Молекулярно-транспортные расстояния в обитаемых пространствах в иерархически организованной фитосистеме

Автор: Празукин Александр Васильевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
По отношению к фитосистемам телесного и надтелесного уровня организации рассматривается морфофункциональный показатель: отношение объёма обитаемого пространства растения, в любом его иерархическом статусе, к его граничной поверхности (L = V/S, см). L интерпретируется как среднее расстояние между всеми точками на поверхности S и всеми точками в объёме ограниченного пространства V. Предлагаемый подход позволяет однотипно рассматривать объекты разного уровня организации и происхождения, и сравнивать их в общем метрическом поле. Показано, что L стабилизируется в онтогенетических и размерных рядах объектов телесного и надтелесного уровней организации за счет формирования оптимальной для этого геометрической формы обитаемого пространства и через создание множеств из физических отдельностей со свойственной им структурой и функцией.
Обитаемое пространство, жизненное пространство, пространственная иерархия, объём, площадь поверхности, архитектоника слоевища, фитосистема, местофункционирование
Короткий адрес: https://sciup.org/146279504
IDR: 146279504 | УДК: 582.26:
Molecular-transport distances in inhabited spaces of a hierarchially organized phytosystem
Here we consider the morphofunctional index: the ratio of the volume of the habitable space of the plant to its boundary surface (L = V/S, cm). L is interpreted as the average distance between all points on the surface S and all points in the volume of the bounded space V. The proposed approach allows to consider and compare objects of different levels of organization and origin. It is shown that L is stabilized in ontogenetic and dimensional series of objects at the organizational and ecological levels of the organization by changing the geometric shape of the habitable space and the structure of the phytocenosis.
Текст научной статьи Молекулярно-транспортные расстояния в обитаемых пространствах в иерархически организованной фитосистеме
Введение. Фитосистема в границах отдельно взятого водоёма представляет собой иерархически организованную систему, функционирующую в структурированном пространстве, в отдельных пространственных отсеках, связанных потоками веществ и энергии, проходящих через их граничные поверхности (рис.1). Условно первый отсек - отсек в границах телесного пространства (Gn), это отдельное растение с характерными для него величинами геометрических характеристик: длины (Zn), площади поверхности (Sn), объёма (Уп). Второй - кроновые пространства отдельно взятых растений (Vn+i), это пространство, охваченное разветвленной частью тела растения (в границах Gn+i) и оно включает в себя как объем пространства, занятого телом растения (Vn), так и объем пространства воды между элементами растения (объём ближайшего функционального пространства, Убфп): Vn+l=Vn + Убфп. Третий — пространство (Уп+з) в границах растительного покрова (растительного полога) (Gn+2) и четвертый - пространство в границах отдельного водоёма (Gn+з) с характерными значениями объёма водной массы (Уп+з) и площади внешних граничных поверхностей (площади поверхности зеркала, 8зерк и площади поверхности дна, 8 дно).
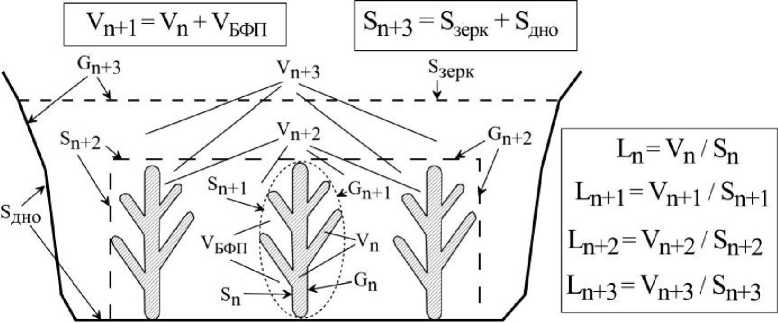
Рис. 1 . Модель иерархически организованной фитосистемы в ограниченном экологическом пространстве, в пространстве водоёма: Gn, Vn , Sn — внешняя граница, объём и площадь поверхности телесного пространства отдельного растения; Gn+i, Vn+i , Sn+i - внешняя граница, объём и площадь поверхности кронового пространства; Gn+2, Vn+2 , Sn+2- внешняя граница, объём и площадь поверхности растительного покрова (растительного полога); Gn+3, Vn+з, Sn+3 - внешняя граница, объём и площадь граничной поверхности водоёма; 8зерк, 8ДН0-площадь зеркала и дна водоёма; Убфп — объём ближайшего функционального пространства; длина молекулярно-транспортного расстояния в границах телесного (Ln) и кронового (Ln+i) пространств, растительного полога (Ln+2) и водоёма (Ьп+з).
В работах (Хайлов, 2001; Хайлов и др. 2001, 2002, 2009; Празукин, 2000, 2015; Празукин и др, 2003) предлагается и обсуждается применительно к конкретным ситуациям кусочно-линейная модель молекулярного пути (L) в ограниченном экологическом пространстве. Согласно высказанным представлениям, длина метаболического пути (L) определяется соотношением: У/S, где: V объём объекта, а 8 его площадь граничной (внешней) поверхности. В таком случае, длина метаболического пути в ограниченном объёме пространства интерпретируется как среднее расстояние между всеми точками на поверхности 8 и всеми точками в объёме ограниченного пространства V (Празукин, 2000; Хайлов, 2001; Хайлов и др. 2009).
