Молекулярные и клеточные механизмы действия смешанного гамма-нейтронного излучения различной мощности дозы

Автор: Рябченко Н.И., Антощина М.М., Насонова В.А., Фесенко Э.В., Рябченко В.И., Иванник Б.П., Ульяненко С.Е., Соколов В.А.
Рубрика: Научные статьи
Статья в выпуске: 1-2 т.15, 2006 года.
Бесплатный доступ
Облучение проводили на импульсном реакторе Барс-6 (ГНЦ РФ - ФЭИ, Обнинск) в виде одиночного импульса длительностью порядка 100 мксек и в виде непрерывного смешанного гамма-нейтронного излучения длительностью 60 минут при мощностях дозы, отличающихся в 3,6-107 раз. Полученные данные свидетельствуют о дифференциальной чувствительности изученных показателей к повреждающему действию смешанных гамма-нейтронных излучений различной мощностью дозы. Проведенные исследования показали, что при облучении мышей в дозах до 2 Гр импульсное излучение в 1,3-1,7 раз более эффективно увеличивает число аберраций в клетках костного мозга по сравнению с непрерывным излучением. В тоже время не было найдено статистически достоверных различий между этими режимами облучения при анализе хромосомных аберраций, индуцированных действием гамма-нейтронного излучения при облучении лимфоцитов человека на стадии G0. Повышенная эффективность цитогенетического действия импульсного гамма-нейтронного излучения на костный мозг мышей может быть связана с естественной гетерогенностью клеточной популяции кариоци-тов, находящихся на различных стадиях клеточного цикла, и увеличением выхода аберраций хромосом в этих клетках за счет ингибирования репарационных процессов при импульсном режиме облучения. В тоже время полученные нами данные свидетельствуют о том, что непрерывный режим облучения более эффективен в повышении уровня полиплоидизации клеток и нарушении мембранных структур. Полученные экспериментальные результаты и анализ данных литературы свидетельствует о перспективности использования импульсных гамма-нейтронных излучений для терапии злокачественных новообразований.
Короткий адрес: https://sciup.org/170169896
IDR: 170169896
Molecular and cellular mechanisms of action of mixed gamma-neutron irradiation at various dose rates
Exposures to mixed gamma-neutron radiation were performed using a BARS-6 pulsed reactor (PEI, RF, Obninsk) either in a regimen of one 100-μsec pulse or in a regimen of continuous 60-min irradiation at dose rates distinguished in 3.6-107 times. The average energy of neutrons was 1.4 MeV. The obtained data testify to differential sensitivity of the investigated parameters to damaging action of mixed gamma-neutron radiation with various of dose rates. It was shown, that irradiation of mice with pulse radiation was 1.3-1.8 times more effective in the induction of chromosome aberrations in bone marrow cells in comparison with continuous regimen of irradiation. At the same time, other biological tests (yield of chromosome aberrations in human lymphocytes, decreasing of the number of cells in thymus) demonstrated that pulsed and continuous regimens have almost equal biological effectiveness. Analysis of chromosome aberration spectra showed that over 90 % of aberrations induced in cultured human blood lymphocytes were chromosome-type aberrations, whereas the proportion of chromosome-type aberrations induced in mice bone marrow cells was 46-58 %. These results suggest that the population of bone marrow cells was represented by cells which were at various cell cycle stages. The increased effectiveness of pulsed gamma-neutron radiation in induction of chromosome aberrations in mice bone marrow may be due to an increase in the proportion of dividing cells in the bone marrow cell population. It was shown that the continuous mode of an irradiating is more effective in rise of polyploidy cells and injury of membranous frames. The obtained experimental results evidence about availability of usage of impulse gamma-neutron radiation for therapy of malignant neoplasm.
Текст научной статьи Молекулярные и клеточные механизмы действия смешанного гамма-нейтронного излучения различной мощности дозы
В последние годы для медико-биологических исследований начали использоваться источники импульсных нейтронных излучений с частотой следования импульсов от 1 до 1000 Гц и мощностью дозы в импульсе более 105 Гр/с, которые могут быть перспективны для лучевой терапии злокачественных новообразований [1, 2]. Изменение временного фактора воздействия поглощенной дозы может оказывать различное влияние на реакцию клеток и тканей, как в сторону усиления, так и в сторону ослабления лучевого воздействия [3]. В связи с этим, представляет интерес импульсный ядерный реактор БАРС-6, созданный в ГНЦ РФ – ФЭИ, Обнинск, который позволяет проводить облучение биологических объектов смешанным гамма-нейтронным излучением в виде одиночного импульса длительностью порядка ~100 мкс (мощность дозы до 106 Гр/с) и в виде непрерывного гамма-нейтронного излучения длительностью 3600 с (мощность дозы до 10-2 Гр/с) [2, 4]. Данные проведенных исследований биологической эффективности смешанных гамма-нейтронных излучений различной мощности дозы противоречивы. Результаты работ, выполненных на лимфоцитах человека [5] и клетках млекопитающих V-79 [6] и аберраций хромосом костного мозга облученных мышей [7], указывали на большую эффективность повреждающего действия импульсного излучения реактора БАРС-6. Однако в других работах при исследовании выживаемости микроорганизмов [8], аберраций хромосом лимфоцитов человека [7, 9] и выживаемости животных [10] не удалось выявить разницы в биологическом действии импульсного и непрерывного нейтронного излучения.
