Мониторинг эктомикоризных грибов ельника кисличного в южнотаежных лесах Пермского края

Автор: Шишигин Александр Сергеевич, Переведенцева Лидия Григорьевна, Боталов Виталий Сергеевич
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2019 года.
Бесплатный доступ
В Пермском крае с 1975 г. по настоящее время проводится мониторинг агарикоидных базидиомицетов в ельнике кисличном подзоны южной тайги. Исследования ведутся стационарным методом на пробной площади, размером 50 × 20 м, в три периода: I-й - в 1975-1977 гг., II-й - в 1994-1996 гг., III-й - в 2010-2012 гг. К настоящему времени в ельнике кисличном обнаружено 214 видов и внутривидовых таксонов агарикоидных базидиомицетов, 80 из которых (37.4 %) являются микоризообразователями, входящими в состав 18 родов и 7 семейств. Преобладают виды сем. Cortinariaceae, Russulaceae и Boletaceae, что характерно для бореальной зоны. Наибольшее число видов содержится в родах Cortinarius (21 вид), Russula (15), Lactarius (10), Amanita (7), что свойственно лесным ценозам подзоны южной тайги. Отмечено, что видовой состав эктомикоризных грибов с течением времени меняется в большей степени (коэффициенты Жаккара (J×100): J = 32-46), чем видовой состав сосудистых растений. Эктомикоризные грибы доминируют по биомассе и числу базидиом. Изучена связь между разнообразием и продуктивностью эктомикоризных грибов с основными климатическими показателями (май-сентябрь). Установлено, что повышение средней месячной температуры воздуха в июне благоприятствует «плодоношению» эктомикоризных грибов в августе (rs = 0.67; p
Эктомикоризные агарикоидные базидиомицеты, мониторинг, ельник кисличный, пермский край
Короткий адрес: https://sciup.org/147227090
IDR: 147227090 | УДК: 582.287.238 | DOI: 10.17072/1994-9952-2019-3-280-290
Monitoring of ectomycorrhizal fungi in the sorrel spruce forest in the southern taiga of the Perm region
The monitoring of agarics in the Perm Region has been done since 1975 till the present time in the sorrel spruce forest (the southern taiga subzone). Research is conducted by a stationary method (the test area is 50 ×20 m) in three stages: I period - 1975-1977; II period - 1994-1996; III period - 2010-2012. By the present time, 214 species and intraspecific taxa of agarics have been found in the sorrel spruce forest, 80 of them (37.4 %) are ectomycorrhizal fungi. The fungi belong to 7 families and 18 genera. Families Cortinariaceae, Russulaceae and Boletaceae prevail, that is typical for a boreal area. The majority of species are in genera: Cortinarius (21 species), Russula (15), Lactarius (10), Amanita (7), that is typical for the southern taiga subzone. It is noted that the species composition of ectomycorrhizal fungi (Jaccard index×100: J = 32-46) changes over time more than the species composition of higher vascular plants. Ectomycorrhizal fungi are dominant by biomass and the number of basidioms. The relationship between diversity and productivity of ectomycorrhizal fungi with the main climatic indicators (May-September) was studied. It was found that the increase in the average monthly air temperature in June favors the "fruiting" of ectomycorrhizal fungi in August (rs = 0.67; p
Текст научной статьи Мониторинг эктомикоризных грибов ельника кисличного в южнотаежных лесах Пермского края
Эктомикоризные грибы, вступающие с высшими растениями в консортивные отношения в форме микосимбиотрофизма, распространены в лесных экосистемах и составляют значительную часть от общего числа обитающих в них агарикоидных базидиомицетов [Переведенцева, 1999; Straatsma, Ayer, Egli, 2001; Фомина, 2001; Straatsma, Krisai-Greilhuber, 2003; Воронина, 2004; Шубин, 2009; Боталов, Переведенцева, 2018].
«Плодоношение» эктомикоризных грибов явля-
ется показателем дополнительного получения углеводов и повышения интенсивности фотосинтеза у древесных растений [Шубин, 2009]. Все микоризообразующие грибы оказывают существенное влияние на минеральное питание, водный баланс древесных растений и играют огромную роль в биохимических циклах важнейших химических элементов в лесных биогеоценозах [Смит, Рид, 2012; Иванов, 2014].