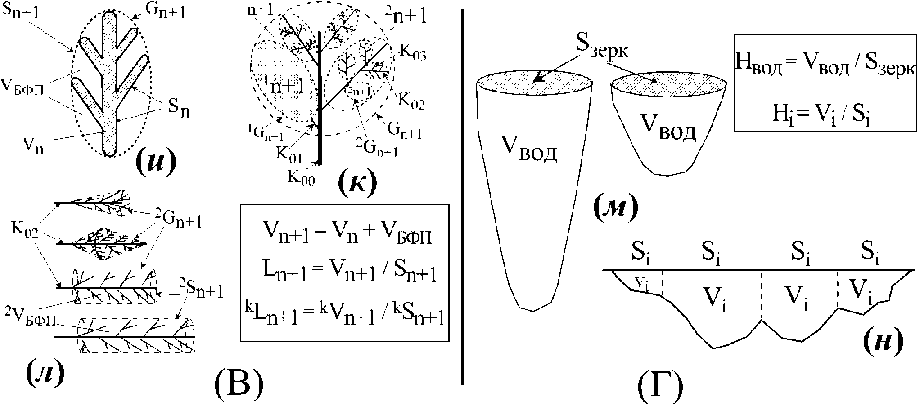
К-04 / к()1
К02 1 .
.к к°з к00
(я)
(б)
(г) - -''
Кт к„з
(б) "^
к02
(A)
Lp-Vn/ Sn OkLn— ®kVn / Ok^ »l^=£OkVn/SOksn
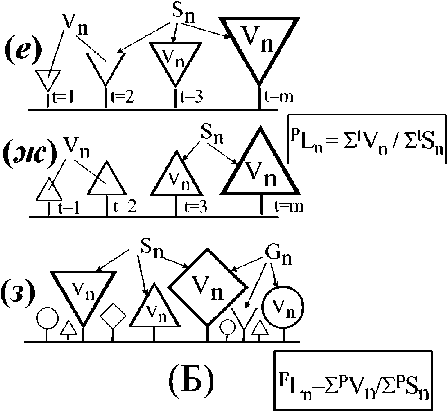
Рис. 2 . A — телесное пространство многоклеточной водоросли, организованное множеством её морфологических структур, а — растение в границах телесного пространства; схема иерархической организации вертикального «побега» цистозир (Cystoseira crinita, С. barbata, б) и его ветвей первого (Ki, г) и второго (К2, д) порядка; в — размерный ряд осевых структур цистозиры, соответственно, их телесных пространств. Б - телесное пространство, представленное множеством телесных пространств отдельных растений, организованных в популяции (е, ж) и в сообщества (з). В — кроновые и субкроновые пространства многоклеточных водорослей, и — растение в границах кронового пространства; к — схема иерархической организации кронового пространства вертикального «побега» цистозир (Cystoseira crinita, С. barbata)', л - ветви цистозиры второго порядка с разной формой субкронового пространства, меняющегося в её онтогенезе. Г - геоморфологические образы водоёмов, м — водоёмы с разным соотношением их стационарного объёма (Увод) и площади поверхности зеркала (S3epK); н — отдельные участки водоёма с харектерными для них значениями площади поверхности зеркала (Si), объёма воды (У;) и средней глубины (Li): Gn, Vn, Sn и Ln — внешняя граница, объём, площадь поверхности телесного пространства отдельного растения и длина молекулярно-транспортного расстояния в его границах; Коо, Koi - Кц4 ствол, осевые структуры цистозиры первого - четвертого порядка; okVn, okSn, и OkLn - объём, площадь поверхности телесного пространства осевых структур цистозиры к порядка и длина молекулярно-транспортного расстояния в их границах; УокУп и £okSn- суммарный объём и суммарная площадь поверхности телесного пространства всех осевых структур к порядка отдельного растения или ветви соответствующего порядка ветвления и их средняя длина молекулярно-транспортного расстояния (-,kLn); kVn, kSn, kLn - объём, площадь поверхности и длина молекулярно-транспортного расстояния телесного пространства ветвей цистозиры к порядка; У' Уп и £'Sn - суммарный объём и суммарная площадь поверхности всех растений популяции соответствующего вида t; £РУП и £pSn - суммарный объём и суммарная площадь поверхности всех видов растений фитоценоза F; средняя для популяции (pLn) и для фитосообщества (FLn) длина молекулярно-транспортного расстояния телесного пространства растений; Gn+i, Vn+i, Sn+i, Ln+i - внешняя граница, объём, площадь поверхности и длина молекулярно-транспортного расстояния кронового пространства; kGn+i, kVn+i, kSn+i, kLn+i — внешняя граница, объём, площадь поверхности и длина молекулярно-транспортного расстояния субкроновых пространств ветвей к порядка; объём водного пространства (Увод), площадь поверхности зеркала (8зерк), средняя глубина (Нвод) водоёма и, соответственно, эти же характеристики его отдельного участка (V;, Si, Hi); Убфп- объём ближайшего функционального пространства.
