Мониторинг интродукционных популяций степных видов растений в условиях экспериментальной степи

Автор: Ибатулина Ю.В.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 3 т.10, 2016 года.
Бесплатный доступ
В условиях экспериментальной степи определено положительное влияние ранневесенних палов на развитие интродукционных популяций боль- шинства модельных видов растений, в том числе и эдификаторов степных фито- ценозов в результате исследования влияния на их динамику антропогенных фак- торов (сенокошение, выжигание), что позволило его рекомендовать для даль- нейших исследований при разработке комплекса мероприятий для эффективно- го регулирования развития степных экосистем.
Интродукционная популяция возрастная, пространственная, виталитетная структура плотность агростепь
Короткий адрес: https://sciup.org/148314572
IDR: 148314572 | УДК: 581.526.53:581.55:
Текст научной статьи Мониторинг интродукционных популяций степных видов растений в условиях экспериментальной степи
Охрана редких видов растений в составе искусственных фитоценозов предусматривает максимальное следование условиям бессрочного функционирования естественных экосистем, в том числе поддержание ксерофит-ности условий существования, включая и формы ресурсопользования (в том числе и отчуждение надземной фитомассы, накопление которой чревато перерождением исходной, природной модели сообщества). Изменение условий существования может привести не только к выпадению раритетных видов из состава искусственных фитоценозов, но и к смене типа растительности. Поскольку искусственные растительные степные сообщества рассматривают как полукультуру – агростепи (Дзыбов, 2001; Остапко, Ибату-лина, 2008; Купрюшина, Остапко, 2011; Приходько и др., 2013; Nazarenko, 2013), то она нуждается в постоянном осуществлении регуляторных мероприятий, обеспечивающих устойчивость её организации и поддерживающих развитие в необходимом направлении, а это не только удаление некоторых видов растений, или сокращение численности их популяций, периодический подсев семян как широко распространенных степных видов, так и охраняемых. Необходима разработка комплекса мероприятий по рациональ- ному использованию степных фитоценозов, сочетающего в себе факторы, способствующие формированию ксерофитной степной растительности – регулируемый выпас, контролируемые палы, умеренная рекреация, косьба (Кондратюк, Чуприна, 1992; Milton, Dean, Klotz, 1997; Valee, 2004; Горбунов и др., 2008; Остапко, Ибатулина, 2008; Назаренко, 2012, 2013).
Искусственные степные фитоценозы в Донецком ботаническом саду создавались с 1968 г. С.Н. Зиман, Д.С. Ивашиным, Е.Н. Кондратюком, Т.Т. Чуприной – были разработаны и успешно испытаны в длительном интродукционном эксперименте несколько методов создания натурных моделей разнотравно-типчаково-ковыльных степей: эдифи-каторно-биоморфологический поэтапный, комбинированный биоморфологический и эдификаторно-ценотический (Кондратюк, Чуприна, 1992). Учёт особенностей структурно-функциональной организации интродукционных популяций позволяет объективно оценивать не только их устойчивость, но и успешность формирования натурных моделей растительных сообществ в целом, дает возможность получать достаточно полное представление о процессах, которые происходят в искусственных фитоценозах,
и прогнозировать состояние агростепей в будущем. Изменения популяционных параметров растений позволят определить наиболее подходящие условия поддержания организации степных фитоценозов (Остапко, Хархота, 1994; Емельянов и др., 2007; Кагало, 2007; Ибатулина, 2013 а, 2013 б). Исследования проводятся в условиях экспериментальной степи, поскольку она максимально подобна природным степным растительным сообществам и для неё характерны аналогичные процессы развития (Кондратюк, Чуприна, 1992; Горбунов и др., 2008; Купрюшина, Остапко, 2011; Назаренко, 2013).
Район исследований
Исследования проводились на территории Донецкого ботанического сада. Комплексная экспозиция расположена на водоразделе балки Богодуховская, увлажнялась только атмосферными осадками. Почвы – обыкновенные чернозёмы на лессовидном карбонатном суглинке. Первый экспериментальный участок (0,04 га) заложен пересадкой 360 дернин размером 30 х 30 см в 1968 г. Растительные сообщества второго экспериментального участка (0,07 га) создавались с 1972 г. на базе питомника размножения степного разнотравья (посев 1968 г.) и видов рода Stipa (посев 1971 г.) комбинированным способом: высадкой дернин, отдельных особей и подсевом семян степных видов растений. Основой для третьего экспериментального участка послужил питомник размножения степных видов растений (0,5 га). Питомник создавался на участке, прилегающем, с одной стороны, к первому экспериментальному участку, с другой – ко второму, площадью 0,5 га (Кондратюк, Чуприна, 1992). С 2002 г. участок был оставлен для самозарастания, что впоследствии привело к формированию разнотравнозлакового фитоценоза, в котором обильно представлены ксеромезофитные и мезоксе-рофитные корневищные злаки и степные виды разнотравья. Флористическое богатство сообществ на I экспериментальном степном участке составляет 103 вида, II – 236 видов, III – 264 вида.
На формирование существующих искусственных растительных сообществ определенное влияние оказывали антропогенные факторы: стихийный выпас мелкого рогатого скота до 2010 г., проводилось периодическое скашивание некоторых участков степи, происходили стихийные пожары и умеренное или сильное вытаптывание из-за рекреационной нагрузки. То есть, в некоторой степени были воспроизведены факторы, которые участвуют в развитии естественной степной растительности. Наблюдения показали, что скашивание не является особенно эффективным, к тому же, в отличие от выпаса и выжигания, оно является не природным, чуждым степным экосистемам. Ежегодное выкашивание только замедляет скорость преобразования фитоценозов в сообщества другого типа (Зауголь-нова, 1994; Остапко, Ибатулина, 2008; Приходько и др., 2013). Поэтому проблема поиска результативных комплексных методов поддержания, регулирования направления развития степных растительных сообществ, в которых сенокошение являлось бы вспомогательным фактором, остаётся открытой и не теряет своей актуальности. В этом направлении в настоящее время в ботаническом саду осуществляется эксперимент на базе искусственных степных фитоценозов по изучению воздействия различных комбинаций антропогенных факторов на динамику интродукционных популяций. Режим природопользования на первом экспериментальном степном участке – ежегодный укос, с дополнительной уборкой сухих растительных остатков в ранневесенний период, на втором – ежегодный укос, с ранневесенним выжиганием (периодичность – раз в два года). Третий экспериментальный участок – режим содержания трех видов: начиная с 2003 г.: ежегодный укос с дополнительной уборкой сухих растительных остатков в ранневесенний период; ежегодное проведение ранневесеннего (конец марта – начало апреля) пала; режим полного невмешательства, подобный абсолютному заповеданию.
Объекты и методы
Цель исследования – определить эффективность влияния различных антропогенных факторов на состояние интродукционных популяций широко распространенных и редких видов в натурных моделях растительных сообществ.
Объектами явились интродукционные популяции как охраняемых, так и широко распространенных степных видов, которые являются эдификаторами степных фитоценозов, а также доминантами или субдоминантами, ассектаторами в сообществах разнотравно-типчаково-ковыльной настоящей степи на чернозёмах и принадлежат к разным жизненным формам в соответствии с классификацией, разработанной С.Н. Зиман (1976): Stipa capillata L., S. lessingiana Trin. et Rupr., S. grafiana Steven , Festuca valesiaca Gaudin, F. rupicola Heuff ., Phleum phleoides (L.) H. Karst., Elytrigia stipifolia (Czern. ex Nevski) Nevski, Veronica sclerophylla Dubovik, Filipendula vulgaris Moench, Fragaria viridis Duchesne, Galium ruthenicum Willd., Paeonia tenuifolia L., Chrysocyathus vernalis L., Сh. wolgensis Steven, Pulsatilla bohemica (Scalický) Tzvelev, Thalictrum minus L., Trifolium caucasicum Tausch, Haplophyllum ciliatum Griseb., Salvia stepposa Des.-Shost., S. tesquicola Klokov et Pobed. , Marrubium praecox Janka , Nepeta parviflora M. Bieb. , Stachys transsilvanica Schur, Jurinea arachnoidea Bunge, Tulipa gesneriana L., Allium firmotunicatum Fomin.
Выделение возрастных групп проводилось в соответствии с онтогенетическими особенностями конкретных видов растений (Борисова, Попова, 1971; Заугольнова, 1974; Берко, 1976; Зиман, 1976; Воронцова, Зауголь-нова, 1978; Заугольнова, 1994), изучение возрастной структуры осуществлялось по общепринятым методикам (Уранов, Смирнова, 1969; Уранов, 1975; Жукова и др., 1989; Гиляров, 1990; Злобин, 1989, 2009). Виталитет-ную структуру изучали по методике Ю.А. Злобина (Злобин, 1989, 2009). По уровню виталитета особи подразделяли на три основных класса: "а" – высший, "b" – средний, "с" – низший. После установления виталитета особей оценку качества ценопопуляций осуществляли по долевому участию особей разного виталитета в составе популяций. По качеству популяции подразделяли на три основных типа:
процветающие – Q равновесные – Q депрессивные – Q
( a + b )
( a + b )
( a + b )
> с ,
= с ,
< с ,
где Q – индекс качества популяции; "a", "b", "c", – соответствующие частоты особей высшего, среднего и низшего классов. Все оценки качества особей были сопряжены с конкретным возрастным состоянием. Для выявления ключевых признаков был применён факторный анализ. Виталитетное состояние определяли по следующим морфометри- ческим параметрам: высота растений, диа- метр дерновин, количество генеративных побегов. Основные типы пространственного распределения особей определялись согласно принципам, изложенным в работе А.М. Гилярова (1990). Для установления типа размещения особей по площади использовали отношение дисперсии к среднему: 5—, где ст2 m
– дисперсия, m – средняя. Если показатель около единицы, то исследуемое распределение – случайное, если больше – контагиозное, если меньше – регулярное.