В настоящей работе обобщены результаты исследований по оценке на молекулярном и клеточном уровнях биологического действия смешанных гамма-нейтронных излучений различной мощности дозы. В качестве объектов исследований были использованы облученные на реакторе БАРС-6 в непрерывном режиме и в режиме одиночного импульса суспензии липосом, растворы белков и ферросульфата, лимфоциты периферической крови человека (появление структурных и численных повреждений хромосом), а также структурные аберрации в костном мозге облученных мышей.
Материалы и методы
Конструкция реактора БАРС-6 (ГНЦ РФ – ФЭИ, Обнинск) позволяла проводить облучение в импульсном и непрерывном режимах при сохранении в этих условиях одинакового (с точностью до 2 %) числа делений в АЗ (активных зонах) реактора [2, 4]. Благодаря этому биологические объекты, размещенные на одних и тех же фиксированных расстояниях от реактора, получали за 100 мкс и 60 мин одну и ту же дозу смешанного гамма-нейтронного излучения при мощностях дозы, отличающихся друг от друга в 3,6 - 10 7 раза. Показано, что по мере увеличения расстояния от реактора меняется не только доза смешанного гамма-нейтронного излучения, но и средняя энергия нейтронов (от 1,4 до 0,5 МэВ), а также увеличивается до 50 % вклад гамма-компоненты в общую дозу гамма-нейтронного излучения. Однако при облучении объектов, расположенных на одних и тех же расстояниях от реактора, энергетический спектр нейтронов и соотношение доз нейтронной и гамма-компонент сохраняли свои значения при облучении в режиме одиночного импульса и непрерывном режиме [4]. Измерение флюенса быстрых нейтронов проводили с помощью активационного детектора из никеля (марка НП-2М), а для определения величины флюенса нейтронов всех энергий вносили поправку на спектр нейтронов на различных расстояниях от реактора. Измерение дозы у -излучения проводили с помощью тер-молюминисцентного дозиметра ИКС-А с использованием термолюминисцентного алюмофос-фатного стекла ИС-7 [4].
Мультиламеллярные липосомы получали из фосфолипидов желтков куриных яиц. Эффективность радиационного окисления фосфолипидов липосом оценивали по выходу малондиальдегида (МДА), который является одним из конечных продуктов перекисного окисления липидов. Концентрацию МДА определяли с помощью тиобарбитуровой кислоты (ТБК) по модифицированному методу [11]. Для проведения определения к 0,5 мл суспензии липосом прибавляли 2,5 мл раствора, содержащего 12 % трихлоруксусной кислоты (ТХУ), 0,3 % ТБК, 0,2 М HCl, 0,004 % ЭДТА и 0,0002 % ионола. Полученную смесь прогревали в течение 20 минут при 100 ° С, охлаждали и центрифугировали. Супернатант фотометрировали при 532 нм и рассчитывали концентрацию МДА, используя коэффициент экстинкции, равный 156000 М-1 см-1.
Для цитогенетического анализа аберраций хромосом, индуцированных в лимфоцитах на стадии G 0 , облучали гепаринизированную (200 ед/мл) кровь. Облучение крови гамма-квантами 60Со проводили на установке «Луч» («Изотоп», Россия) при мощности дозы 0,5 Гр/мин. Облучение крови на реакторе БАРС-6 проводили в пластиковых пробирках, которые размещали на различных фиксированных расстояниях от реактора. В опытах использовали стандартную методику культивирования лимфоцитов человека, согласно которой через 3-4 часа после облучения 1 мл гепаринизированной крови добавляли к 8 мл культуральной среды, состоящей из 80 % среды RPMI c глютамином, 20 % эмбриональной сыворотки телят, антибиотиков пенициллина и стрептомицина. Затем в среду для культивирования для стимуляции деления лимфоцитов вносили ФГА (10-20 мкг/мл) и для дифференциальной окраски хроматид вносили бромдезоксиуридин в конечной концентрации 20 мкг/мл. Идентификацию метафаз первого, второго, третьего и четвертого митозов в облученных и необлученных лимфоцитах проводили по модифицированной методике дифференциального окрашивания сестринских хроматид с помощью бромдезоксиуридина [7]. За два часа до фиксации клеток для накопления метафаз в среду для культивирования вводили колхицин в конечной концентрации 0,5 мкг/мл. Для получения цитологических препаратов клетки гипотонизировали, фиксировали смесью ледяной уксусной кислоты и метилового спирта в соотношении 1:3 и окрашивали азур-эозином. На цитологических препаратах в метафазах первого митоза, содержащих 44-46 хромосом, анализировали структурные аберрации хромосом. В метафазах 1-го, 2-го, 3-го и 4-го митозов определяли также число полиплоидных клеток (ППК), к которым относили тетраплоидные клетки, а также гипотетраплоидные клетки, содержащие более 86 хромосом.