Большой интерес представляют исследования эктомикоризных грибов в климаксных сообществах, которые имеют довольно стабильный видовой состав высших растений, в особенности древесных пород. В подзоне южной тайги Пермского края климаксными сообществами являются еловые леса.
В связи с этим, целью нашей работы является мониторинг эктомикоризных грибов ельника кисличного. Для достижения цели были поставлены следующие задачи: 1) выявление видового состава эктомикоризных агарикоидных базидиомицетов ельника кисличного; 2) проведение анализа таксономической структуры микоризообразователей; 3) определение доминирующих видов грибов по числу базидиом и их биомассе; 4) выявление зависимости «плодоношения» эктомикоризных грибов от количества осадков и температуры воздуха.
Материал и методы исследования
Материал
В Пермском крае (Добрянский р-н, окрестности ООПТ «Верхняя Кважва») в лесных ценозах с 1975 г. проводится мониторинг агарикоидных ба-зидиомицетов стационарным методом, позволяющим количественно оценить и выявить структуру грибного компонента, что необходимо для понимания развития природных экосистем. Пробная площадь размером 50×20 м была заложена в ельнике кисличном Л.Г. Переведенцевой [1999]. Геоботаническое описание было выполнено согласно В.Н. Сукачёву и Е.Н. Зонну [1961]. Латинские названия сосудистых растений приводятся по «Иллюстрированному определителю растений Пермского края» [2007].
Ельник кисличный расположен на равнинной местности, коренное сообщество, возраст которого около 135 лет. Состав древостоя 5Е2П2Лп1Б. Сомкнутость крон 0.6. Подрост состоит из Picea obovata Ledeb., Abies sibirica Ledeb., Tilia cordata Mill. и Betula pendula Roth. Кустарниковый ярус образован такими видами, как Sorbus aucuparia L., Lonicera xylosteum L., Padus avium Mill. Проективное покрытие кустарничково-травяного яруса составляет 70–80%, где доминирует Oxalis acetosella L., Dryopteris carthusiana Vill., Gymnocar-pium dryopteris L. Зелёные мхи растут, в основном, около стволов деревьев. На стволах древесных растений, кроме зеленых мхов, встречаются и лишайники. В лесу много валежника. Видовой состав древесного яруса в течение 1975–2012 гг. оставался неизменным.
За время исследований (1975–1977 гг., 1994– 1996 гг. и 2010–2012 гг.), по данным метеостанции г. Добрянки, погодные условия отличались от средних многолетних характеристик повышенным фоном температур воздуха. Количество осадков варьировало и зачастую отличалось от средних многолетних данных. Самыми неблагоприятными для развития грибов следует считать 1975 и 2011 гг. как наиболее засушливые, и 1994 г., – отличающийся большим количеством осадков и пониженными температурами воздуха в июне– августе.
Методы исследования
Учет видового разнообразия, количества и биомассы плодовых тел грибов осуществлялся один раз в декаду, в августе. В сентябре изучался лишь видовой состав грибов. Первая серия наблюдений была проведена в 1975–1977 гг., вторая – в 1994–1996 гг., третья – в 2010–2012 гг.
Степень сходства или различия по видовому составу грибов вычислялась по коэффициенту Жаккара [Грейг-Смит, 1967; Леонтьев, 2008]:
J = —c—, a + b - c где J – индекс общности; c – число общих видов в двух сравниваемых биоценозах; a, b – число видов грибов в каждом из ценозов.
Доминирующие виды грибов устанавливались по числу базидиом (шт/га) и по их воздушно-сухой биомассе (кг/га). Для выявления доминирующих видов грибов по биомассе и числу базидиом использовался индекс доминирования [Bochus, Babos, 1960]:
Д = а / b ×100, где Д – индекс доминирования; а – число базидиом (или биомасса) грибов данного вида; b – число ба-зидиом (или биомасса), собранных на всей учетной площади.
К доминантам были отнесены лишь те виды грибов, которые имели индекс доминирования, равный 5 или более, что составляет 5% или более от общего числа плодовых тел или их биомассы.
Состав эколого-трофических групп грибов в изучаемом ценозе определялся по шкале, предложенной А.Е. Коваленко, с дополнениями некоторых авторов [Коваленко, 1980; Столярская, Коваленко, 1996; Морозова, 2001].