Фитоценотическая позиция устанавливалась как доминант (по обилию и проективному покрытию) или ассектатор. Средняя плотность определялась числом особей или счетных единиц на 1 м2. В качестве счётной единицы для злаков использовали особь – проростки и ювенильные, имматурные, виргиниль-ные, молодые генеративные растения; клон – зрелые, старые генеративные растения, то есть совокупность партикул, возникших вегетативным путём из семенной особи (обычно, такую счётную единицу применяют в том случае, если клон характеризуется компактностью и воспринимается как единый источник фитогенного воздействия); партикулы – субсенильные и сенильные растения, если связь между частями дерновины не сохраняется. Для других видов растений использовали особь, клон, парциальный побег, парциальный куст. Выбор зависел от того, к какой жизненной форме относится вид (Жукова и др., 1989; Злобин, 1989). Уровни охраны видов растений приведены согласно литературным источникам (Губанов, 2002; Дідух, 2009; Остапко, 2010).
Результаты
В настоящее время на первом экспериментальном степном участке в ДБС существует два степных растительных сообщества, которые относятся к ассоциациям Fes-tucetum ( valesiacae ) stiposum ( lessingianae ), Festucetum ( valesiacae ) bromopsiosum ( ri-pariae ), на втором – к ассоциациям Fes-tucetum ( valesiacae ) elytrigiosum ( repentis ) и Festucetum ( valesiacae ) viciosum ( tenuifoliae ).
Ассоциации Festucetum (valesiacae) elytrigiosum (repentis), Festucetum (valesiacae) viciosum (tenuifoliae) в результате высокого обилия длиннокорневищных луговых эвримезофитов – Elytrigia repens (L.) Nevski, Bromopsis inermis (Leyss.) Holub (проективное покрытие 20-10%), луговостепного ксеромезофита Poa angustifolia L. (покрытие 5%) и короткокорневищного элемента – Bromopsis riparia (Rehmann) Holub (покрытие 10-20%), могут быть звеньями резерватогенной сукцессии, следствием мезо-фитизации степного растительного покрова, которая направлена в сторону формирования луговых сообществ из-за повышенного увлажнения. В сообществе ассоциации Fes-tucetum (valesiacae) stiposum (lessingianae) соэдификатором и субдоминантом является эвксерофит Stipa lessingiana (проективное покрытие 20%), обилие которого во второй ассоциации значительно снижается (покрытие 5%). Длиннокорневищные злаки играют небольшую фитоценотическую роль (ассек-таторы второго-третьего рангов). Среди разнотравья доминируют Salvia nutans L., Ga-latella dracunculoides (Lam.) Ness, Thalictrum minus. Подобная ситуация наблюдалась во втором фитоценозе. Видовая насыщенность на 1 м2 составляет 13-18. Общее проективное покрытие составляет 80%. В Festucetum (valesiacae) elytrigiosum (repentis) и Festucetum (valesiacae) viciosum (tenuifoliae) виды разнотравья составляют смешанный травостой, среди них доминирующими являются Trifolium ambiguum M. Bieb., T. montanum L., Plantago lanceolata L., P. urvillei Opiz, Galatella dracunculoides, Thalictrum minus, Fragaria viridis, Vicia tenuifolia L. В очень малом обилии представлены Stipa lessingiana, S. capillata, S. grafiana. Видовая насыщенность на 1 м2 в ассоциации Festucetum (valesiacae) ely-trigiosum (repentis) – 15-29, на 1 м2, в ассоциации Festucetum (valesiacae) viciosum (tenuifoliae) – 11-22. Общее проективное покрытие составляет 95-100%.
В ассоциации Festucetum (valesiacae) ely-trigiosum (repentis) доминируют короткокорневищные растения, в Festucetum (vale-siacae) viciosum (tenuifoliae) – длиннокорневищные. Это может быть обусловлено большей мезофитизацией второго сообщества. Различия в эколого-фитоценотических условиях, установившиеся в различных сообществах, привели к формированию двух типов биоморф у Galium ruthenicum. В природных условиях в фитоценозах настоящей разнотравно-типчаково-ковыльной и типчаковоковыльной степи отмечали преобладание компактной формы. В фитоценозах, в которых отмечены процессы олуговения, в частности в условиях абсолютно заповедного режима, преобладает длиннокорневищная форма, что так же отмечено в условиях искусственной степи. Фитоценозы второго участка характеризуются гораздо большей степенью мезофитизации, они приближаются к разно-травно-длиннокорневищно-злаковым сообществам, которые приходят на смену разнотравно-типчаково-ковыльным и типчаковоковыльным растительным сообществам при абсолютно заповедном режиме. Тем не менее, эти фитоценозы в настоящем остаются устойчивыми, что подтверждает и структура популяций видов растений (не только эдифи- каторов, но и сопутствующих видов (Купрю-шина, Остапко, 2011; Приходько и др., 2013).
Исследования показали, что положение популяций некоторых редких и исчезающих видов растений в искусственных степных фитоценозах является непрочным из-за их низкой плотности, неполночленности возрастного состава и угнетенного состояния растений (табл. 1, 2). Поэтому для поддержания эколого-фитоценотических условий, которые отвечают требованиям степных видов растений, необходимо не только постоянное применение комплекса агротехнических мероприятий, но и его усовершенствование. В це- лом, интродукционные популяции большинства изученных модельных степных видов как широко распространенных, так и раритетных, занимают прочное положение в сложившихся искусственных сообществах, являются саморегулирующимися системами и не зависят от заноса диаспор. Наиболее устойчивыми являются популяции видов-эдификаторов. Для изучения динамики развития интродукционных популяций плотнодерновинных злаков в искусственных степных фитоценозах в качестве модельных видов использованы: Festuca rupicola, F. valesiaca и Stipa lessingiana.
Таблица 1. Плотность, возрастная структура интродукционных популяций раритетных видов в искусственных степных фитоценозах на І и ІІ экспериментальных участках
Density, age structure of the introduced populations of rarity species in the artificial steppe phytocenoses in I and II experimental plots
|
* |
Вид и уровень охраны, статус |
Плотность особь / м2 |
Участие онтогенетических групп, % от общего числа особей |
||||||||
|
рl |
j |
im |
v |
g 1 |
g 2 |
g 3 |
ss |
s |
|||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
«-Н |
Chrysocyathus vernalis , региональный уровень охраны, уязвимый |
1,4 ± 0,5 |
0 |
0 |
9,3 |
14,8 |
21,3 |
41,7 |
8,3 |
4,6 |
0 |
|
Chrysocyathus wolgensis, региональный уровень охраны, уязвимый |
одиночные зрелые генеративные особи |
||||||||||
|
Pulsatilla bohemica региональный уровень охраны, уязвимый |
1,1 ± 0,4 |
0 |
0 |
0 |
10,0 |
22,5 |
57,5 |
10,0 |
0 |
0 |
|
|
Paeonia tenui-folia , Е, U, уязвимый |
1,3 ± 0,6 |
0 |
3,5 |
8,4 |
18,2 |
28,1 |
34,3 |
3,5 |
2,1 |
1,9 |
|
|
Haplophyllum ciliatum, региональный уровень охраны, редкий |
одиночные зрелые генеративные особи |
||||||||||
|
Stipa capillata, U, редкий |
2,0 ± 0,7 |
0 |
0 |
0 |
5,1 |
24,2 |
46,5 |
22,2 |
2,0 |
0 |