В настоящей статье вместо термина «длина метаболического пути» будем использовать термин «длина молекулярно-транспортного расстояния». С формальной точки зрения, L - это средняя длина линейного (диффузионного) пути молекул вещества, проходящих от внешней поверхности объекта во все точки в его объёме. Иными словами, L - это длина среднего минимально возможного молекулярнотранспортного расстояния в толщу, в объем объекта (поступление пищевых веществ) или обратно (выделение наружу молекулярных продуктов жизнедеятельности).
Чем меньше молекулярно-транспортное расстояние, равно и светового пути, тем меньше потерь и тем больше доступно растению вещество и энергия, а значит, что фитосистемы любого уровня организации должны формировать свою геометрическую структуру, оптимизируя её по длине метаболического пути.
Рассмотрим примеры регуляции названной характеристики в размерных и онтогенетических рядах объектов телесного и надтелесного уровня организации биогенного происхождения, а также расмотрим, как в размерном ряду водоемов (геогенный уровень организации) изменяется их средняя глубина. В последнем случае может идти речь об «управлении» геоморфологии водоемов и, соответственно, их водообменом, тепловым режимом, химией и жизнью в водной среде через рельеф, занятый ложем водоёма и окрестной местности (Хайлов и др., 2009).
Объекты и их параметрические характеристики
Вначале рассмотрим две группы объектов, они же два уровня пространственной организации фитосистем.
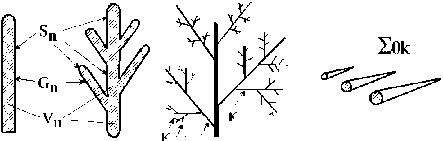
Телесный уровень организации. Объекты этого класса представлены множеством биологических отдельностей с физически организованной поверхностью. Это отдельное растение (рис. 2 а, б) с характерными размерами его тела (Vn) и внешней поверхностью (Sn), это различные структурные элементы тела растения (рис. 2 в - д), это и множество тел одного вида растений (популяция отдельного вида растений, рис. 2 е, ж) или множество тел разных видов растений (сообщество видов, рис. 2 з\ Во всех этих случаях идет речь о телесном пространстве (Vn), ограниченном физической поверхностью (Sn). В морфологической структуре многоклеточных водорослей с осевой структурой таллома мы выделяем две группы объектов: осевые структуры (Ко, Koi -Ко4, рис. 2 б, в) и ветви (рис. 2 г, д) соответствующего порядка ветвления. «Ветвь» рассматривается нами как сложноразветвленный элемент слоевища (иерархически организованный ансамбль осевых структур), обладающий собственной морфофункциональной структурой, закономерно меняющейся во времени. В таком случае, ветвь первого порядка (рис. 2 г) - это ось первого порядка (Koi) со всеми осевыми структурами последующих порядков ветвления (К02- К04), а ветвь второго порядка (рис. Id) - это ось второго порядка (К02) со всеми осевыми структурами последующих порядков ветвления (Коз - К04).
Алгоритмы расчета величин Уп и Sn на уровне отдельного растения и его структурных элементов, на уровне популяции и на уровне фитосообщества подробно описаны в работах (Хайлов, Фирсов, 1985; Хайлов и др., 2004; Празукин, 2015, 2016), а варианты расчета длины молекулярно-транспортного расстояния у вышеназванных объектов представлены на рис. 2 А, Б и в работах (Хайлов и др., 2009; Празукин и др, 2003; Празукин, 2015).
Объекты надтелесного уровня организации. Это объекты в границах кронового пространства Gn+i (рис. 1, 2 и, к) и в границах растительного полога Gn+2 (рис. 1). В обоих случаях их внешняя граничная поверхность не является физической поверхностью и проходит по внешнему контуру окончаний структурных элементов растения, как это показано пунктирной линией на рис. 2 и, к. Аналогичным образом выделяются и субкроновые пространства ветвей соответствующего порядка ветвления (рис. 2 к, л). Иерархическое членение надтелесного пространства как водных, так и наземных растений (рис. 2 к) следует иерархии тела растения (рис. 2 б). У цистозир (Cystoseira crinita (Desf.) Bory, 1832, C. barbata (Good. Et Wood.) Ag., 1821) с хорошей определенностью вычленяются субкроновые пространства ветвей первого и второго порядка (рис. 2 к), форма и размер которых закономерно меняются в онтогенезе (рис. 2 л) и в разной степени зависят от условий подвижности воды.
Методические приёмы, используемые для измерения параметрических характеристик кроновых и субкроновых пространств (включая определение объёма пространства и площади граничной поверхности) у разных видов водных и наземных растений представлены в работах (Празукин, 2005, 2007, 2015, 2017), а алгоритмы расчета длины молекулярно-транспортного расстояния - на рис. 2 В.
