую активность диафрагмы и наружных межреберных мышц. Результаты. Зарегистрировано сокращение продолжительности вдоха и снижение амплитудных показателей респираторных реакций при электростимуляции большого ядра. Микроинъекции габазина в данную структуру также приводили к уменьшению длительности инспирации, дыхательного объема, биоэлектрической активности наружных межреберных мышц, к небольшому увеличению длительности экспирации. Микроинъекции ГАМК, наоборот, вызывали рост биоэлектрической активности интеркостальных мышц и минутного объема дыхания. При электростимуляции на фоне микроинъекции ГАМК максимальная амплитуда осцилляций в залпах активности инспираторных мышц также повышалась. При этом были зарегистрированы снижение частотных и увеличение амплитудных параметров внешнего дыхания. Электростимуляция большого ядра на фоне введения габазина также вызывала увеличение дыхательного объема, минутного объема дыхания, максимальной амплитуды осцилляций залповой активности диафрагмы и наружных межреберных мышц. Заключение. Полученные данные указывают на участие ГАМК в модулирующем респираторном влиянии нейронов большого ядра шва, реализующемся через ГАМКА-рецепторы.
Большое ядро срединного шва, гамма-аминомасляная кислота, габазин, регуляция дыхания
Короткий адрес: https://sciup.org/14113228
IDR: 14113228 | УДК: 612.751.3
Modulatory effects of GABA and gabazin on respiratory responses during electrostimulation of nucleus raphe magnus
Objective. Nucleus raphe magnus, preliminary consisting of serotonergic and GABAergic neurons, is involved in modulation of different respiratory responses. GABA, a neurotransmitter participating in res-piratory rhythmogenesis, is known to have inhibitory effects on respiratory parameters. Materials and Methods. The authors studied the responses of respiratory pattern and bioelectric activity of inspiratory muscles in rats to electrostimulation of the rostral nucleus raphe magnus before and during microinjections of GABA or gabazine. Experiments were conducted on adult rats anaesthetized with urethane. Results. A decrease both in inspiratory duration and in amplitude parameters during electrical stimulation of the nucleus raphe magnus was found. Gabazine microinjected into the same nucleus induced inhalation shortening, reduction in tidal volume, decrease in bioelectric activity of external intercostal muscles, and uptick in expiratory duration. Microinjections of GABA induced increase in bioelectric activity of external intercostal muscles and minute volume. During electric stimulation after microinjection of GABA, maximal amplitude of oscillations of inspiratory muscles also increased. Besides, breathing frequency reduction and amplitude increase were recorded. Electric stimulation after administration of gab-azine also caused increase of the tidal volume, minute volume, maximal amplitude of oscillations of bursting activity of the diaphragm and external intercostal muscles. Conclusion. The obtained data confirm the involvement of GABA via activation of GABAa receptors in modulation of respiration by neurons of nucleus raphe magnus.
Текст научной статьи Модулирующее влияние ГАМК и габазина на респираторные реакции в условиях электростимуляции большого ядра срединного шва
Введение. Атмосферный воздух представляет собой смесь газов: азота, кислорода, двуокиси углерода и инертных газов. Изменение процентного соотношения компонентов вдыхаемого воздуха, безусловно, оказывает влияние на состояние живого организма. Регуляция дыхания, в основе которой лежат нейронные механизмы, реализующиеся на уровне дыхательного центра, а также на основе взаимодействия дыхательного центра с различными нейронными образованиями, направлена на поддержание гомеостаза организма в различных условиях окружающей среды [1, 2]. К числу нейронных образований, активно взаимодействующих с дыхательным центром, относится большое ядро срединного шва, являющееся частью серотонинергической системы [3].
Данная структура локализована в стволе мозга на уровне ядра лицевого нерва и характеризуется анатомической и физиологической гетерогенностью [4, 5]: наряду с серотонинергическими нейронами, она включает в себя клетки, продуцирующие различные нейромедиаторы и модуляторы, такие как гамма-аминомасляная кислота (ГАМК), аминоук- сусная кислота, вещество P, тиролиберин, лейцин-энкефалин и метионин-энкефалин [6]. Тонкая координация активности тех или иных клеток большого ядра обеспечивается наличием ауторецепторов у серотонинергических нейронов, а также ГАМКергической интернейронной сетью, взаимодействующей с другими (в т.ч. не-ГАМКергическими) клетками и регулирующей их активность через ГАМКA- и ГАМКB-рецепторы [7–10].