|
Продолжение таблицы 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
|
Stipa lessingiana, U, редкий |
5,6 ± 0,6 |
0 |
0 |
8,1 |
9,2 |
21,6 |
36,7 |
16,3 |
8,1 |
0 |
||
|
Ct |
Chrysocyathus vernalis, региональный уровень охраны, уязвимый |
1,3 ± 0,7 |
0 |
0 |
5,9 |
6,9 |
12,8 |
39,1 |
28,0 |
7,3 |
0 |
|
|
Pulsatilla bohemica, региональный уровень охраны, уязвимый |
одиночные зрелые генеративные особи |
|||||||||||
|
Paeonia tenuifolia, Е, U, уязвимый |
3,8 ± 1,9 |
0 |
3,2 |
15,1 |
24,6 |
20,4 |
25,0 |
7,5 |
4,2 |
0 |
||
|
Haplophyllum ciliatum, региональный уровень охраны, редкий |
одиночные зрелые генеративные особи |
|||||||||||
|
Salvia stepposa, региональный уровень охраны, редкий |
1,5 ± 0,5 |
0 |
3,5 |
5,5 |
18,4 |
18,4 |
36,7 |
11,2 |
6,3 |
0 |
||
|
Stipa capillata, U, редкий |
2,6 ± 0,6 |
0 |
0 |
10,5 |
12,4 |
16,7 |
34,2 |
18,7 |
7,3 |
0 |
||
|
Stipa lessingiana, U, редкий |
2,4 ± 0,5 |
0 |
0 |
14,9 |
10,6 |
13,5 |
33,3 |
27,7 |
0 |
0 |
||
|
rf> |
Trifolium caucasicum, региональный уровень охраны, редкий |
4,8 ± 0,9 |
0 |
2,8 |
7,8 |
10,7 |
20,6 |
30,7 |
24,1 |
3,3 |
0 |
|
|
Paeonia tenuifolia, Е,U, уязвимый |
3,8 ± 1,4 |
0 |
2,7 |
10,8 |
16,8 |
17,1 |
30,2 |
18,5 |
3,8 |
0 |
||
Окончание таблицы 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
сП |
Tulipa gesneriana, U, под угрозой исчезно вения |
одиночные зрелые генеративные особи |
|||||||||
|
Allium firmotuni-catum , региональный уровень охраны, редкий |
|||||||||||
|
Stipagrafiana, U, редкий |
1,3 ± 0,4 |
0 |
0 |
11,5 |
13,5 |
15,4 |
17,4 |
32,7 |
9,5 |
0 |
|
|
Stipa lessingiana, U, редкий |
2,4 ± 0,9 |
0 |
0 |
11,3 |
20,8 |
27,1 |
25,0 |
15,8 |
0 |
0 |
|
|
Trifolium caucasicum, региональный уровень охраны, редкий |
4,6 ± 0,6 |
0 |
1,9 |
6,9 |
7,9 |
16,8 |
20,8 |
36,6 |
7,4 |
1,5 |
|
|
Tulipa gesne-riana, U, под угрозой исчезновения |
одиночные зрелые генеративные особи |
||||||||||
|
Allium firmotuni-catum , региональный уровень охраны, редкий |
|||||||||||
|
Elytrigia stipifolia, W, E, U, редкий |
|||||||||||
|
Stipagrafiana, U, редкий |
1,2 ± 0,7 |
0 |
0 |
6,9 |
19,0 |
23,1 |
37,2 |
10,0 |
3,8 |
0 |
|
|
Stipa lessingiana, U, редкий |
1,8 ± 0,3 |
0 |
0 |
13,2 |
21,3 |
25,8 |
31,3 |
8,3 |
0 |
0 |
|
Примечание: * номер ассоциации, 1 – Festucetum ( valesiacae ) stiposum ( lessingianae ), I участок; 2 – Fes-tucetum ( valesiacae ) bromopsiosum ( ripariae ) I участок; 3 – Festucetum ( valesiacae ) elytrigiosum ( repentis ) II участок; 4 – Festucetum ( valesiacae ) viciosum ( tenuifoliae ) II участок
Таблица 2. Виталитетная структура и пространственное размещение особей в интродукционных популяциях некоторых раритетных видов в искусственных степных фитоценозах на І и ІІ экспериментальных участках
Vitality structure and individual spacing within the introduced populations of some rarity species in the artificial steppe phytocenoses in I and II experimental plots
|
* |
Вид и уровень охраны, статус |
Размещение |
Виталитетные классы |
Q |
||
|
c |
b |
a |
||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
i-Ч |
Chrysocyathus vernalis , региональный уровень охраны, уязвимый |
контагиозное |
0,440 |
0,280 |
0,240 |
0,260 < c |
|
Chrysocyathus wolgensis, региональный уровень охраны, уязвимый |
единичные угнетённые особи |
|||||
|
Pulsatilla bohemica, региональный уровень охраны, уязвимый |
0,360 |
0,320 |
0,320 |
0,320 < с |
||
|
Paeonia tenuifolia, Е, U, уязвимый |
0,320 |
0,400 |
0,280 |
0,340 > с |
||
|
Haplophyllum ciliatum, региональный уровень охраны, редкий |
единичные угнетённые особи |
|||||
|
Stipa capillata, U, редкий |
0,200 |
0,520 |
0,280 |
0,400 > с |
||
|
Stipa lessingiana, U, редкий |
0,440 |
0,200 |
0,360 |
0,280 < с |
||
|
|
Chrysocyathus vernalis , региональный уровень охраны, уязвимый |
контагиозное |
0,360 |
0,200 |
0,400 |
0,300 < с |
|
Pulsatilla bohemica, региональный уро вень охраны, уязвимый |
единичные угнетённые особи |
|||||
Продолжение таблицы 2
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
n |
Paeonia tenuifolia, Е, U, уязвимый |
контагиозное |
0,320 |
0,280 |
0,400 |
0,340 > c |
|
Haplophyllum ciliatum , региональный уровень охраны, редкий |
единичные угнетённые особи |
|||||
|
Stipa capillata, U, редкий |
0,440 |
0,400 |
0,160 |
0,420 > с |
||
|
Stipa lessingiana, U, редкий |
0,400 |
0,320 |
0,280 |
0,300 < с |
||
|
m |
Trifolium caucasicum, региональный уровень охраны, редкий |
контагиозное |
0,320 |
0,360 |
0,320 |
0,340 > c |
|
Paeonia tenuifolia, Е, U, уязвимый |
0,320 |
0,280 |
0,400 |
0,340 > c |
||
|
Tulipagesneriana, U, под угрозой исчезновения |
единичные угнетённые особи |
|||||
|
Allium firmotunicatum, региональный уровень охраны, редкий |
||||||
|
Stipagrafiana, U, редкий |
0,320 |
0,400 |
0,280 |
0,340 > с |
||
|
Stipa lessingiana, U, редкий |
0,480 |
0,160 |
0,360 |
0,260 < с |
||
|
Tf |
Trifolium caucasicum, региональный уровень охраны, редкий |
контагиозное |
0,222 |
0,389 |
0,389 |
0,389 > c |
|
Tulipagesneriana, U, под угрозой исчезновения |
единичные угнетённые особи |
|||||
Окончание таблицы 2
|
Allium firmotunicatum, региональный уровень охраны, редкий |
единичные угнетённые особи |
|||||
|
Elytrigia stipifolia, W, E, U, редкий |
||||||
|
Stipagrafiana, U, редкий |
0,400 |
0,280 |
0,320 |
0,340 < с |
||
|
Stipa lessingiana, U, редкий |
0,360 |
0,320 |
0,320 |
0,320 < с |
||
Примечание: * номер ассоциации, 1 – Festucetum ( valesiacae ) stiposum ( lessingianae ), I участок; 2 – Festucetum ( valesiacae ) bromopsiosum ( ripariae ) I участок; 3 – Festucetum ( valesiacae ) elytrigiosum ( repentis ) II участок; 4 – Festucetum ( valesiacae ) viciosum ( tenuifoliae ) II участок
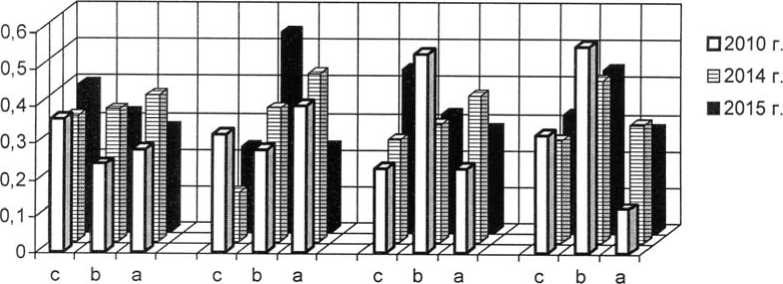
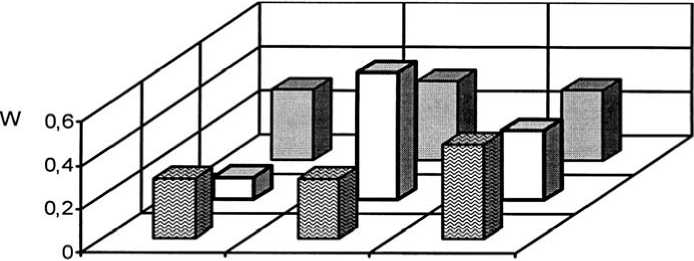
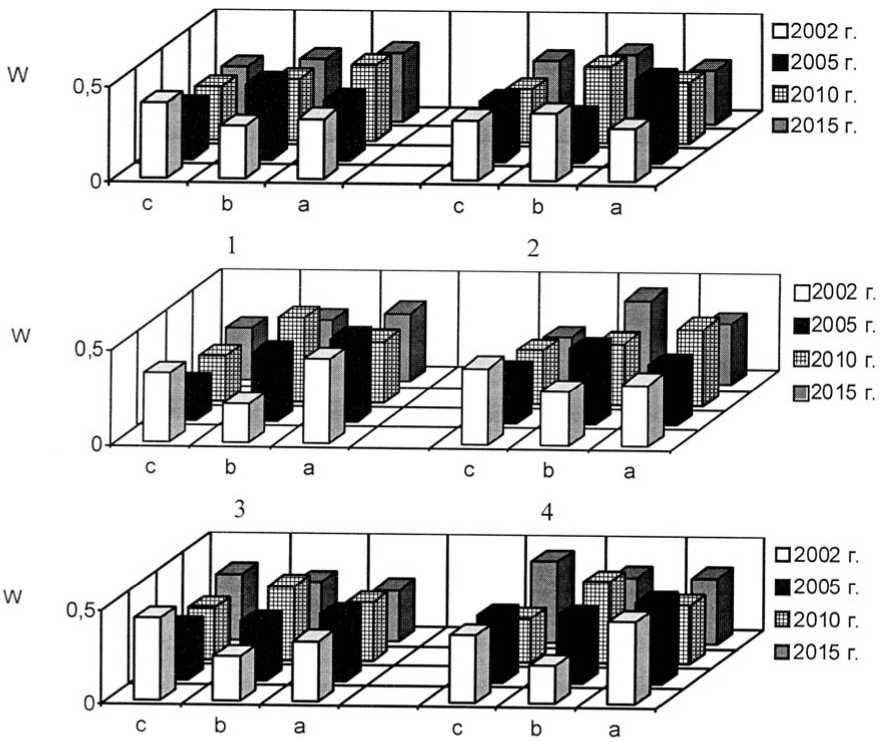
Как уже было обозначено, отличительной чертой I и II экспериментальных степных участков является выраженная мезофи-тизация растительного покрова, но большинство популяций злаков в искусственных фитоценозах можно отнести к устойчивым. На плотность, продолжительность возрастных периодов и другие процессы в популяциях оказывает сильное воздействие жизненное состояние особей. В 2015 г. популяции плотнодерновинных злаков относились к депрессивному и процветающему витали-тетным типам (рис. 1-3).
w
Ухудшение жизненного состояния особей может отрицательно сказаться на их генеративной функции, что впоследствии приводит к снижению эффективности семенного возобновления и нарушению регулярности замещения поколений. Результатом этого является неполночленность возрастного состава, что ведёт к снижению устойчивости самих популяций. Это характерно для популяций Stipa lessingiana и S. capillata. Отсутствие молодых растений из-за нарушения цикличности семенного возобновления приводит к нарушению саморегуляции популяций.