В опытах использовали мышей линии С57Bl и F1(CBAxC57Bl). Животных облучали в контейнерах из органического стекла толщиной 5 мм, расположенных на различных расстояниях от реактора. При проведении облучения на реакторе БАРС-6 в двух изучаемых режимах контейнеры с животными располагали на одних и тех расстояниях от реактора. Через 24 часа после облучения мышей забивали, извлекали костный мозг и тимус, которые использовали для определения аберраций хромосом и числа тимомоцитов. За 2 часа до забоя для накопления метафаз мышам вводили внутрибрюшинно по 0,2 мл 0,025 %-ного раствора колхицина. Клетки костного мозга гипотонизировали в 0,075 М KCl в течение 20 минут при температуре 37 ° С. По окончании гипотонизации клетки осаждали центрифугированием, фиксировали в смеси, состоящей из метилового спирта и ледяной уксусной кислоты в соотношении 3:1, наносили на предметные стекла, высушивали над пламенем и окрашивали азур-эозином.
Результаты и обсуждение
Особенность использования реактора БАРС-6 в режиме одиночного импульса и непрерывном режиме состоит в том, что в обоих режимах потоки нейтронов создавали одну и ту же поглощенную дозу смешанного гамма-нейтронного излучения, величина которой определялась расстоянием от реактора [2, 4]. Обработка методом двухфакторного дисперсионного анализа экспериментально полученных результатов дозиметрических измерений подтвердила этот факт и показала отсутствие (p>0,05) статистически достоверной разницы в поглощенных дозах, создаваемыми двумя режимами облучения на одних и тех расстояниях от реактора. Результаты обработки экспериментальных данных показали, что вклад гамма-компоненты в общую дозу смешанного гамма-нейтронного излучения различен на различных расстояниях от реактора (р<0,05), однако для режима одиночного импульса и непрерывного режима облучения этот вклад одинаков (p>0,05) для объектов, расположенных одних и тех же расстояниях от реактора. В табл. 1 приведены для различных расстояний от реактора БАРС-6 усредненные для этих режимов данные о поглощенных дозах смешанного гамма-нейтронного излучения и вкладе в общую дозу гамма-компоненты. Следует отметить, что дозиметрия гамма-нейтронного импульсного излучения представляет серьезную научную задачу, поскольку на разных расстояниях от реактора не только изменяется соотношение гамма- и нейтронного излучения, но, что более существенно, изменяется и энергетический спектр нейтронов. В связи с этим данные табл. 1 следует рассматривать в качестве полученных с точностью 15-25 % оценочных данных о поглощенных дозах, и которые в процессе проведения дальнейших исследований будут уточняться.
Таблица 1 Дозиметрические характеристики смешанного гамма-нейтронного излучения на различных расстояниях от реактора
|
Расстояние от реактора , м |
Доза смешанного гамма - нейтронного излучения , Гр |
Доля γ - компоненты в общей дозе излучения , % |
Мощность дозы при различных режимах облучения , Гр / с |
||
|
одиночный импульс II |
непрерывный |
||||
|
0 |
808,0 |
8,0 |
8,1 ⋅ |
106 |
0,22 |
|
100 |
671 |
8,7 |
6,7 ⋅ |
106 |
0,19 |
|
200 |
371 |
9,8 |
3,7 ⋅ |
106 |
0,10 |
|
300 |
191 |
12,0 |
1,9 ⋅ |
106 |
0,053 |
|
400 |
117 |
12,7 |
1,2 ⋅ |
106 |
0,032 |
|
800 |
40 |
20,1 |
0,4 ⋅ |
106 |
0,011 |
|
6500 |
2,8 |
37,9 |
2,6 ⋅ |
106 |
0,047 |
|
8500 |
2,1 |
46,2 |
2,0 ⋅ |
106 |
0,036 |
|
10200 |
1,6 |
50,8 |
1,5 ⋅ |
106 |
0,026 |
|
12000 |
1,1 |
44,3 |
1,0 ⋅ |
106 |
0,018 |
|
15000 |
0,62 |
35,7 |
0,6 ⋅ |
106 |
0,010 |
Примечание : расстояние 0 мм означает , что образцы были размещены непосредственно на наруж ной поверхности реактора .