Данные литературы свидетельствуют о том, что воздействие ГАМК на дыхательный центр приводит к торможению респираторной активности [11–13]. Известно также, что ГАМК участвует в механизмах формирования ритма дыхания [13–14]. Согласно данным некоторых авторов, ингибирующее влияние на параметры внешнего дыхания при электрическом или химическом раздражении большого ядра обусловлено преимущественным участием ГАМК, а не серотонина [15].
Цель исследования. Изучение реакции паттерна дыхания и биоэлектрической активности инспираторных мышц у крыс при электростимуляции (50 Гц, 15 В) рострального отдела большого ядра (координаты в мм относительно bregma: Р – 10,3; L – 0,0 ; V – 9,9) [16] до и на фоне микроинъекции ГАМК (10-5 М). С целью выяснения возможного участия ГАМК А -рецепторов в реализации этих изменений было изучено влияние локального введения специфического антагониста ГАМК А -рецепторов габазина в исследуемую область большого ядра на характер реакций паттерна внешнего дыхания и электрической активности инспираторных мышц при электростимуляции данной точки.
Материалы и методы. Эксперименты проведены на 24 взрослых нелинейных крысах обоих полов массой 180–250 г. Животных наркотизировали уретаном (1,5 г/кг внутрибрюшинно). Паттерн дыхания регистрировали методом спирографии. На полученных спирограммах оценивали дыхательный объем (VТ, мл), продолжительность инспираторной (TI, с) и экспираторной (TE, с) фаз цикла внешнего дыхания. Рассчитывали частоту дыхания (f=60/ТТ, мин-1) и минутный объем дыхания (VI=fVТ, мл/мин). Одновременно с паттерном дыхания регистрировали биоэлек- трическую активность диафрагмы и наружных межреберных мышц (VI–VIII межреберье) с правой стороны тела животных с помощью стальных игольчатых электродов биполярным способом. На полученных электромиограммах определяли максимальную амплитуду осцилляций (отн. ед.) в залпах активности инспираторных мышц. Инъекции ГАМК и габазина осуществляли микрошприцом МШ-1 через стеклянную пипетку, диаметр кончика которой составлял 20–30 мкм. Вещество растворяли ex tempore в искусственной цереброспинальной жидкости и вводили в объеме 0,5 мкл со скоростью около 0,1 мкл/с. В контрольных наблюдениях в ту же точку ствола мозга вводили искусственную цереброспинальную жидкость в объеме 0,5 мкл. Для электростимуляции большого ядра применяли биполярный концентрический электрод диаметром 0,02 мм. Использовали следующие параметры стимулирующего тока – 50 Гц, 15 В.
Полученные экспериментальные данные обрабатывали статистически с использованием теста ANOVA для повторных измерений, тестов Dunnett's и Tukey для сравнения величины исследуемых показателей с их исходным уровнем. Статистически значимыми считали изменения при р<0,05.
Результаты и обсуждение. В результате настоящего исследования установлено, что в условиях электростимуляции большого ядра продолжительность вдоха максимально сокращалась на 30 % (р<0,001). При этом отмечена тенденция к росту частоты дыхания и уменьшению глубины дыхания относительно исходного уровня. Максимальная амплитуда осцилляций в залпах активности диафрагмы уменьшалась на 13 % (р<0,001). Максимальная амплитуда осцилляций залпов активности наружных межреберных мышц уменьшалась на 15 % (р<0,05).
Введение 10-5 М ГАМК в большое ядро приводило к усилению биоэлектрической активности только наружных межреберных мышц на 37 % (р < 0,01; рис. 1). Имела место и слабая (статистически незначимая) тенденция к повышению минутного объема дыхания. При этом частотные характеристики внешнего дыхания оставались неизменными.