1 2 3 4
Рис. 1. Динамика виталитетной структуры интродукционных популяций Festuca valesiaca Gaudin в искусственных степных фитоценозах: 1 – Festucetum ( valesiacae ) stiposum ( lessingianae ); 2 – Festucetum ( valesiacae ) bromopsiosum ( ripariae ); 3 – Festucetum ( valesiacae ) elytrigiosum ( repentis ); 4 – Festucetum ( valesiacae ) viciosum ( tenuifoliae ); W – встречаемость, "а" – высший класс, "b" – средний класс, "с" – низший класс
Vitality structure development of the introduced populations of Festuca valesiaca Gaudin in the artificial steppe phytocenoses
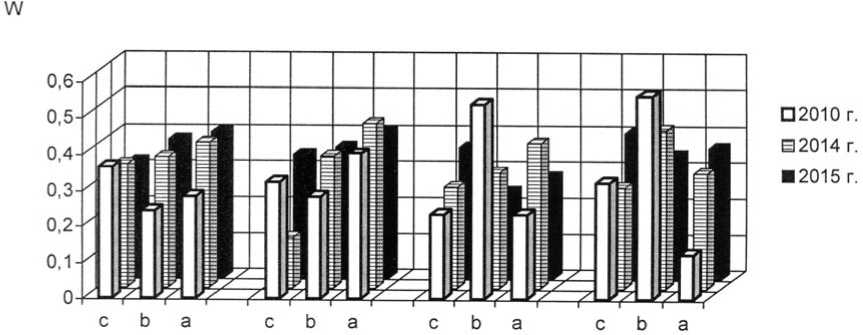
Рис. 2. Динамика виталитетной структуры интродукционных популяций Festuca rupicola Heuff. в искусственных степных фитоценозах: 1 – Festucetum (valesiacae) stiposum (lessingianae) ; 2 – Festucetum (valesiacae) bromopsiosum (ripariae) ; 3 – Festucetum (valesiacae) elytrigiosum (repentis) ; 4 – Festucetum (valesiacae) viciosum (tenuifoliae) ; W – встречаемость, "а" – высший класс, "b" – средний класс, "с" – низший класс
Vitality structure development of the introduced populations of Festuca rupicola Heuff. in the artificial steppe phytocenoses
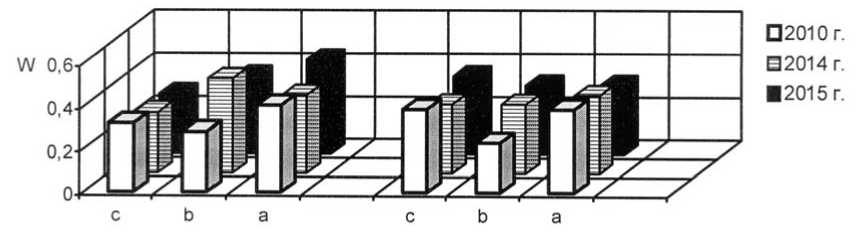
Рис. 3. Динамика виталитетной структуры интродукционных популяций Stipa lessingiana Trin. et Rupr. в искусственных степных фитоценозах: 1 – Festucetum ( valesiacae ) stiposum ( lessingianae ); 2 – Festucetum ( valesiacae ) bromopsiosum ( ripariae ); W – встречаемость, "а" – высший класс, "b" – средний класс, "с" – низший класс
Vitality structure development of the introduced populations of Stipa lessingiana Trin. et Rupr. in the artificial steppe phytocenoses
Плотность популяций степных ценозооб-разователей в искусственных степных растительных сообществах подвержена некоторым колебаниям, но поддерживается практически на одном уровне (табл. 3, на примере интродукционной популяции Festuca valesiaca). Наблюдения за динамикой развития интродукционных популяций злаков в модельных искусственных степных фитоценозах на экспериментальных участках, начиная с 2002 г., показали, что их возрастные спектры остаются левосторонними. Это является результа- том того, что популяции эдификаторов дефинитивные. Возрастная структура интродукционных популяций ценозообразователей испытывает небольшие флюктуации: соотношение возрастных групп не оставалось неизменным, но характер возрастного спектра не менялся. Некоторое омоложение за счёт появления ювенильных особей отмечено в 2003 г., 2008-2010 г., 2015 г. Возрастное развитие происходит медленными темпами и выражается в основном в переходе растений в следующее онтогенетическое состояние.
Таблица 3. Динамика плотности интродукционных популяций эдификаторов степных фитоценозов в искусственных растительных сообществах (особь /м2)
Density development of edificator species of the steppe phytocenoses in artificial plant commnuni-ties
|
Ассоциации |
Вид |
2006 г. |
2010 г. |
2015 г. |
|
Festucetum ( valesiacae ) stiposum ( lessingianae ) |
Festuca rupicola |
8,7±2,4 |
8,7±2,4 |
8,5±2,6 |
|
Festuca valesiaca |
13,9±1,5 |
10,9±1,4 |
11,2±1,4 |
|
|
Stipa lessingiana |
4,5±1,6 |
4,9±1,8 |
5,6±1,4 |
|
|
Festucetum ( valesiacae ) bromopsiosum ( ripariae ) |
Festuca rupicola |
8,5±1,5 |
6,2±1,7 |
6,9±1,7 |
|
Festuca valesiaca |
15,5±1,8 |
13,2±1,6 |
13,8±1,6 |
|
|
Stipa lessingiana |
2,5±0,5 |
3,1±0,9 |
2,4±0,5 |
|
|
Festucetum ( valesiacae ) elytrigiosum ( repentis ) |
Festuca rupicola |
- |
- |
- |
|
Festuca valesiaca |
12,1±2,3 |
11,3±2,1 |
14,7±2,5 |
|
|
Stipa lessingiana |
- |
- |
- |
|
|
Festucetum ( valesiacae ) viciosum ( tenuifoliae ) |
Festuca rupicola |
- |
- |
- |
|
Festuca valesiaca |
8,5±2,7 |
8,4±2,7 |
9,5±2,5 |
|
|
Stipa lessingiana |
Подавляющая часть особей в популяциях Festuca rupicola и F. valesiaca относятся к молодой фракции: молодые вегетативные особи и молодые генеративные растения. Неполно-членность данных популяций, выражающаяся в отсутствии субсенильных и сенильных растений, объясняется не только временным фактором, как результат замедления онтогенеза, но и, возможно, более жёсткими условиями существования, когда особи заканчивают свой жизненный цикл на более ранних этапах развития.
Менее прочное положение в растительном сообществе занимает популяция Stipa lessingiana в ассоциации Festucetum (valesiacae) bromopsiosum (ripariae), что является следствием меньшей ее плотности и полноты возрастного состава, не только из-за отсутствия старой генеративной и постгенеративных групп, но и более молодой части возрастного спектра популяции в результате длительного перерыва в семенном возобновлении, элиминации части молодых растений. Популяции Stipa lessingiana неполночленные молодые, нормальные. Доля имматурных особей за время наблюдений также подвергалась некоторым изменениям либо за счёт поступления особей из группы ювенильных растений, либо за счёт отмирания части особей и перехода в следующее возрастное состояние, что вызвало в некоторых популяциях увеличение количества виргинильных особей. Пополнение групп генеративных особей не оказало существенного влияния на характер возрастных спектров, как и флюктуационные изменения доли особей в молодой части интродукционных популяций. Данные популяции являются дефинитивными, им свойственна флюктуационная и низкоамплитудная динамика плотности. Однонаправленные изменения плотности характерны для сукцес-сивных популяций, к которым ни одна из рассмотренных популяций степных злаков не относится.