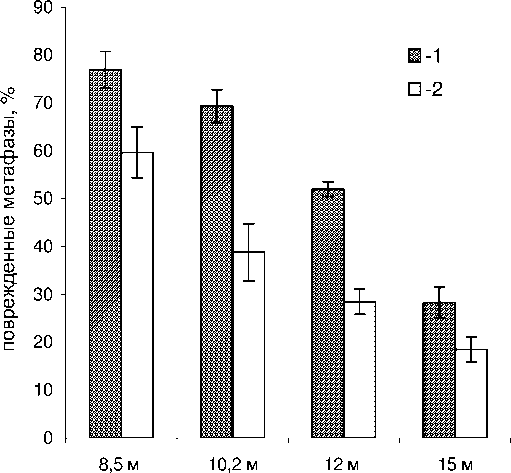
Учитывая особенности облучения на реакторе БАРС-6, нами сравнивалось действие излучений различной мощности дозы на объекты, которые были расположены на одинаковых для обоих режимов расстояниях от реактора. Было проведено облучение мышей смешанным гамма-нейтронным излучением реактора БАРС-6 в режиме одиночного импульса длительностью порядка 100 мкс и в режиме непрерывного облучения длительностью 60 минут. В каждую группу мышей, облученных в этих режимах на различных расстояниях от реактора, входило по 3-4 животных, у каждого из которых анализировали по 100 метафаз. На рис. 1 и 2 представлены данные, полученные при цитогенетическом анализе хромосомных аберраций в костном мозге мышей линии С57Bl и мышей линии F1(CBAxC57Bl), облученных на реакторе БАРС-6. У необ-лученных животных в костном мозге мышей линии С57Bl6 содержание поврежденных метафаз составляло 2,3±0,9 %, а у мышей линии F1(CBAxC57Bl) процент поврежденных метафаз составлял 0,6±0,4 %. Полученные результаты свидетельствуют о том, что через 24 часа после облучения мышей в костном мозге увеличивается содержание метафаз с аберрациями хромосом. Выход аберраций увеличивается по мере уменьшения расстояния от реактора, что является следствием увеличения дозы облучения по мере приближения мышей к реактору. Методом двухфакторного дисперсионного анализа была проведена статистическая обработка данных о появлении поврежденных метафаз в костном мозге мышей при облучении в режиме одиночного импульса и непрерывном режиме. Результаты проведенной статистической обработки данных показали у анализируемых линий мышей наличие достоверной (p<0,001) разницы между двумя режимами облучении в отношении индукции поврежденных метафаз. Аналогичные данные о более эффективном цитогенетическом воздействии облучения в режиме одиночного импульса были получены при анализе общего числа аберраций в метафазах костного мозга двух линий мышей. В отличие от результатов определения аберраций хромосом при анализе клеточности тимуса не было обнаружено статистически достоверной разницы в эффективности лимфолитического действия режима облучения одиночным импульсом и непрерывного режима облучения.
расстояние от реактора, м
Рис. 1. Поврежденные метафазы, %, в костном мозге мышей линии С57Bl, облученных на реакторе БАРС-6 при различных режимах облучения.
-
1 – облучение одиночным импульсом ; 2 – непрерывный режим облучения .
Абсцисса – расстояние от реактора , м . Ордината – % поврежденных метафаз .
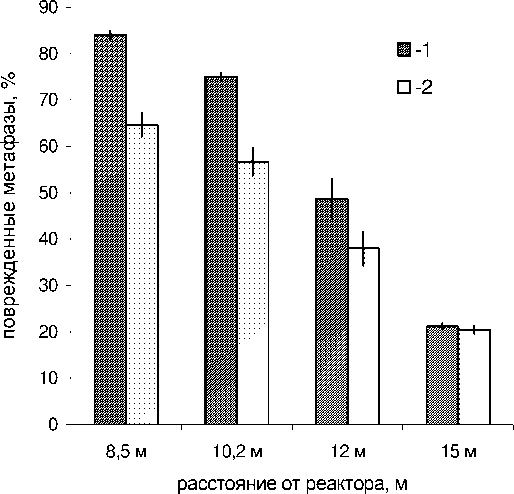
Рис. 2. Поврежденные метафазы, %, в костном мозге мышей линии F1(С57BlxCBA), облученных на реакторе БАРС-6 при различных режимах облучения.
1 – облучение одиночным импульсом ; 2 – непрерывный режим облучения . Абсцисса – расстояние от реактора , м . Ордината – % поврежденных метафаз .
Было изучено влияние различных режимов гамма-нейтронного излучения реактора БАРС-6 на выход хромосомных аберраций в метафазах первого митоза облученных лимфоцитов периферической крови человека. Проведенные ранее исследования [12] показали, что выход аберраций хромосом в метафазах первого митоза одинаков при различных (до 120 часов) сроках культивирования облученных лимфоцитов. Поэтому нами были суммированы результаты определения аберраций хромосом в метафазах первого митоза, полученных после культивирования лимфоцитов с ФГА и бромдезоксиуридином в течение 48 и 72 часов. В контроле в метафазах первого митоза процент поврежденных клеток составлял в среднем 1,1 ± 0,5 %. Как следует из данных, представленных на рис. 3, в метафазах первого митоза при облучении смешанным гамма-нейтронным излучением реактора БАРС-6 выход аберраций хромосом увеличивается по мере приближения облучаемых образцов к активной зоне реактора. Методом двухфакторного дисперсионного анализа была проведена статистическая обработка данных о появлении поврежденных метафаз при облучении в режиме одиночного импульса и непрерывном режиме облучения лимфоцитов. Результаты проведенного статистического анализа показали отсутствие достоверной (p>0,05) разницы в появлении поврежденных метафаз под влиянием этих режимов облучения. Аналогичные результаты об отсутствии достоверной разницы между эффективностью действия гамма-нейтронного излучения различной мощности дозы на лимфоциты человека были получены и при двухфакторном дисперсионном анализе результатов определения числа индуцированных структурных аберраций, приходящихся на одну метафазу первого митоза.