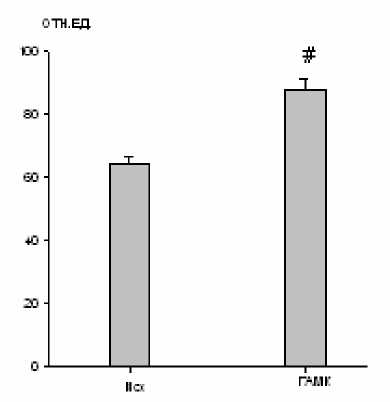
Рис. 1. Изменение максимальной амплитуды осцилляций залповой активности наружных межреберных мышц после микроинъекции ГАМК (10-5 М) в большое ядро (# – р<0,01)
Через 10 мин после микроинъекции ГАМК осуществляли электростимуляцию большого ядра, в результате чего происходило урежение частотных и увеличение амплитудных параметров дыхания. Частота дыхания снижалась на 8 % (р < 0,05; рис. 2). Минутный объем дыхания увеличивался на 6 %
(р < 0,05; рис. 2). Одновременно с этим происходило усиление биоэлектрической активности инспираторных мышц, проявившееся, однако, в увеличении амплитуды осцилляций только наружных межреберных мышц на 24 % (р < 0,05; рис. 2).
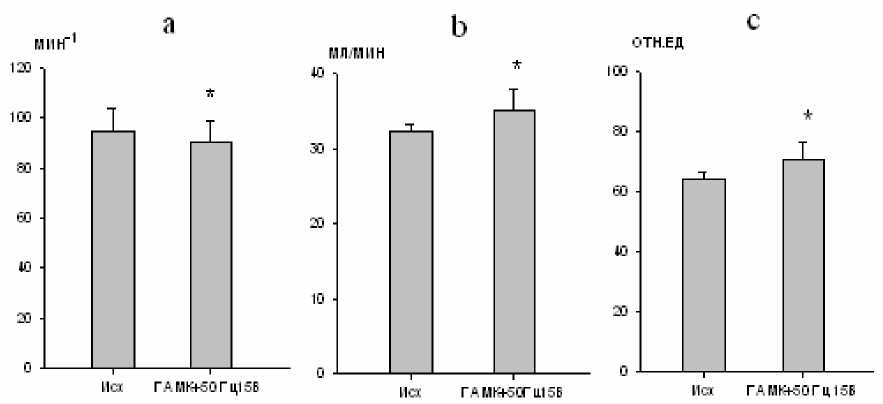
Рис. 2. Изменение частоты дыхания (a), минутного объема дыхания (b)
и максимальной амплитуды осцилляций залповой активности наружных межреберных мышц (c) при электростимуляции большого ядра (частота тока 50 Гц, сила стимула 15 В)
на фоне ГАМК (10-5 М)
( * - р<0,05)
Нами установлено, что микроинъекции 10-5 М габазина в большое ядро вызывали закономерные изменения ряда дыхательных показателей. Продолжительность экспирации увеличивалась на 10 % (р<0,05; рис. 3), а дли- тельность инспирации уменьшалась на 14 % (р<0,05; рис. 3). Дыхательный объем также уменьшался на 10 % (р<0,05; рис. 4). Одновременно изменялась и биоэлектрическая активность инспираторных мышц, причем мак- симальная амплитуда осцилляций наружных межреберных мышц снижалась на 46 % (р<0,01; рис. 4), тогда как статистически зна- чимых реакций аналогичного показателя активности диафрагмы не зарегистрировано.
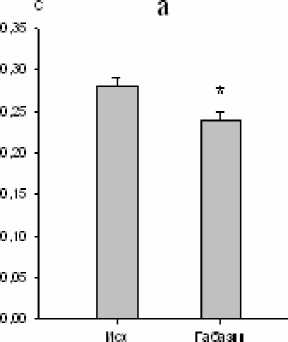
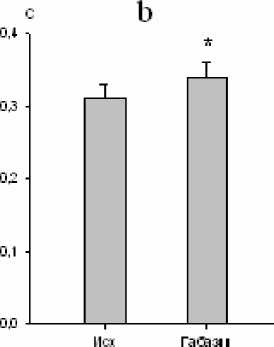
Рис. 3. Изменение времени инспирации (a) и времени экспирации (b) после микроинъекции габазина (10-5 М) в большое ядро ( * – р<0,05)
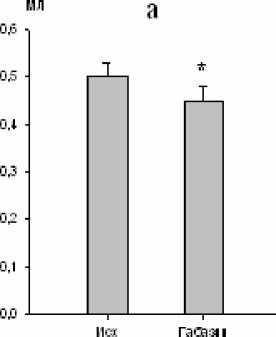
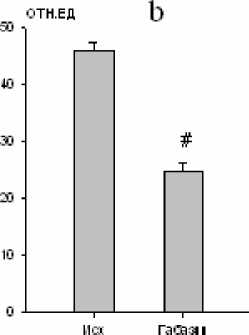
Рис. 4. Изменение дыхательного объема (a) и максимальной амплитуды осцилляций залповой активности наружных межреберных мышц (b) после микроинъекции габазина (10-5 М) в большое ядро ( * – р<0,05; # – р<0,01)
При электростимуляции большого ядра на фоне микроинъекции габазина (через 10 мин) имела место тенденция к увеличению продолжительности инспирации и снижению частоты дыхания, тогда как продолжительность экспирации оставалась стабильной. Дыхательный объем и минутный объем дыхания увеличивались на 26 (р < 0,01; рис. 5) и 18 % (р < 0,05; рис. 5) соответственно. Одновременно с этим был отмечен рост биоэлектрической активности инспираторных мышц.