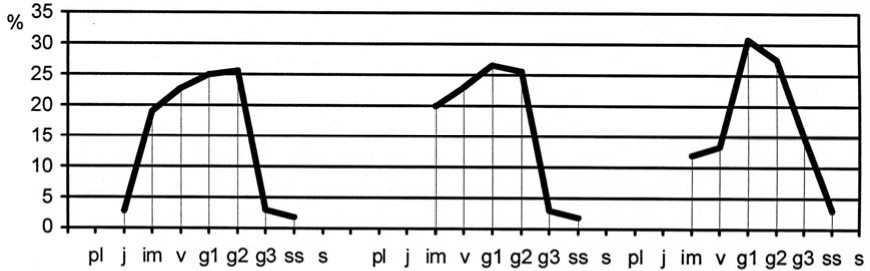
Более интенсивная мезофитизация растительного покрова второго экспериментального участка способствовала формированию неблагоприятных условий произрастания для плотнодерновинных эдификаторов степных фитоценозов. Однако, после проведения экспериментов с палами, развитие популяций Festuca rupicola, F. valesiaca и Stipa lessingianа ускорило свои темпы благодаря улучшению условий существования. Увеличилась не только плотность особей (табл. 4), но и доля участия генеративных растений в возрастном составе интродукционных популяций злаков (рис. 4, на примере интродукционной популяции Festuca valesiaca), а также их мощность, что связано с повышением жизненного уров- ня особей [рис. 1, популяция Festuca valesiaca в ассоциациях Festucetum (valesiacae) bromopsiosum (ripariae), Festucetum (valesiacae) viciosum (tenuifoliae)]. Этот факт связан с тем, что палы угнетают развитие популяций длиннокорневищных луговых и лугово-степных видов, в том числе Elytrigia repens, Vicia tenuifolia, которые характеризуются ранним началом вегетации. Ранневесенние палы существенно повреждают надземные побеги и задерживают развитие растений. Влияние длиннокорневищных видов на плотнодерновинные злаки ослабляется, что способствует развитию их популяций. Отмечено, что преобразование условий существования способствует быстрому изменению виталитетного типа популяций. Ухудшение интродукционных популяций в результате накопления в их составе растений третьего виталитетного класса. Переход на более низкие уровни жизненности может увеличивать продолжительность пребывания растений в отдельных возрастных состояниях. Это повышает устойчивость популяций, так как сохраняется разнообразие их возрастного состава. В 2015 г. отмечался переход части популяций плотнодерновинных злаков к процветающему виталитетному типу (рис. 1-3, на примере интродукционных популяций Fes-tuca valesiaca, Festuca rupicola и Stipa lessingiana), что способствовало упрочнению позиций видов в растительных сообществах и усилению их влияния на среду в результате некоторого улучшения условий существова- условий вызывает изменение жизненности ния.
2010 г.
2014 г.
2015 г.
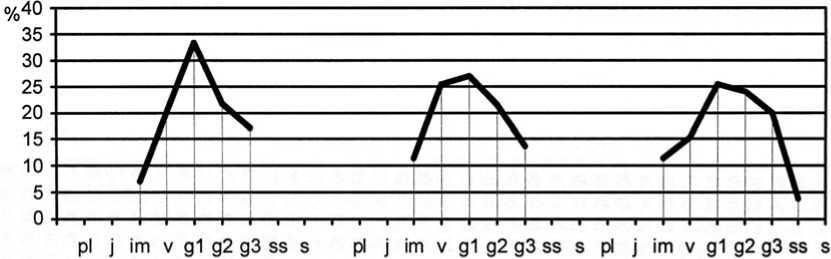
Рис. 4. Динамика возрастной структуры интродукционной популяции Festuca valesiaca Gaudin в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae )
Age structure development of Festuca valesiaca Gaudin introduced population in Festucetum ( vale-siacae ) viciosum ( tenuifoliae ) association
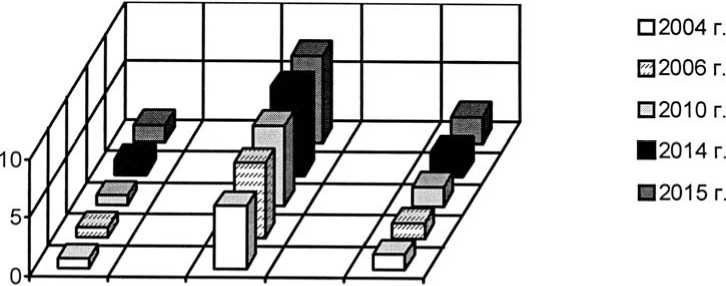
Интродукционные популяции стержнекорневых многолетников характеризуются стабильной возрастной структурой и поддержанием плотности почти на одном уровне. Её величина меняется редко и в популяциях не всех видов, как правило, за счет появления молодых вегетативных растений, часть из которых может погибнуть за вегетационный период. Увеличение численности популяций этих видов связано с появлением потомства в результате семенного размножения в весенние месяцы. В связи с разнообразием факторов, которые влияют на появление и приживание новых растений, чередование подъемов и спадов плотности популяций стержнекорневых растений носит случайный характер. Так, в период наблюдений в структуре популяций Salvia tesquicola, S. nutans, Marrubium praecox, Nepeta parviflora, Jurinea arachnoidea не отмечены заметные изменения и отсутствовали колебания плотности особей, у других – периоды омоложения и старения чередуются (Linum nervosum, Stachys transsіlvanica). Развитие осуществляется медленно, что подтверждает и отсутствие в составе многих популяций этих видов старой генеративной и постгенеративных групп, и выражается в основном в переходе незначительной доли молодых растений в следующее возрастное состояние. Подобное может происходить как из-за увеличения продолжительности онтогенеза в целом, так и отдель- ных его периодов, особенно за счет накопления особей с низким жизненным уровнем, которые способны в результате этого долго находится на определённой стадии развития. Большинство популяций стержнекорневых многолетников являются нормальными, но не имеют полного набора возрастных групп (рис. 5-6, на примере интродукционной популяции Linum nervosum). При этом выпадение возрастных групп правой части возрастного спектра связано с развитием популяции во времени. Их появления можно ожидать по мере старения молодых растений. Но выпадение отдельных возрастных групп может быть связано и с ухудшением условий существования (растения отмирают на более ранней стадии развития). В результате сокращения онтогенеза увеличивается скорость обновления, т.е. замещения поколений, в этих интродукционных популяциях особи могут быстрее достигать генеративного состояния и давать новое потомство. Для всех исследованных интродукционных популяций стержнекорневых видов (Linum austriacum, Marrubium praecox, Nepeta parviflora, Salvia tesquicolа) отмечено очень медленное старение. Только у Linum nervosum, Stachys transsіlvanica осуществилось пополнение популяций за счет особей семенного происхождения (2010 г., 2015 г.), но процесс старения этих популяций в целом преобладает над «омоложением». Большинство популяций этих видов являются депрессивными. На угнетенность указывает и контагиозное размещение растений в пространстве, что позволяет видам существовать в условиях сильного конкурентного влияния со стороны других растений.
2010 г.
2014 г.
2015 г.
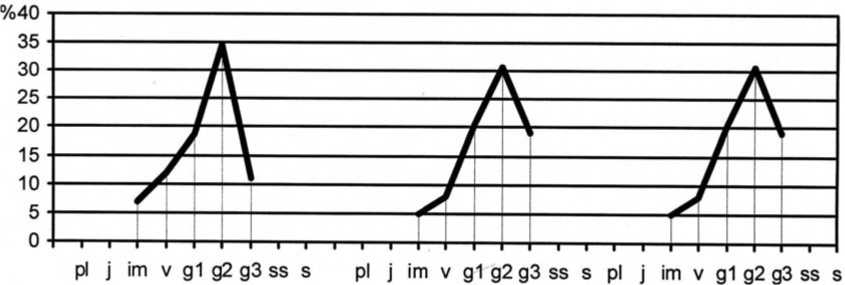
Рис. 5. Динамика возрастной структуры интродукционной популяции Linum nervosum Waldst. et Kit. в ассоциации Festucetum ( valesiacae ) elytrigiosum ( repentis )
Age structure development of Linum nervosum Waldst. et Kit. introduced population in Festucetum ( valesiacae) elytrigiosum (repentis ) association
2010 г.
2014 г.
2015 г.
Рис. 6. Динамика возрастной структуры интродукционной популяции Linum nervosum Waldst. et Kit. в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae )
Age structure development of Linum nervosum Waldst. et Kit. introduced population in Festucetum ( valesiacae ) viciosum ( tenuifoliae )
Исследованные популяции короткокорневищных видов ( Filipedula vulgaris , Fragaria viridis , Galium ruthenicum , Veronica sclerophylla ) характеризуются неполночлен-ностью и преобладанием молодых растений в возрастном составе (рис. 7-8). Такой характер возрастных спектров можно объяснить, с одной стороны, влиянием неблагоприятных факторов, которые тормозят активность генеративного размножения и благоприятствуют активизации вегетативного, с другой стороны – торможением онтогенетического развития. В первую очередь это касается вир-гинильного и молодого генеративного возрастных состояний, в которых особи могут находиться очень продолжительное время. На начальных стадиях онтогенеза это явление проявляется менее четко и имматурные растения довольно быстро переходят в вирги-нильное состояние. Почти все исследованные популяции короткокорневищных видов относятся к молодым нормальным, неполночленным в результате отсутствия старых генеративных, субсенильных и сенильных групп.
Для короткокорневищных видов отмечено преобладание вегетативного размножения при ухудшении условий существования. В свою очередь, активизация вегетативного нием жизненного уровня растений, которое связано с увеличением плотности особей и обострением внутри- и межвидовой конкуренции, что отражается на габитуальных параметрах растений, вызывает уменьшение ассимиляционной поверхности.
В период наблюдения, отмечено, что процесс старения популяций Galium ruthenicum носит однонаправленный характер, но и это еще не меняет характер возрастного типа популяций в целом (рис. 7).