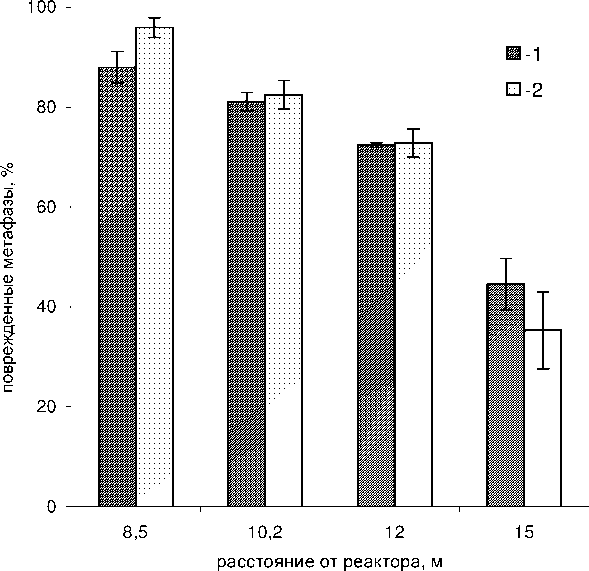
Рис. 3. Поврежденные метафазы первого митоза, %, в лимфоцитах периферической крови человека, облученных на реакторе БАРС-6 при различных режимах облучения. 1 – облучение одиночным импульсом; 2 – непрерывный режим облучения.
Абсцисса – расстояние от реактора, м. Ордината – % поврежденных метафаз.
Для анализа возможных механизмов различной чувствительности кариоцитов костного мозга мышей и лимфоцитов человека к воздействию гамма-нейтронного излучения в режиме одиночного импульса и непрерывном режиме был исследован качественный состав индуцированных структурных аберраций хромосом. В табл. 2 приведены результаты определения соотношения аберраций хромосомного и хроматидного типов в лимфоцитах человека и костном мозге мышей при различных условиях облучения гамма-квантами 60Со и гамма-нейтронным излучением реактора БАРС-6. При определении этого соотношения были суммированы результаты анализов аберраций хромосом при дозах излучения, индуцирующих более 10 % поврежденных метафаз. Лимфоциты периферической крови представлены естественно синхронизированной популяцией клеток, находящихся на стадии G0. Анализ качественного состава структурных аберраций хромосом (табл. 2) свидетельствует о том, что при воздействии гамма-квантов на лимфоциты в стадии G0 (дозы 0,5-3 Гр) в метафазах индуцируются структурные аберрации, более 90 % которых представлены аберрациями хромосомного типа. В то же время при облучении лимфоцитов на стадии G2 более 95 % индуцируемых аберраций составляют аберрации хроматидного типа. При облучении лимфоцитов на стадии G0 смешанным гамма-нейтронным излучением с различной мощностью дозы более 90 % индуцируемых аберраций представлены аберрациями хромосомного типа (табл. 2). Эти результаты находятся в соответствии с извест- ными в литературе данными [13] о смене типов аберраций хромосом при облучении клеток на различных стадиях клеточного цикла. Согласно этим данным, лучевое воздействие на пресин-тетических стадиях G0 и G1 индуцирует появление аберраций хромосомного типа. При облучении на постсинтетической стадии G2 индуцируются аберрации хроматидного типа, а на стадии S в метафазах появляются аберраций как хромосомного, так и хроматидного типов.
Таблица 2 Соотношение аберраций хромосомного (ХС) и хроматидного (ХТ) типов в лимфоцитах человека и костном мозге (КМ) мышей при различных условиях гамма ( у ) облучения и гамма-нейтронного ( у , n) облучения на реакторе БАРС-6 в режиме одиночного импульса и непрерывном режиме
|
Объект |
Условия облучения |
Число метафаз |
Общее число ХС и ХТ аберраций |
Соотношение, %, ХС и ХТ аберраций |
|
|
Лимфоциты, стадия G0 |
γ -кванты 60Со, 0,5 Гр/мин |
3167 |
1401 |
ХС ХТ: |
: 94,3 ± 3,6 5,7 ± 3,6 |
|
Лимфоциты, стадия G2 |
γ -кванты 60Со, 0,5 Гр/мин |
1350 |
1287 |
ХС ХТ: |
: 1,7 ± 1,3 98,3 ± 1,3 |
|
Лимфоциты, стадия G0 |
( γ ,n)-излучение, импульс 100 мкс |
1246 |
1889 |
ХС ХТ: |
: 97,1 ± 3,2 2,9 ± 3,2 |
|
Лимфоциты, стадия G0 |
( γ , n)-излучение, непрерывный |
1067 |
1477 |
ХС ХТ: |
: 96,1 ± 3,6 3,9 ± 3,6 |
|
КМ, мыши С57Bl |
γ -кванты 60Со, 0,5 Гр/мин |
700 |
227 |
ХС ХТ: |
: 45,4 ± 5,4 54,6 ± 5,4 |
|
КМ, мыши F1(CBAxC57Bl) |
γ -кванты 60Со, 0,5 Гр/мин |
1400 |
632 |
ХС ХТ: |
: 38,1 ± 2,9 62,9 ± 2,9 |
|
КМ, мыши С57Bl |
( γ , n)-излучение, импульс 100 мкс |
1200 |
2530 |
ХС ХТ: |
: 58,7 ± 1,9 41,3 ± 1,9 |
|
КМ, мыши С57Bl |
( γ , n)-излучение, непрерывный |
1563 |
1505 |
ХС ХТ: |
: 57,1 ± 1,5 42,9 ± 1,5 |
|
КМ, мыши F1(CBAxC57Bl) |
( γ , n)-излучение, импульс 100 мкс |
1285 |
4208 |
ХС ХТ: |
: 46,2 ± 1,3 53,8 ± 1,3 |
|
КМ, мыши F1(CBAxC57B) |
( γ , n)-излучение, непрерывный |
1500 |
3026 |
ХС ХТ: |
: 52,8 ± 1,8 47,2 ± 1,8 |
Как следует из данных, представленных в табл. 2, у мышей, облученных на реакторе БАРС-6, в метафазах костного мозга доля индуцируемых аберраций хромосомного типа составляет 46-58 %. Возможной причиной повышения выхода аберраций хроматидного типа состоит в том, что популяция кариоцитов гетерогенна и представлена клетками, находящимися в момент облучения на пресинтетической, синтетической и постсинтетической стадиях клеточного цикла. При облучении такой гетерогенной популяции, с учетом возможной радиационной задержки продвижения по циклу, индуцируются аберрации как хромосомного, так и хроматидного типов. Следует также отметить, что согласно данным литературы [13, 14] повышенный уровень аберраций хроматидного типа может также указывать на появление в пресинтетической стадии клеточного цикла длительно живущих потенциальных повреждений, которые реализуются в постсинтетической стадии в хроматидные аберрации. Предполагается [13, 14], что появление таких потенциальных повреждений может быть следствием ингибирования репарации разрывов ДНК.