Водоёмы и их геоморфологические параметры. Основными геоморфологическими параметрами водоёмов считаются: объём водного пространства (Увод), площадь поверхности зеркала (8зерк) и дна (8ДН0) водоёма, его средняя глубина (Н) и др. Отношение объёма водного пространства к его всей граничной поверхности (80бщая = 8зерк + 8 дно): ЕВод=Увод/8 общая рассматривается как метаболическая глубина водоёма (Хайлов и др. 2009), тогда как его средняя глубина (Нвод) определяется соотношением: Нвод = Увод/8зерк. В рамках этой статьи рассматривается показатель средней глубины водоёма и как этот показатель соотносится с величиной объёма водного пространства.
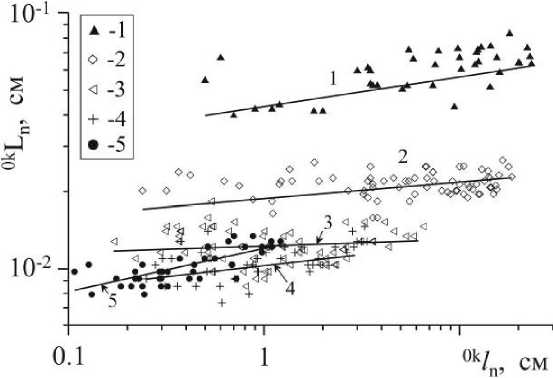
Рис. 3 . Соотношение индивидуальной длины (ок/п) и длины молекулярнотранспортного расстояния (okLn) в границах телесного пространства у осевых структур С. crinita нулевого - четвертого (1-5) порядков ветвления
Результаты и обсуждения
Фитосистемы телесного уровня организации. У водорослей диффузионные процессы играют определяющую роль в переносе веществ, что накладывает определенные ограничения на размеры их тела и на функциональную дифференциацию тканей по вертикальной составляющей от граничной поверхности тела. Так, у сложноорганизованных водорослей, к каким относятся ламинариевые и фукусовые, слой клеток, содержащих хлоропласты, состоит чаще из 1 -5 и до 10 рядов клеток и располагается непосредственно у внешней границы тела растения (Зинова, 1976; Калугина-Гутник, 1975; Камнев и др., 1988; Камнев, 1989) и здесь сосредоточена основная масса хлоропластов. Доля этого слоя в общем объеме осевых структур цистозир (С. crinita, С. barbata?) составляет 9 - 19% (рассчитана по рисункам, Калугина-Гутник, 1975, стр. 164, 165, 171). Последнее говорит о том, что между интенсивностью фотосинтеза, рассчитанного на единицу объема (цу), и Ln может существовать обратно пропорциональная зависимость, а это значит, что рост таллома в толщину (увеличение диаметра у осевых структур и толщины у пластинчатых слоевищ) растениям метаболически невыгоден. Как следует из рис. 3, характерная длина молекулярно-транспортного расстояния осевых структур С. crinita мало изменяется с ростом структур в длину, но уменьшается с увеличением номера порядка ветвления, что связано с уменьшением размеров их диаметра (Ln = Vn/SbOK = d/4, где: Sbok - боковая поверхность оси, d - её диаметр). Крайние значения Ln структур нулевого и четвертого порядков различаются на порядок величин.
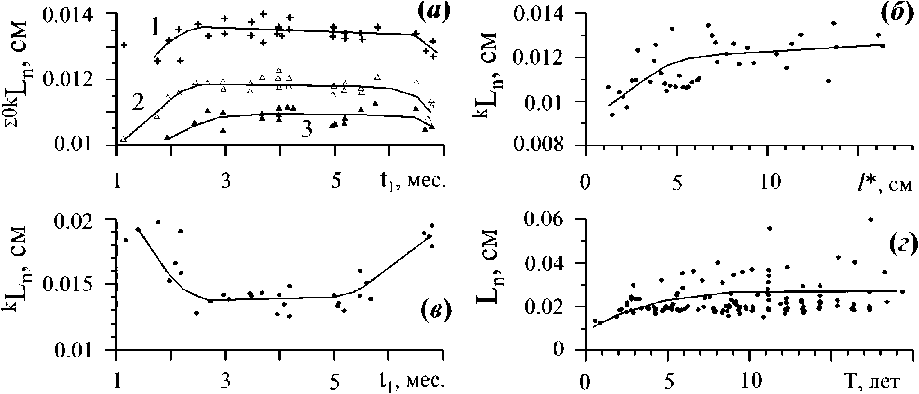
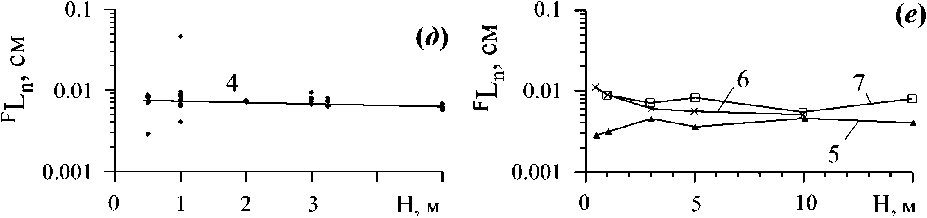
Рис. 4. Значения величин длины молекулярно-транспортного расстояния (Ln) в границах фитосистем телесного уровня организации, каждая из которых представлена множеством физических отдельностей а - изменение средних значений Ln осевых структур С. barbata в онтогенезе ветвей первого порядка (значения рассчитаны по Празукин, 1983); Изменение индивидуальных значений Ln в онтогенетических рядах ветвей второго (С. crinita, б) и первого (С. barbata, в, значения рассчитаны по Празукин, 1983) порядка ветвления и вертикальных побегов С. crinita (г, значения рассчитаны по Хайлов, Парчевский, 1983); д, е - Значения величин Ln макрофитоценозов в условиях разной глубины и уровня антропогенного эвтрофирования на разных участках крымского побережья Черного моря (значения рассчитаны по Ковардаков, Празукин, 2010, 2012). 1, 2, 3 - осевые структуры С. barbata 2, 3 и 4 порядка ветвления соответственно.