У Veronica sclerophylla осуществляется постоянное семенное возобновление и пополнение популяций молодыми растениями, но часть из них погибает. В связи с этим плотность популяций вида меняется не существенно. Популяция Veronica sclerophylla в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae ) была представлена только молодой частью возрастного спектра и являлась инвазионной. После проведения эксперимента с ранневесенними палами, на втором экспериментальном участке в 2006 г., в возрастном составе популяции Veronica sclerophylla появились молодые генеративные растения, а с 2010 г. отмечено и появление зрелых генеративных особей, в последующие года наблюдалось очень медленное увеличение численности генеративных растений.
размножения часто сопровождается ухудше-
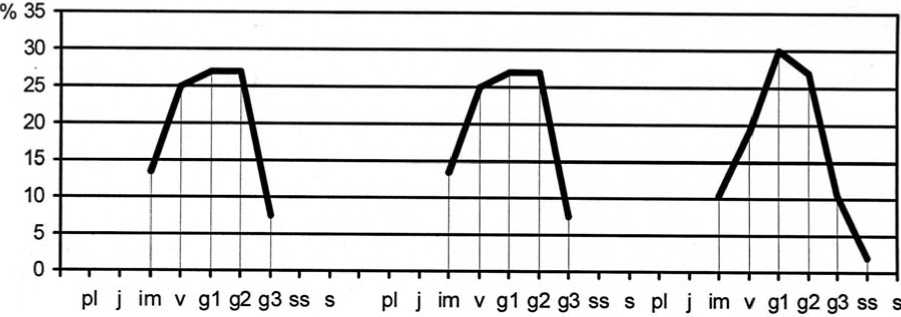
2010 г. 2014 г. 2015 г.
Рис. 7. Динамика возрастной структуры интродукционной популяции Galium ruthenicum Willd. в ассоциации Festucetum ( valesiacae ) elytrigiosum ( repentis )
Age structure development of Galium ruthenicum Willd. introduced population in Festucetum (valesiacae) elytrigiosum (repentis) association
В интродукционных популяциях видов (Filipendula vulgaris, Fragaria viridis), характе- ризующихся интенсивным вегетативным размножением, в результате которого возникает глубоко омоложенное потомство (преобладают растения ранних возрастных состояний, среди которых отмечены и ювенильные особи), проявляются более резкие колебания плотности. Онтогенетическое развитие, особенно на первых этапах, осуществляется быстрее, чем у особей предыдущего вида, но в целом, происходит постепенное старение популяций. Так, если в конце вегетационного периода 2004 г., популяции Filipendula vulgaris занимали промежуточное положение между инвазионными и молодыми нормальными, то в 2010 г. они уже принадлежали к молодым нормальным, неполночленным в
( lessingianae ) и Festucetum ( valesiacae ) viciosum ( tenuifoliae ), благодаря появлению молодых и средневозрастных генеративных растений, доля которых возрастает в течение 2011-2015 гг. (рис. 8). Будущее таких популяций уже более прогнозируемо. Они имеют более высокие шансы стабилизировать свое положение в фитоценозах, увеличить фито-ценотическую роль в условиях мезофитиза-ции растительного покрова за счет увеличения численности генеративных особей, в большей степени ответственных за возобновление, активизации вегетативного размножения, что обеспечит популяции жизнеспособ-
2010 г. .......X'1 2014 г.
■2015 г.
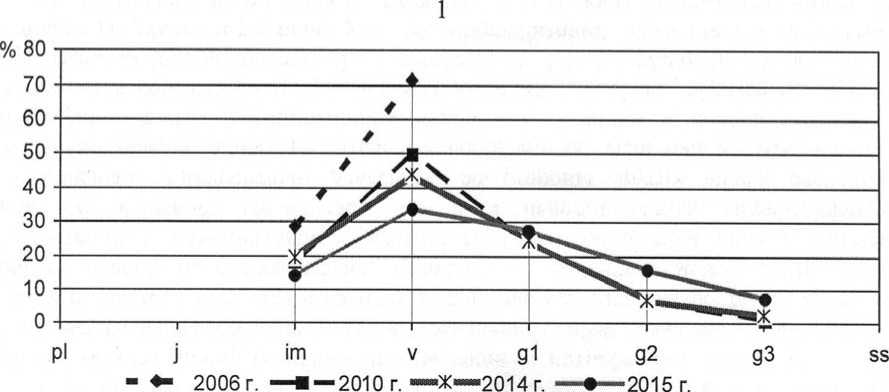
Рис. 8. Динамика возрастной структуры интродукционных популяций короткокорневищних видов в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae ): 1 – Filipendula vulgaris Moench, 2 – Veronica sclerophylla Dubovik
Age structure development of the introduced populations of short-rhizome species in Festucetum ( valesiacae ) viciosum ( tenuifoliae ) association: 1 – Filipendula vulgaris Moench, 2 – Veronica sclero-phylla Dubovik
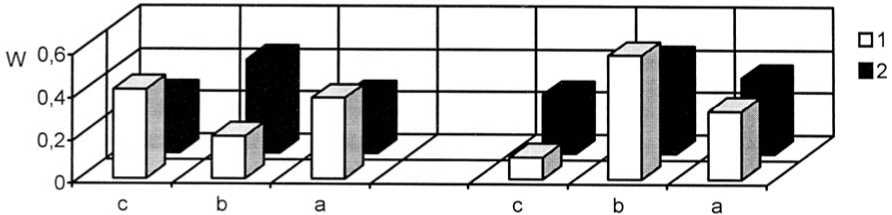
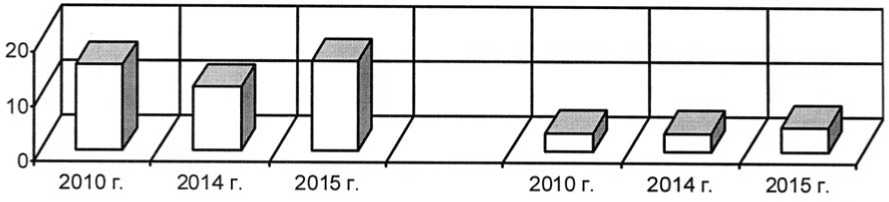
Отмечено повышение плотности и жизненности популяций Filipendula vulgaris и Veronica sclerophylla после проведения палов (рис. 9-10) и сокращения численности популяции Vicia tenuifolia (вид, оказывающий лена из фитоценозов). В результате изменилось положение этих видов в растительных сообществах в лучшую сторону. В целом, отмечено повышение плотности интродукционных популяций всех модельных коротко- сильное конкурентное влияние на степные корневищных видов.
виды растений в меняющихся условиях существования, поэтому часть особей была уда-
2014 г.
2015 г.
Рис. 9. Динамика виталитетной структуры интродукционных популяций короткокорневищных видов в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae ): 1 – Filipendula vulgaris Moench, 2 – Veronica sclerophylla Dubovik; W – встречаемость, "а" – высший класс, "b" – средний класс, "с" – низший класс
Vitality structure development of the introduced populations of short-rhizome species in Festucetum ( valesiacae ) viciosum ( tenuifoliae ) association
1 2
Рис. 10. Динамика плотности интродукционных популяций короткокорневищных видов в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae ): 1 – Filipendula vulgaris Moench, 2 – Veronica sclerophylla Dubovik ; по оси ординат – плотность особей популяций (особь / м2) Density development of the introduced populations of short-rhizome species in Festucetum ( valesia-cae ) viciosum ( tenuifoliae ) association
В исследованных интродукционных популяциях Thalictrum minus и Paeonia tenuifolia (в статье предоставлена информация не только по популяциям видов-эдификаторов, до-минантов, но и видов, в структуре популяций которых осуществляются более заметные и быстрые изменения) в период наблюдения не зафиксировано семенное возобновление (для этих видов характерны семенной и вегетативный способы размножения, но в условиях мезофитизации начинает преобладать второй), что возможно, объясняется отсутствием благоприятных условий для прорастания и приживания подроста, плотной задер-нованостью почвы. Вместе с этим, интенсивное вегетативное размножение обеспечивает пополнение молодыми растениями. Интродукционные популяции неполночленные, в возрастном составе преобладают молодые растения. При пополнении популяции Paeo-nia tenuifolia молодыми особями в результате вегетативного размножения отмечено омо- ложение потомства до имматурного состояния. В зависимости от условий произрастания у Thalictrum minus образуются две формы (короткокорневищная и длиннокорневищная). Короткокорневищные растения вегетативно малоподвижны, самоподдержа-ние популяций осуществляется за счет семенного размножения, вегетативное размножение происходит в результате неспециализированной нормальной партикуляции особей в средневозрастном и старом генеративном состоянии (новая особь чаще всего находится в том же возрастном состоянии, что и материнская, иногда возможно омоложение потомства до молодого генеративного состояния). Короткокорневищная форма образуется в более ксерофитных условиях произрастания. Длиннокорневищные растения являются вегетативно подвижными, вегетативное размножение становится более значимым, возможно омоложение вегетативно возникшего потомства до виргинильного состояния (Остапко, Ибатулина, 2008). Это явление в пределах Донбасса описано для природных степных сообществ С.Н. Зиман (1976). Формирование длиннокорневищной формы характерно для фитоценозов, в которых степной растительный покров подвергается процессам мезофитизации, что и наблюдается в данном случае.