Наряду со структурными аберрациями хромосом, появляющимися в результате воздействия кластогенных факторов (ионизирующее излучение, блеомицин, митомицин), важнейшим показателем нарушения хромосомного аппарата клеток является изменения числа хромосом, вызываемое действием анеогенных факторов (винкрестин, винбластин, таксол и другие), нарушающих структуру митотического веретена. Предполагается, что анеуплоидия и полиплоидия играет существенную роль в индукции канцерогенеза и хромосомной нестабильности [15, 16]. Проведенные нами исследования показали, что под влиянием смешанного гамма-нейтронного излучения различной мощности в метафазах лимфоцитов человека увеличивается число полиплоидных клеток (ППК).
При анализе 979 метафаз первого, второго, третьего и четвертого митозов необлученных лимфоцитов не было найдено ни одной ППК, в то время как при анализе 5297 метафаз первого-четвертого митозов облученных лимфоцитов было найдено 335 ППК. Из 335 ППК в метафазах первого митоза было найдено 8 ППК (2,4 %), в метафазах второго митоза было найдено 233 ППК (69,6 %), в метафазах третьего митоза было найдено 91 ППК (27,2 %) и 3 ППК (0,8 %) было найдено в метафазах четвертого митоза. Полученные данные свидетельствуют о том, что более 96 % ППК, индуцированных гамма-нейтронным излучением, появляются в метафазах второго и третьего митозов. В связи с этим представляло интерес выяснить связь между числом аберраций хромосом, индуцированных облучением в метафазах первого митоза, и долей (%) ППК в метафазах второго и третьего митозов. Для этого образцы крови располагали на различных расстояниях (15 м, 12 м и 10,2 м) от активной зоны реактора, облучали в условиях одиночного импульса и непрерывном режиме. Данные по анализу процента аберрантных метафаз первого митоза при различных дозах гамма-нейтронного излучения различной мощности дозы приведены на рис. 3. На рис. 4 приведены данные о проценте ППК в метафазах второго и третьего митозов при различных уровнях повреждения хромосом в метафазах первого митоза. Полученные данные свидетельствуют о том, что по сравнению с импульсным режимом при одном и том же числе структурных аберраций хромосом непрерывный режим гамма-нейтронного излучения индуцирует в метафазах второго и третьего митозов большее число ППК.
Известно, что изменение численного содержания хромосом под влиянием анеогенных факторов является следствием не только повреждения хромосом, но влиянием этих факторов на структуру митотического веретена, в состав которого входят белково-липидные комплексы. В этой связи представляло интерес оценить эффективность гамма-нейтронного излучения различной мощности в повреждении мембранных структур.
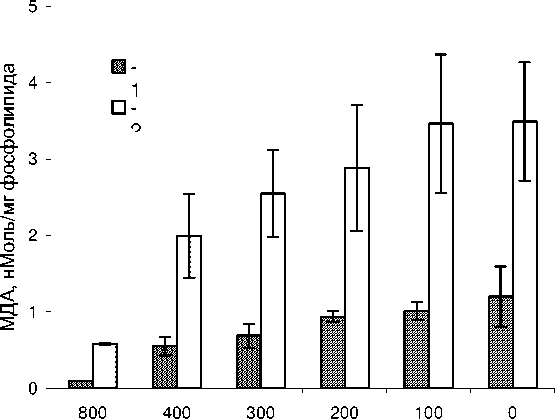
Для исследования радиационной пероксидации липосом при различных мощностях дозы смешанного гамма-нейтронного излучения реактора БАРС-6 в облученных суспензиях липосом определяли содержание малондиальдегида (МДА), который является одним из основных конечных продуктов перекисного окисления липидов. На рис. 5 представлены результаты анализа появления МДА в облученных суспензиях липосом. Полученные результаты свидетельствуют о том, что гамма-нейтронное излучение реактора БАРС-6 увеличивает содержание МДА в липосомах, однако при облучении образцов на расстояниях менее 200 мм от реактора процесс об- разования МДА выходит на плато. Как следует из полученных данных, при одних и тех же поглощенных дозах непрерывное облучение суспензии липосом в течение 60 минут в среднем в 3,8 раза более эффективно увеличивает содержание МДА в липосомах по сравнению с режимом облучения в виде одиночного импульса длительностью 100 мкс.