Максимальная амплитуда осцилляций наружных межреберных мышц увеличивалась на 52 % (р < 0,01; рис. 6), а диафрагмы – на 10 % (р < 0,01; рис. 6).
Микроинъекции искусственной цереброспинальной жидкости в большое ядро в контрольных наблюдениях не вызывали статистически значимых изменений исследуемых параметров паттерна внешнего дыхания и биоэлектрической активности инспираторных мышц (р>0,05).
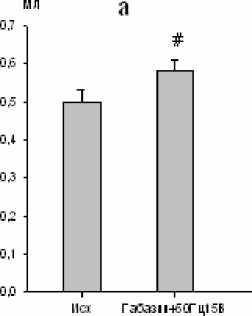
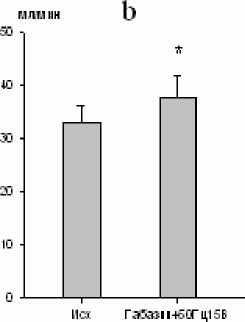
Рис. 5. Изменение дыхательного объема (a) и минутного объема дыхания (b) при электростимуляции большого ядра (частота тока 50 Гц, сила стимула 15 В) на фоне габазина (10-5 М) ( * - р<0,05; # - р<0,01)
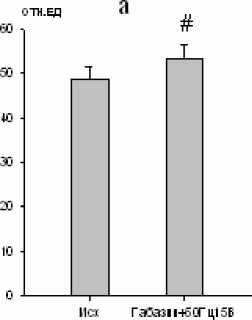
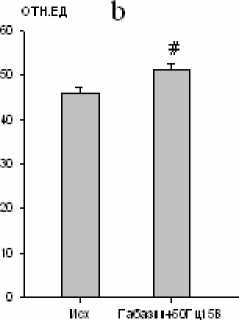
Рис. 6. Изменение максимальной амплитуды осцилляций залповой активности наружных межреберных мышц (a) и диафрагмы (b)
при электростимуляции большого ядра (частота тока 50 Гц, сила стимула 15 В) на фоне габазина (10-5 М) (# – р<0,01)
Таким образом, электростимуляция большого ядра приводила к сокращению времени инспирации и снижению амплитудных показателей – уменьшению биоэлектрической активности дыхательных мышц. Такую же направленность респираторных реакций мы зарегистрировали при введении габа-зина (ГАМКА-блокатора) в большое ядро. Возможно, при электростимуляции происходит возбуждение серотонинергических нейронов, которые ингибируют ГАМКергиче-ские клетки, или активизируются ГАМКВ-рецепторы, которые тормозят выход ГАМК. Микроинъекции ГАМК, наоборот, вызывали увеличение биоэлектрической активности интеркостальных мышц и минутного объема дыхания. Это может быть свидетельством того, что в данных условиях активировались ГАМКА-рецепторы, способствующие высвобождению ГАМК из локально расположенных аксонных терминалей [7, 15]. В условиях электростимуляции на фоне микроинъекции ГАМК максимальная амплитуда осцилляций в залпах активности инспираторных мышц также повышалась, но выраженность биоэлектрической активности наружных межреберных мышц стала меньше (24 % против 37 %), что, возможно, связано с возбуждением серотонинергических нейронов и их влиянием на ГАМКергические клетки. При этом было зарегистрировано снижение частотных и увеличение амплитудных параметров внешнего дыхания. Электростимуляция большого ядра на фоне введения габазина также вызывала увеличение дыхательного объема, минутного объема дыхания, максимальной амплитуды осцилляций залповой активности диафрагмы и наружных межреберных мышц. Можно предполагать, что при электростимуляции помимо серотониергиче-ских нейронов возбуждаются и ГАМКерги-ческие, что приводит к повышению амплитудных параметров.