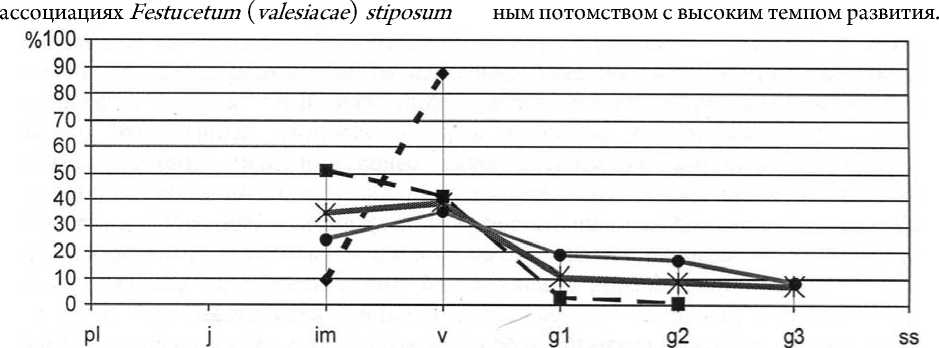
На начальном этапе исследований интродукционные популяции Paeonia tenuifolia в ассоциациях Festucetum ( valesiacae ) stiposum ( lessingianae ) и Festucetum ( valesiacae ) viciosum ( tenuifoliae ) были инвазионными, не устойчивыми (в результате гибели вегетативных растений популяция может исчезнуть из состава растительного сообщества). На участках осуществлялось ежегодное сенокошение и уборка пожнивных остатков. После проведения эксперимента с палами на втором экспериментальном участке в популяции этого вида в фитоценозе Festucetum ( valesiacae ) viciosum ( tenuifoliae ) увеличилась доля молодых растений, в основном вегетативного происхождения. Возрастной состав популяции стал разнообразнее в результате ускоренного прохождения особями онтогенетического развития (рис. 11), также отмечено и улучшение жизненного состояния растений (рис. 12).
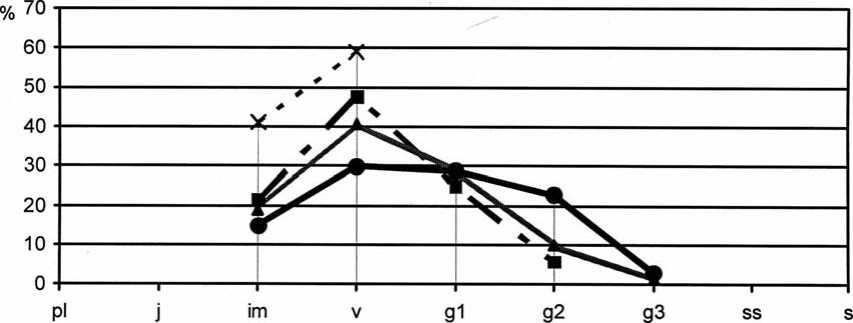
- X- 2006г. -* 2010г. .......*" 2014г. • 2015г.
Рис. 11. Динамика возрастной структуры интродукционной популяции Paeonia tenuifolia L. в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae )
Density structure development of the introduced populations of Paeonia tenuifolia L. association in Festucetum ( valesiacae ) viciosum ( tenuifoliae ) association
c b a
02010 г. 02014 г. 0 2015 г
Рис. 12. Динамика виталитетной структуры интродукционной популяции Paeonia tenuifolia L. в ассоциации Festucetum ( valesiacae ) viciosum ( tenuifoliae ); W – встречаемость, "а" – высший класс, "b" – средний класс, "с" – низший класс
Vitality structure development of Paeonia tenuifolia L. introduced populations in Festucetum ( vale-siacae ) viciosum ( tenuifoliae ) association
Популяция Paeonia tenuifolia заняла более прочное положение в растительных сообществах, в чем не последнюю роль сыграло ослабление давления со стороны обильного ме-зофитного разнотравья и хорошо развитых популяций длиннокорневищных злаков, которые в большей степени характерны для лугово-степных и луговых фитоценозов. Подобная картина наблюдалась и для популяций Thalictrum minus .
Менее лабильной является пространственная структура популяций всех видов. Как показали наблюдения, исследованные попу- ляции степных видов растений в течение многих лет сохраняют контагиозное размещение особей в пространстве. Контагиозный тип размещения растений является общей чертой не только для интродукционных популяций эдификатров степных фитоценозов, но и для интродукционных популяций видов-спутников в искусственных степных растительных сообществах на I и II экспериментальных участках. Также не отмечено и заметных изменений плотности этих популяций по годам (рис. 13-14).
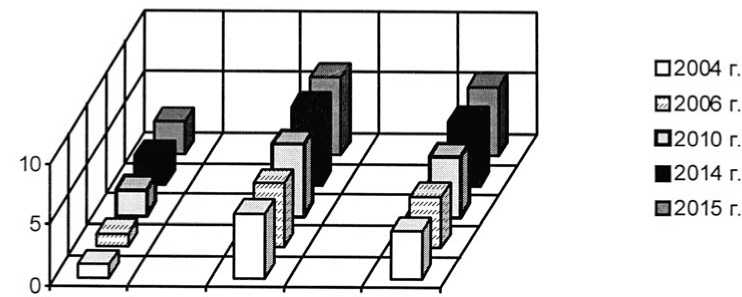
Рис. 13. Динамика плотности интродукционных популяций Thalictrum minus L. в искусственных степных фитоценозах: ассоциации 1 – Festucetum ( valesiacae ) stiposum ( lessingianae ), 2 – Festucetum ( valesiacae ) bromopsiosum ( ripariae ), 3 – Festucetum ( valesiacae ) elytrigiosum ( repentis ); по оси ординат – плотность особей ценопопуляции (особь /м2)
Density development of the introduced populations of Thalictrum minus L. in the artificial steppe phytocenoses
1 2 3
Рис. 14. Динамика плотности интродукционных популяций Paeonia tenuifolia L. в искусственных степных фитоценозах: ассоциации 1 – Festucetum ( valesiacae ) stiposum ( lessingianae ), 2 – Festucetum ( valesiacae ) elytrigiosum ( repentis ), 3 – Festucetum ( valesiacae ) viciosum ( tenuifoliae ); по оси ординат – плотность особей ценопопуляции (особь /м2)
Density development of the introduced populations of Paeonia tenuifolia L. in the artificial steppe phytocenoses
Результаты
Таким образом, интродукционные популяции большинства модельных степных видов занимают прочное положение в сложившихся искусственных сообществах, являются саморегулирующимися системами и не зависят от заноса диаспор. Наиболее устойчивыми являются популяции видов-эдификаторов. Изменения эколого-демографической структуры популяций степных видов растений проявляются в упрочении их положения в составе растительных сообществ, усилении фитоценотической роли, вследствие чего фитоценозы остаются сегодня устойчивыми системами. Но, мезофитизация растительного покрова (очень медленная) искусственных фитоценозов в будущем может привести к существенному ухудшению состояния популяций степных видов. Ежегодное выкашивание не является сильным сдерживающим фактором, а только замедляет скорость преобразования фитоценозов в сообщества другого типа. Положительное влияние пирогенного фактора на развитие популяций степных видов в искусственных фитоценозах явилось основанием для более детального изучения его воздействия, что позволяет рекомендовать выжигание для дальнейших исследований при разработке комплекса мероприятий эффективного регулирования развития степных экосистем (третий экспериментальный участок, режим ежегодного проведения ранневесенних [конец марта – начало апреля] палов). В этом случае существенную роль играет совпадение сроков выжигания с периодом стабильного выпадения осадков. Наиболее подходящим периодом является ранневесенний.
После проведения ранневесенних палов на протяжении 12-ти лет отмечено, что в фитоценозе не изменяется ни флористический, ни биоморфологический состав, сохраняется его организация. Степные виды растений, в том числе злаки и осоки, нормально функционировали: цвели и плодоносили, они не получили от огня существенных повреждений, поскольку только начинали вегетировать и повреждения были поверхностные. Выгорание было неравномерное: подстилка, в местах, где она плотно прилегала к почве, местами была лишь поверхностно задета огнём, сила которого была не велика, он был беглым и поверхностным, что не вызывало выгорания особей. Изменилось соотношение видов: увеличилось проективное покрытие плотно-дерновинных злаков за счёт разрастания их дерновин и увеличения плотности популяций, в результате угнетающего воздействия пирогенного фактора на развитие популяций длиннокорневищных вегетативно подвижных видов: Vicia tenuifolia, Elytrigia repens, Bromopsis inermis. Растительное сообщество очень быстро восстановилось, хотя это скорее отрастание надземных частей побегов, чем восстановление, предполагающее наличие каких-либо сукцессионных стадий. Так как палы способствовали ослаблению влияния длиннокорневищных видов, то злаки (Stipa capillata, S. lessingiana, S. grafiana, Fes-tuca rupicola, F. valesiaca, Phleum phleoides) начинают занимать освобожденное место и особи ускоряют темпы прохождения онтогенетических фаз развития. При этом возрастает не только общее количество особей на единицу поверхности, но и увеличивается площадь, занимаемая отдельными экземплярами (усиливается мощность их развития). В основном, это касается Stipa capillata, Festuca rupicola, F. valesiaca. В итоге возможна смена доминантов в будущем и возникновение нового фитоценоза. Интродукционные популяции Festuca rupicola, F. valesiaca, видов рода Stipa в искусственном растительном сообществе красочной разнотравно-злаковой степи на третьем экспериментальном участке, относятся к устойчивым, они занимают довольно прочное положение в фитоценозе. Популяции являются неполночленными нормальными, зрелыми. Максимум в возрастном спектре приходится на группу зрелых генеративных растений (рис. 15-20). Эти изменения типа возрастного спектра интродукционных популяций проходили постепенно с момента начала проведения эксперимента по воздей- ствию на растительный степной покров пирогенного фактора. Ускорился процесс развития популяций Stipa capillata, S. lessingiana, S. grafiana, Festuca rupicola, F. valesiaca, Phleum phleoides по сравнению с популяциями на участках с абсолютно заповедным режимом и с ежегодным укосом осенью, на которых популяции эдификаторов остаются неполночленными (отсутствуют субсенильные и сенильные растения) нормальными, молодыми. Увеличились плотность интродукционных популяций (рис. 21), возросла доля участия генеративных растений в их возрастном составе, а также мощность особей, что в существенной степени связано с повышением жизненного состояния растений (рис. 22). Для поддержания жизнеспособности популяций плотнодерновинных степных эдификаторов ключевыми являются группы генеративных растений, особенно средневозрастных как наиболее развитых. Наличие большого количества генеративных растений даёт возможность популяциям быстро увеличивать плотность, что способствует укреплению позиций вида в растительном сообществе. Посредством семенного потомства обеспечивается поливариантность развития популяций, виталитетных спектров, то есть в целом повышается адаптивный потенциал. Не только разнообразие возрастного, но и виталитеного состава повышает устойчивость популяций, так как разнокачественные особи наиболее полно используют жизненное пространство.