Макрофитоценоз из эвтрофных (б. Круглая (4), б. Карантинная (5), севастопольское побережье) и из условно чистых акваторий (б. Планерская (6), б. Ласпи (7)).
В свою очередь, рост только в длину у водорослей с осевой структурой таллома и только в ширину у водорослей с пластинчатой формой слоевища также невыгоден и по причине снижения в этом случае механической прочности таллома.
У водорослей с осевой структурой таллома ветвление является выгодным «приобретением», как с позиций метаболизма, так и с позиций сохранения механической прочности растения в условиях активной подвижности воды. При ветвлении увеличивается площадь ассимилирующей поверхности при относительно небольших изменениях размеров тела, а также уменьшается или сохраняется на относительно постоянном уровне длина молекулярно-транспортного расстояния.
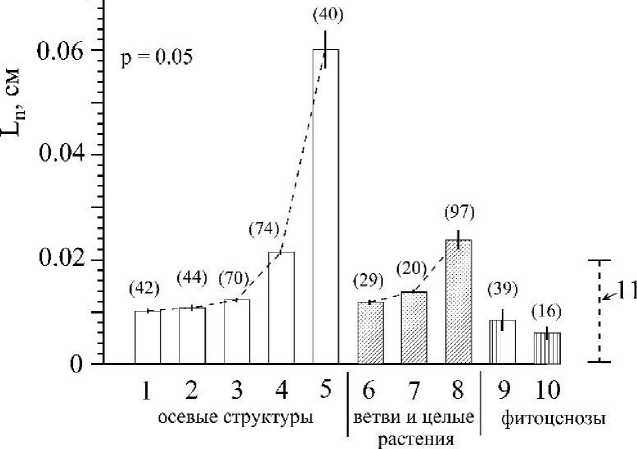
Р и с . 5 . Средние значения длины молекулярно-транспортных расстояний (Ln) в границах телесного пространства у объектов организменного и надоргонизменного уровней организации:
1 - 4 и 5 - соответственно осевые структуры С. crinita четвертого - первого порядка (Ко4 - Koi) и её ствол (Коо); ветви второго (6) и первого (7) порядка и целое слоевище (8) С. crinita; Макрофитоценозы из б. Круглая (9), и из трех бухт: Карантинная, Планерская, Ласпи (10, Ковардаков, Празукин, 2010, 2012); 11- диапазон значений Ln характерный для видов многоклеточных водорослей из рассматриваемых фитоценозов. В круглых скобках указано число исследуемых объектов, р = 0.05.
Как следует из рис. 4 а - г, благодаря ветвлению, созданию множеств из физических отдельностей, цистозиры способны сохранять постоянство длины молекулярно-транспортного расстояния как в онтогенезе отдельной ветви (рис. 4 а - в), так и в онтогенезе вертикального «побега» (рис. 4 г), причем при значительном увеличении их телесного пространства, соответственно, и телесной массы, W (Хайлов Парчевский, 1983; Празукин, 1983, 2015; Ковардаков и др. 1995). У ветвей это постоянство достигается только при наличии всего ансамбля осевых структур. У вертикального побега - за счет динамики верхушечных ветвей (на вертикальном побеге каждого возраста они представлены определенным набором) и за счет структур адвентивного комплекса, роль которых с возрастом «побега» усиливается (Хайлов Парчевский, 1983; Ковардаков и др. 1985; Празукин, 2008).
В фитоценозе регуляция Ln осуществляется так же как и в структуре отдельного растения, через множество отдельностей, за счет изменения вклада в общую массу и поверхность видов водорослей, составляющих ценоз. Как следует из рис. 4 д, е, рассматриваемый показатель удерживается на относительно постоянном уровне в условиях разной глубины.