Заключение. Полученные данные указывают на участие ГАМК в модулирующем респираторном влиянии нейронов большого ядра шва, реализующемся через ГАМК А -ре-цепторы..
Список литературы Модулирующее влияние ГАМК и габазина на респираторные реакции в условиях электростимуляции большого ядра срединного шва
- Nichols N.L., Johnson R.A., Satriotomo I., Mitchell G.S. Neither serotonin nor adenosine-dependent mechanisms preserve ventilatory capacity in ALS rats. Respir. Physiol. Neurobiol. 2014; 197: 19-28.
- Subramanian H.H., Holstege G. The midbrain periaqueductal gray changes the eupneic respiratory rhythm into a breathing pattern necessary for survival of the individual and of the species. Prog. Brain. Res. 2014; 212: 351-384.
- Pilowsky P.M. Peptides, serotonin, and breathing: the role of the raphe in the control of respiration. Prog. Brain. Res. 2014; 20: 169-189.
- Bagdy E., Kiraly I., Harsing L.G. Reciprocal innervation between serotonergic and GABAergic neurons in raphe nuclei of the rat. Neurochemical Research. 2000; 25 (11): 1465-1473.
- Harsing L.G. The Pharmacology of the neurochemical transmission in the midbrain raphe nuclei of the rat. Current Neuropharmacology. 2006; 4: 313-339.
- Serrats J., Mengod G., Corte's R. Expression of serotonin 5-HT2C receptors in GABAergic cells of the anterior raphe nuclei J. Chemical Neuroanatomy. 2005; 29: 83-91.
- Инюшкин А.Н., Меркулова Н.А., Орлова А.О., Инюшкина Е.М. Локальная ГАМКергическая модуляция активности серотонергических нейронов большого ядра шва. Российский физиологический журнал им. И.М. Сеченова. 2009; 95 (7): 750-761.
- Bie В., Fields H.L., Williams J.T., Pan Z.Z. Roles of i-t, -and Q^-adrenoceptors in the nucleus raphe magnus in opioid analgesia and opioid abstinence-induced hyperalgesia. Neuroscience. 2003; 23 (21): 7950-7957.
- Перфилова В.Н., Тюренков И.Н. ГАМКВ-рецепторы: строение и функции. Экспериментальная и клиническая фармакология. 2010; 73 (11): 44-48.
- Ковалев А.М., Ведясова О.А., Сафонов В.А., Тараканов И.А. Роль ГАМКВ-рецепторов в регуляции паттерна дыхания структурами вентральной респираторной группы. Вестник Удмуртского университета. 2011; 2: 110-115.
- Александрова Н.П., Александров В.Г., Иванова Т.Г. Влияние гамма-аминомасляной кислоты на инспираторно-тормозящий рефлекс Геринга-Брейера. Рос. физиол. журн. им. И.М. Сеченова. 2008; 94 (12): 1356-1364.
- Татаринцева Д.С., Маньшина Н.Г., Ведясова О.А. Реакции дыхания при микроиньекциях ГАМК и пинициллина в область дорсальной респираторной группы у крыс. Ульяновский медико -биологический журнал. 2013; 1: 109-115.
- Маньшина Н.Г., Ведясова О.А., Татаринцева Д.С. Изменение паттерна дыхания при микроиньекциях бикукулина в комплекс Бетцингера и комплекс пре-Бетцингера у крыс. Вестник Тверского государственного университета. Биология и экология. 2013; 29: 185-194.
- Маньшина Н.Г., Ведясова О.А. Роль ГАМКА-рецепторов комплекса Бетцингера в регуляции дыхания у крыс. Ульяновский медико-биологический журнал. 2014; 1: 138-144.
- Cao Y., Matsuyama K., Fujito Y., Aoki M. Involvement of medullary GABAergic and serotonergic raphe neurons in respiratory control: electrophysiological and immunohistochemical studies in rats. Neuroscience Research. 2006; 56 (3): 322-331.
- Paxinos G., Watson. C. The rat brain in stereotaxic coordinates. San Diego: Academic; 1998. 474.