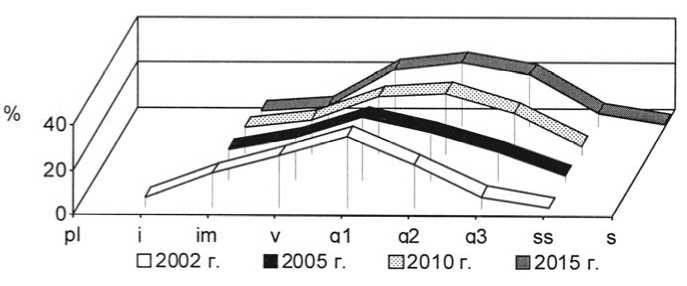
Рис. 15. Динамика возрастной структуры интродукционной популяции Festuca valesiaca Gaudin
на III экспериментальном участке при воздействии пирогенного фактора
Age structure development of Festuca valesiaca Gaudin in III trial plot under pyrogenic effect
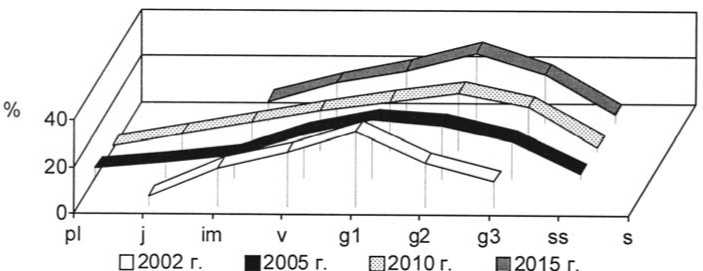
Рис. 16. Динамика возрастной структуры интродукционной популяции Festuca rupicola Heuff . на III экспериментальном участке при воздействии пирогенного фактора
Age structure development of Festuca rupicola Heuff in III trial plot under pyrogenic effect
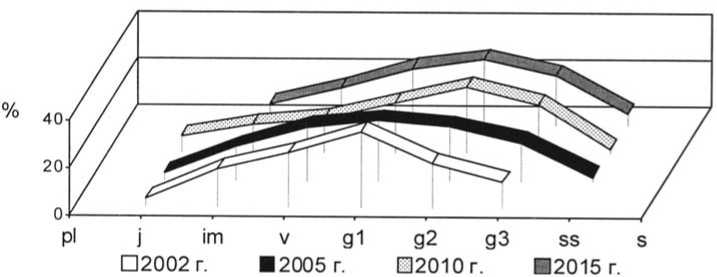
Рис. 17. Динамика возрастной структуры интродукционной популяции Stipa lessingiana Trin. et Rupr. на III экспериментальном участке при воздействии пирогенного фактора
Age structure development of Stipa lessingiana Trin. et Rupr. in III trial plot under pyrogenic effect
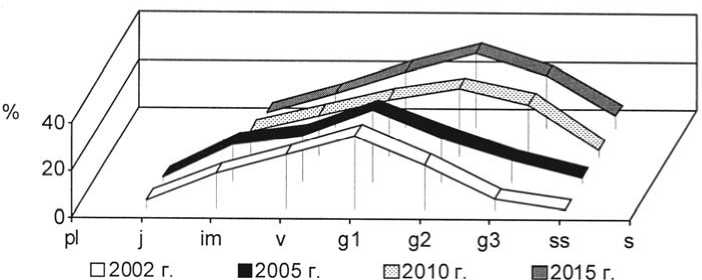
Рис. 18. Динамика возрастной структуры интродукционной популяции Stipa capillata L. на III экспериментальном участке при воздействии пирогенного фактора
Age structure development of Stipa capillata L. in III trial plot under pyrogenic effect
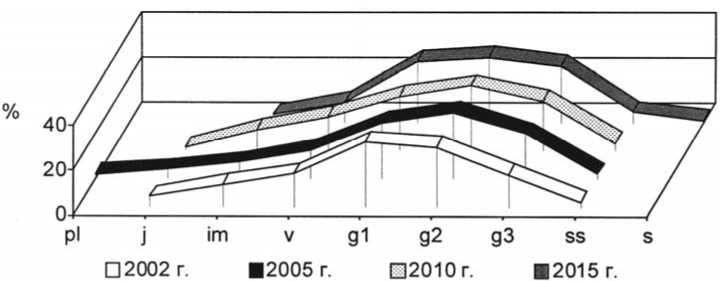
Рис. 19. Динамика возрастной структуры интродукционной популяции Stipa grafiana Steven на III экспериментальном участке при воздействии пирогенного фактора
Age structure development of Stipagrafiana Steven in III trial plot under pyrogenic effect
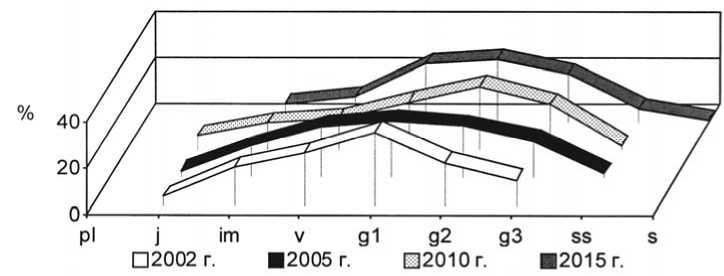
Рис. 20. Динамика возрастной структуры интродукционной популяции Phleum phleoides (L.) H. Karst. на III экспериментальном участке при воздействии пирогенного фактора
Age structure development of Phleumphleoides (L.) H. Karst. in III trial plot under pyrogenic effect
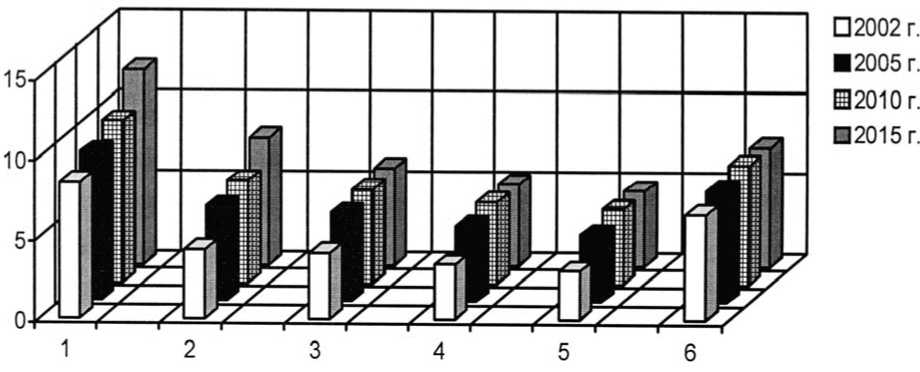
Рис. 21. Динамика плотности интродукционных популяций эдификаторов степных фитоценозов на III экспериментальном участке при воздействии пирогенного фактора: 1 – Festuca vale-siaca Gaudin , 2 – F. rupicola Heuff ., 3 – Stipa lessingiana Trin. et Rupr., 4 – S. capillata L., 5 – S. grafi-ana Steven, 6 – Phleumphleoides (L.) H. Karst.
Density development of the introduced populations of steppe phytocenoses edificator species in III trial plot under pyrogenic effect
Список литературы Мониторинг интродукционных популяций степных видов растений в условиях экспериментальной степи
- Берко Й.М. До питання вивчення i перiодизацiї великого життєвого циклу вегетативно-рухливих рослин. Укр. ботан. журн., 1976, т. 33, № 6, с. 605-609. Борисова И.В., Попова Т.А. Возрастные этапы формирования дерновины степных злаков. Ботан. журн., 1971, т. 56, № 5, с. 619-626.
- Боровик Л.П. Влияние пирогенного фактора на популяции дерновинных злаков. Збереження бiорiзноманiтностi на пiвденному сходi України. Донецьк: ТОВ "Лебiдь", 2004, с. 17-18.
- Боровик Л.П., Боровик Е.Н. Проблема режима сохранения степи в заповедниках: пример Стрельцовской степи. Степной бюллетень, 2006, № 20, с. 29-33.
- Воронцова Л.И., Заугольнова Л.Б. Мультивариантность развития особей в течение онтогенеза и её значение в регуляции численности и состава ценопопуляций растений. Журн. общ. биологии, 1978, № 4, с. 555- 561.
- Гавриленко В.С. Абсолютно заповедный режим и пожары в степных заповедниках: непривычное решение. Степной бюллетень. 2007а, № 23-24. с. 25-26.