На рис. 5 представлены значения средних величин Ln по трем группам объектов телесного уровня организации. В ряду осевых структур от К04 к Коо и в ряду: ветви второго и первого порядка -вертикальные побеги происходит нарастающее увеличение средних значений Ln, что может являться ограничивающим фактором в дальнейшем увеличении индивидуальных размеров (W, V) по каждой группе объектов у данного вида водорослей (С. crinita). В свою очередь, характерные значения Ln, рассчитанные для фитоценозов (рис. 5), где два вида цистозир (С. crinita, С. barbata) по массе занимают определяющее положение, оказываются значительно ниже, чем это отмечается для структурных элементов цистозир. Наблюдается последовательное преодоление предельных (метаболически невыгодных) индивидуальных размеров растений, в первом случае -через ветвление структур в границах отдельного растения, во втором -за счет создания множеств особей (популяций) разных видов водорослей в рамках фитоценоза и, в частности, за счет растений эпифитного комплекса с характерными для них малыми значениями Ln.
Фитосистемы надтелесного уровня организации (кроновые и субкроновые системы). Растение своим расположением охватывает часть пространства и создаёт в его границах градиентные условия по веществу и энергии, которые, в свою очередь, накладывают определенные ограничения на форму и размер кроновых систем. Другими словами, форма, размер и пространственная структура кроны растения адаптивно организованы к создаваемым условиям в границах кронового пространства. В таком случае можно говорить о соорганизованности живого и косного.
Растения со сферической и полусферической формами кронового пространства хорошо адаптированы к условиям как направленного, так и рассеянного света (Шульгин и др., 1975). Однако, при таких формах кронового пространства увеличение размеров растений всегда ведет к увеличению длины метаболического пути (Ln+i, рис. 6 в), что невыгодно с позиций внешнего метаболизма.
Как и в случае с объектами телесного уровня организации в границах надтелесного пространства существует структурная и функциональная неоднородность. Например, в случае с водным макрофитом, бриопсисом гипнообразным (Bryopsis hypnoides J.V. Lamouroux, 1809) 63% всей массы растения сосредоточена вблизи верхней границы кронового пространства, т. е. в условиях наилучшего обеспечения фотосинтетически активной радиацией (ФАР). При полусферической форме кронового пространства и высоте 4 см толщина активного слоя поглощения углерода (где 67% углерода поглощается растением) варьировалась в пределах 1.3 - 1.5 см, расчетная величина Ln+i составляла 1.3 см (Празукин, 2007). С увеличением объема кронового пространства увеличивается доля элементов растения, находящихся в условиях пониженного обеспечения ФАР, что в целом ведет к снижению интенсивности фотосинтеза, рассчитанного на единицу объема кронового пространства (цу) и на единицу массы растения (pw). В плане внешнего обмена веществ, растения с малыми размерами кронового пространства находятся в более выигрышном положении, и это может быть одной из многих причин эволюционной трансформации жизненных форм цветковых растений при переходе от деревьев к кустарникам и далее к травам (Тооминг, 1977; Хохряков, 1981).
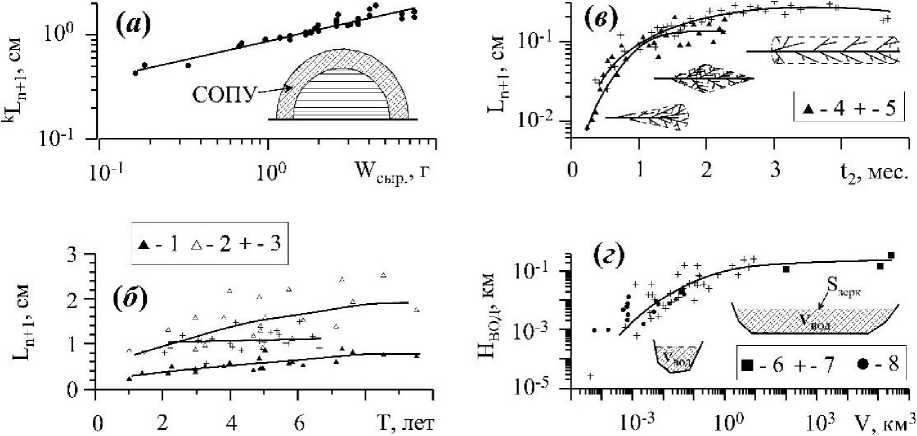
Рис. 6 . Изменение длины молекулярно-транспортного расстояния (Ln+i) и глубины в границах объектов надтелесного уровня организации биогенного и геогенного происхождения а-в размерном ряду (WCMP) слоевищ Bryopsis hypnoides J.V.Lamouroux,1809 (Празукин, 2007). САПУ - слой активного поглощения углерода;
б - в возрастном ряду (Т) вертикальных побегов С. crinita, взятых с глубины 0.5 (3) и 4 (2, 1) м в «раскрытом» (3, 2) и «сомкнутом» (1) состояниях (Празукин, 2005);
в-в суб кроновом пространстве ветвей второго порядка С. crinita в их онтогенезе (12) при возрасте ветвей первого порядка: 2.5 (4) и 3.9 (5) месяца (Празукин, 2005);
г - соотношение водного пространства (Увод) и средней глубины акватории Нвод в водоёмах разного типа и в их геоморфологических зонах (Хайлов и др., 2009). В глубинных зонах шельфа Баренцева моря (6), в губах и бухтах Кольского полуострова и в их глубинных зонах (7), в бухтах и их глубинных зонах Чёрного моря (8).