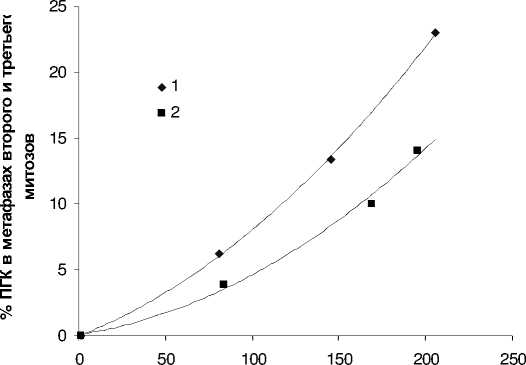
число аберраций на 100 метафаз первого митоза
Рис. 4. Содержание полиплоидных клеток (ППК), %, в метафазах второго и третьего митозов при различных уровнях аберраций хромосом в метафазах первого митоза.
1 – непрерывный режим облучения; 2 – облучение одиночным импульсом. Абсцисса – число аберраций хромосом на 100 метафаз первого митоза. Ордината – % ППК в метафазах второго и третьего митозов.
расстояние от реактора, мм
Рис. 5. Радиолиз суспензии липосом смешанным гамма-нейтронным излучением различной мощности дозы.
-
1 – облучение в режиме одиночного импульса; 2 – непрерывный режим облучения. Абсцисса – расстояние от реактора, мм. Ордината – концентрация малондиальдегида, нМоль на 1 мг фосфолипида.
Дальнейшее исследование влияние мощности дозы на радиолиз водных растворов был проведен с использованием ферросульфатного раствора Фрике. Полученные данные показали, что облучение раствора ферросульфата увеличивает концентрацию Fe(3+) за счет радиационного окисления Fe(2+). Степень радиационного окисления Fe(2+) увеличивается по мере приближения образцов к реактору, однако оба режима облучения при одних и тех же поглощенных дозах имеют одинаковую эффективность в отношении изменения этого показателя. Аналогичные результаты были получены и при исследовании влияния гамма-нейтронного излучения БАРС-6 на появление в облученных растворах БСА белковых перекисей (ROOH). Сравнительный анализ растворов БСА, облученных в режиме одиночного импульса и в непрерывном режиме, свидетельствует о том, что оба режима облучения на одних и тех расстояниях от реактора практически с одинаковой эффективностью увеличивают концентрацию перекисей белков в растворах БСА.
Результаты, полученные при анализе суспензий липосом, растворов ферросульфата и БСА, свидетельствуют о том, что изменение мощности дозы гамма-нейтронного излучения по-разному влияет на радиолиз исследуемых систем. При малых интенсивностях излучения радикальные продукты, диффундирующие в объем раствора из ионизационных треков, практически все перехватываются растворенными соединениями и не рекомбинируют, поскольку не достигают соседних треков. При больших мощностях дозы треки располагаются достаточно близко друг от друга и, несмотря на присутствие в растворе перехватчиков, часть радикалов достигают соседних треков и рекомбинируют. Можно выделить несколько факторов, способствующих снижению эффективности радиационной пероксидации липосом при высоких мощностях дозы смешанного гамма-нейтронного излучения. Одной из причин снижения эффективности окислительной модификации липосом при облучении их суспензий в режиме одиночного импульса может служить концентрационная гетерогенность фосфолипидных эмульсий, что повышает вероятность рекомбинации радикальных продуктов в микрообъемах воды, находящихся вне ли-посомных структур. Известно, что кислород существенно ускоряет процессы радиационной пероксидации липидов. Поэтому нельзя исключить участия в снижении эффективности повреждения фосфолипидов липосом при высоких мощностях доз гамма-нейтронного излучения и радиационного уменьшения концентрации кислорода, которое не восполняется за счет диффузионных процессов во время проведения облучения одиночным импульсом длительностью 100 мкс.
Полученные данные свидетельствуют о дифференциальной чувствительности изученных показателей к повреждающему действию смешанных гамма-нейтронных излучений различной мощностью дозы. Повышенная эффективность цитогенетического действия импульсного гамма-нейтронного излучения на костный мозг мышей (рис. 1, 2) может быть связана с естественной гетерогенностью клеточной популяции кариоцитов (табл. 2), находящихся на различных стадиях клеточного цикла, и увеличением выхода аберраций хромосом в этих клетках за счет ингибирования репарационных процессов при импульсном режиме облучения. В тоже время полученные нами данные свидетельствуют о том, что непрерывный режим облучения более эффективен в повышении уровня полиплоидизации клеток (рис. 4) и нарушении мембранных структур (рис. 5).