Растения с цилиндрической и близкой к ней формой кронового пространства, увеличиваясь преимущественно в длину, сохраняют значения Ln+i на относительно неизменном уровне (рис. 6 б). Это хорошо видно в случае со слоевищами С. crinita, обитающими в условиях активной подвижности воды. Растения, находясь в постоянно меняющемся потоке воды, приобретают сигарообразную форму кронового пространства, а их длина молекулярнотранспортного расстояния на протяжении всего онтогенеза практически не меняется и удерживается в узком диапазоне значений. Тогда как на глубине 4 м, в условиях относительно слабой подвижности воды, вертикальные побеги С. crinita представлены большим разнообразием форм кроновых пространств, преимущественно это сочетание фигур: цилиндра, конуса и полусферы. Соответственно, диапазон варьирования значений Ln+i здесь гораздо шире, чем это наблюдается при активной подвижности воды, а её средние значения монотонно увеличиваются в онтогенезе растений. Эти же растения, помещенные в условиях эксперимента в направленный поток воды, принимали сигарообразную форму кронового пространства, и в этом случае наблюдается двукратное уменьшение значений длины метаболического пути, и одновременно сужается и диапазон её варьирования. Представленные примеры демонстрируют возможные диапазоны изменений длины молекулярно-транспортного расстояния (Ln+i) в границах кронового пространства при эндогенной и экзогенной регуляции. В то же время, форма надтелесного пространства ветвей второго порядка С. crinita мало зависит от условий подвижности воды, но закономерно меняется в онтогенезе (рис. 2 л). У ветвей в зрелом возрасте формируется цилиндрическая форма субкронового пространства, при которой длина молекулярно-транспортного расстояния мало зависит от размеров надтелесного пространства (рис. 6 в) при условии приоритетного увеличения ветви в длину.
Соотношение объёма водного пространства и глубины акватории в водоёмах разного типа. Растения и животные своей массой заполняют определенную часть географического пространства и, в частности, водоемы. В границах отдельного водоёма, с участием обитающих в нем растений и животных, формируется биокосная система, структура и функция которой во многом определена геоморфологией водоёма. Одна из важных геоморфологических характеристик водоема, а значит и биокосной системы в его границах - это его средняя глубина (НВОд), определяемая через соотношение: Увод/8зерк (рИС. 2 Г).
На рис. 6 г показано соотношение V и Нвод в глубинных зонах Баренцева моря, в его бухтах и губах на Кольском полуострове, а также в бухтах и разноглубинных зонах Черного моря (Хайлов и др., 2009). Наблюдается общая зависимость НВОд по V, на фоне которой каждая группа водоёмов имеет свое характерное положение в общей метрике, т.е. обнаруживает специфические особенности своей геоморфологии. Можно сказать, что геоморфология водного ложа является информационной матрицей, которая передается в морфологию водоема. Геоморфологическая конфигурация водоёма, рельеф его дна, химический состав воды и другие характеристики водоема, в свою очередь, передаются в его биоту и интегрируются в таких важных биологических параметрах водоёмов, как общая масса живого вещества (W) и её концентрация (Cw) в воде (Хайлов, 2003; Хайлов и др., 2009).
Заключение. Подводя итог статьи отметим, что матричная роль (импринтинг) Земли последовательно проявляется на всех уровнях жизни в геометрии обитаемых пространств. В пространстве отдельного водоема с его обитателями, в пространстве растительного полога и в кроновых пространствах отдельных растений, и в пространстве тела растения. Поступление вещества и энергии в обитаемое пространство и их распределение в нём, и, соответственно, распределение жизни и её деятельность во многом предопределены геометрий пространства, соотношением «питающего» объёма и «питаемой» поверхностью (L = V/S).
Длина молекулярно-транспортного расстояния стабилизируется в онтогенетических и размерных рядах объектов телесного и надтелесного уровней организации за счет формирования оптимальной для этого геометрической формы обитаемого пространства и через создание множеств из физических отдельностей со свойственной им структурой и функцией.
«Длина молекулярно-транспортного расстояния» не является реальным выражением длины молекулярного пути в объёме пространства, но использование этой характеристики как инструмента в описании пространственных и метаболических отношений («местофункционирование»), существующих в иерархически организованной фитосистеме, полезно. Предлагаемый подход позволяет однотипно рассматривать объекты разного уровня организации и происхождения и сравнивать их в общем метрическом поле. В итоге мы можем говорить о структурном подобии рассматриваемых объектов и, возможно, о функциональном. В таком случае, «местофункционирование» становится самостоятельным объектом рассмотрения.
Список литературы Молекулярно-транспортные расстояния в обитаемых пространствах в иерархически организованной фитосистеме
- Зинова А.Д. 1967. Определитель зеленых, бурых и красных водорослей южных морей СССР. Л.: Наука. 400 с.
- Калугина-Гутник А.А. 1975. Фитобентос Черного моря. Киев: Наук. думка. 248 с
- Камнев А.Н. 1989. Структура и функция бурых водорослей. -М.: Изд-во МГУ. 200 с.