Полученные нами результаты позволяют сделать вывод о том, что импульсный режим облучения более эффективен в повреждении генетического аппарата делящихся клеток и в то же время менее эффективен в индукции полиплоидных клеток и повреждении мембранных структур. Увеличенная степень полиплоидизации клеток, повышающая риск возникновения вторичного канцерогенеза, и повышенная пероксидация липидов может приводить к развитию постлучевых повреждений в нормальных тканях. В этой связи проведенные нами исследования дают основание полагать, что использование для лучевой терапии импульсных источников гамма-нейтронного излучения позволит повысить поражаемость генетического аппарата делящихся злокачественных клеток и снизить риск появления побочных эффектов воздействия излучения, связанных с развитием повреждения мембранных структур нормальных тканей и развитием у них опасности возникновения отдаленных канцерогенных эффектов.
Работа выполнена при финансовой поддержке РФФИ и Правительства Калужской области (проект 04-04-97240).
Список литературы Молекулярные и клеточные механизмы действия смешанного гамма-нейтронного излучения различной мощности дозы
- Hall E., Brenner D. The dose-rate effect revisited: radiobiological considerations of importance in radiotherapy//Radiation Oncology Biol. Phys. -1991. -V. 21. -P. 1403-1414.
- Цыб А.Ф., Ульяненко С.Е., Мардынский Ю.С. и др. Нейтроны в лечении злокачественных новообразований. Научно-методическое пособие. -Обнинск: МРНЦ РАМН, 2003. -110 с.
- Даренская Н.Г., Козлова Л.Б., Акоев И.Г. и др. Относительная биологическая эффективность излучений. Фактор времени облучения. -М.: Атомиздат, 1968. -376 с.
- Прохоров Ю.А., Кононов В.И., Кувшинников В.И. и др. Дозиметрия на импульсном реакторе БАРС-6//Атомная энергия. -1998. -Т. 85, № 5. -С. 391-397.
- Поздышкина О.В., Севанькаев А.В., Обатуров Г.М. Сравнительное изучение закономерностей образования аберраций хромосом в культуре лимфоцитов человека при импульсном и пролонгированном нейтронном облучении//Радиационная биология. Радиоэкология. -2000. -Т. 40, № 6. -С. 251-255.
- Филимонов А.С., Обатуров Г.М. Математическое моделирование радиобиологических эффектов при действии стационарного и импульсного нейтронного излучения//Атомная энергия. -1998. -Т. 85, № 5. -С. 396-400.
- Рябченко Н.И., Ульяненко С.Е., Антощина М.М. и др. Действие смешанного у-нейтронного излучения с различной мощностью дозы на содержание клеток в тимусе и хромосомы костного мозга мышей и лимфоцитов человека//Радиационная Биология. Радиоэкология. -2005. -Т. 45, № 5. -С. 592-598.
- Цыб Т.С., Комарова Е.В., Потетня В.И. и др. Биологическая эффективность импульсного и непрерывного нейтронного излучения для клеток дрожжей Sacharamyces//Радиационная биология. Радиоэкология. -2001. -Т. 41, № 3. -С. 291 -295.
- Корякина Е.В., Севанькаев А.В., Потетня В.И., Потетня О.И. Цитогенетическое действие излучения импульсного реактора БАРС в режиме непрерывного и однократного импульсного воздействия со сверхвысокой мощностью дозы на лимфоциты человека//Радиационная Биология. Радиоэкология. -2005. -Т. 45, Вып. 4. -С. 405-411.
- Ульяненко С.Е., Ротт Г.М., Кузнецова М.Н. и др. Выживаемость мышей и содержание металлотионеинов в их печени и почках как критерий оценки воздействия импульсного нейтронного излучения//Радиационная биология. Радиоэкология. -2000. -Т. 40, № 4. -С. 396-400.
- Рябченко Н.И., Ульяненко С.Е., Рябченко В.И. и др. Радиолиз липосом, растворов ферросульфата и альбумина смешанным у-нейтронным излучением с различной мощностью дозы//Радиационная Биология. Радиоэкология. -2005. -Т. 45, № 5. -С. 571-573.
- Рябченко Н.И., Антощина М.М., Насонова В.А. и др. Аберрации хромосом в лимфоцитах человека при различной продолжительности культивирования после облучения//Радиационная Биология. Радиоэкология. -2004. -Т. 44, № 2. -С. 146-150.
- Севанькаев А.В. Радиочувствительность хромосом лимфоцитов человека в митотическом цикле. -М.: Энергоатомиздат, 1987. -158 с.
- Дубинин Н.П. Потенциальные изменения в ДНК и мутации. -Молекулярная цитогенетика. -М.: Наука, 1978.-246 с.
- Li R., Sonik A., Stindl R. et al. Aneuploidy vs. gene mutation hypotasis of cancer: Resend study claims mutation but is found to support aneuploidy//Proc. Natl. Acad. Sc. USA. -2000. -V. 97, N 7. -P. 3236-3241.
- Olaharski A.J., Sotekr R., Solorza-Luna G. et al. Tetraploidy and chromosomal instability are early events during cervical carcinogenesis//Carcinogenesis. -2006. -V. 27, N 2. -P. 337-343.