- Камнев А.Н., Бурдин К.С., Камнева М.А., Родова Н.А. 1988. Анатомическое строение разновозрастных слоевищ бурой водоросли Sargassum pallidum (TURN) C. Ag.//Известия АН СССР. № 2. С. 301-305.
- Ковардаков С.А., Празукин А.В. 2010. Структурно-функциональные характеристики фитоценозов морских прибрежных акваторий Крыма.//Системы контроля окружающей среды: сборник научных трудов/Морской Гидрофизический институт НАН Украины. Севастополь. Вып. 14.С. 230-235.
- Ковардаков С.А., Празукин А.В. 2012. Cтруктурно-функциональные характеристики донного фитоценоза бухты Круглой (Севастополь)//Экосистемы, их оптимизация и охрана. Вып. 7. С. 138-148.
- Ковардаков С.А., Празукин А.В., Фирсов Ф.К., Попов А.Е. 1985 Комплексная адаптация цистозиры к градиентным условиям. Киев: Наук. думка. 216 с.
- Празукин А.В. 1983.Феноменологическое описание роста ветвей Cystoseira barbata как основа периодизации их онтогенеза.//Экология моря. Вып. 13. С. 49-58
- Празукин А.В. 2000. Структура кронового пространства слоевища черноморской бурой водоросли цистозиры (Сystoseira crinita (Desf.) Bory) (Phaeophyta)//Альгология. Т. 10. № 2. C. 119-130.
- Празукин А.В. 2005. Иерархическое членение обитаемого пространства и его заполнение биоорганическим веществом на примере черноморской водоросли цистозиры (Cystoseira crinita (Desf.) Bory)//Морской экологический журнал. Т. 4. № 3. C. 15-36.
- Празукин А.В. 2007. Различные приемы описания фитосистем разного уровня организации на примере зеленой водоросли бриопсис (Bryopsis Lamouroux) Черного моря//Морской экологический журнал. Т. 6. № 1. C. 66-87.
- Празукин А.В. 2008. Морфофункциональные образы растения (на примере многоклеточных водорослей)//Вестн. ТвГУ. Сер. Биология и экология. Вып. 9. С. 206-218.
- Празукин А.В. 2015. Экологическая фитосистемология. М.: Перо. 375 с.
- Празукин А.В. 2016. Морфометрический подход в оценке величины площади поверхности таллома многоклеточной водоросли Callithamnion corymbosum (J. E. Smith) Lyngb//Вестн. ТвГу. Сер.: Биология и экология. № 4. С.195-205
- Празукин А. В. 2017. Иерархическое членение кронового пространства многоклеточной водоросли Callithamnion corymbosum (J. E. Smith) Lyngbye, 1819 и его заполнение биоорганическим веществом//Вестник Тверского государственного университета. Сер.: Биология и экология. № 1. С. 196-206.
- Празукин А.В., Хайлов К.М., Ковардаков С.А. 2003.Сравнение структурно-функциональных соотношений в морских фитосистемах разного уровня организации на основе их унифицированного описания)//Морской Экологический журнал. Т. 2, № 3. С. 51-62.
- Тооминг Х.Г. 1977. Солнечная радиация и формирование урожая. Л.: Гидрометеоиздат. 200 с.
- Хайлов К.М. 2001. Что такое жизнь на Земле? Одесса: Друк. 238 с.
- Хайлов К.М. 2003. Косное вещество биосферы и распределение живой массы растений, животных и людей.//Известия АН. Серия географическая. № 5. С. 7-15.
- Хайлов К. М., Ковардаков С. А., Празукин А. В. 2004. Биологические поверхности многоуровневых фитосистем и расчет их численных характеристик.//Мор. экологический журнал. Т. 3, № 3. С. 61-77.
- Хайлов К.М. Ковардаков С.А., Шошина Е.В. 2002. Связь объемной концентрации донных водорослей с геометрическими параметрами обитаемых пространств//Океанология. Т. 42, № 3. C. 402-407.
- Хайлов К.М., Парчевский В.П. 1983. Иерархическая регуляция структуры и функции морских растений. Киев: Наукова думка. 253 с.
- Хайлов К.М., Празукин А.В, Смолев Д.М., Юрченко Ю.Ю. 2009. Школа биогеоэкологии. Севастополь: ЭКОСИ-Гидрофизика. 325 с.
- Хайлов К.М., Фирсов Ю.К. 1985. Расчет величины ассимиляционной поверхности Sargassum на разных уровнях организации//Ботанический журнал Т. 70. № 2. С. 215-221.
- Хайлов К. М., Юрченко Ю. Ю., Шошина Е. Е. 2001. О связи геометрических, биологических и трофодинамических характеристик водных биокосных фитосистем.//Океанология. Т. 41, № 3. С. 400-407.
- Хохряков А.П. 1981. Эволюция биоморф растений. М.: Наука. 168 с.
- Шульгин И.А., Климов С.В., Ничипорович А.А. 1975. Об адаптивности архитектоники растений к солнечной радиации.//Физиология растений. Т. 22, вып. 1. С. 40-48.