Корреляционный анализ проводился с использованием коэффициента ранговой корреляции Спирмена (rs), так как этот коэффициент не требует проверки распределения на нормальность. Зна- чение коэффициента корреляции rs от 0.3 до 0.7 при p < 0.05 означает среднюю положительную и статистически значимую корреляцию между признаками. При значении rs ≥ 0.70 (p < 0.05) отмечается сильная положительная корреляция; отрицательное значение rs соответствует обратной корреляции [Трухачёва, 2013].
Математическая обработка осуществлялась при помощи программ Microsoft Office Excel 2016 и StatSoft Statistica 10. При корреляционном анализе нами рассматривалось влияние некоторых климатических показателей на биоту эктомикоризных грибов. В качестве характеристик микобиоты были взяты: число и биомасса базидиом эктомикориз-ных грибов за август; число видов за август– сентябрь. В качестве климатических показателей использованы данные метеостанции г. Добрянки: средняя месячная температура воздуха ( ° С) за май–сентябрь; сумма осадков (мм) по месяцам с мая по сентябрь; сумма осадков по декадам августа.
Результаты и их обсуждение
Биота эктомикоризных грибов ельника кисличного довольно разнообразна. Далее приведен список видов, составленный по системе, принятой М. Moser [1983], с некоторыми дополнениями [Пере-веденцева, 1999]. Для каждого вида приводятся латинское и русское названия. В скобках указаны синонимы грибов, соответствующие современной классификации [MycoBank Databases]. Римскими цифрами обозначены месяцы вегетационного периода. Хозяйственная значимость: съед. – съедобный, несъед. – несъедобный, яд. – ядовитый. Арабскими цифрами обозначен период выявления вида: 1 – в 1975–1977, 2 – в 1994–1996, 3 – в 2010–2012 гг.
Аннотированный список видов грибов
AGARICALES Clements – Агарикальные
Amanitaceae R. Heim ex Pousar – мухоморовые: Amanita battarrae (Boud.) Bon – поплавок умбро-во-желтый, VII–IX, съед. (2); A. crocea (Quél.) Singer – п. шафранный, под лиственными деревьями, VIII, съед. (1, 3); A. fulva Fr. – п. желтокоричневый, VII–VIII, съед. (1–3); A. muscaria (L.) Lam. – мухомор красный, VII–VIII, яд. (1, 3); A. porphyria Alb. & Schwein. – м. порфировый, VIII–IX, яд. (1, 3); A. rubescens var. rubescens Pers. – м. краснеющий, VIII–IX, съед. (3); A. vaginata (Bull.) Lam. – п. серый, VIII–IX, съед. (1, 2).
Cortinariaceae R. Heim ex Pouzar – паутинни-ковые: Cortinarius acutus (Pers.) Fr. – паутинник заостренный, VIII, несъед. (2, 3); C. albidus Peck – п. беловатый, VIII, несъед. (3); C. alboviolaceus (Pers.) Fr. – п. бело-фиолетовый, VII–VIII, съед.
(3); C. anomalus (Fr.) Fr. – п. необычный, VIII, не-съед. (1); C. argentatus (Pers.) Fr. – п. серебристый, VIII–IX, несъед. (3); C. armeniacus (Schaeff.) Fr. – п. абрикосово-оранжевый, VII–VIII, несъед. (1, 3); C. betuletorum M.M. Moser ex M.M. Moser – п. березовый, VIII, несъед. (1, 3); C. brunneus (Pers.) Fr. – п. темно-бурый, VIII–IX, несъед. (3); C. decipiens Fr. – п. обманчивый, VIII, несъед. (3); C. decoloratus (Fr.) Fr. – п. обесцвеченный, VIII, несъед. (3); C. gentilis (Fr.) Fr. – п. благородный, VI–VIII, несъед. (3); C. hemitrichus (Pers.) Fr. – п. полуопушенный, VII–VIII, несъед. (1–3); C. impennis Fr. – п. неопушенный, VIII, несъед. (1); C. jubarinus Fr. – п. сияющий, VIII, несъед. (3); C. leucopus (Bull.) Fr. – п. белоножковый, VIII, не-съед. (3); C. megasporus Singer – п. крупноспоровый, VIII, несъед. (2, 3); C. privignoides Rob. Henry – п. клубненогий, VIII, несъед. (3); C. rigidus (Scop.) Fr. – п. умбровый, VIII, несъед. C. tubulipes J. Favre – п. полоножковый, VIII–IX, несъед. (3); C. uraceus Fr. – п. обожженный, VIII, несъед. (2, 3); C. violaceus (L.) Gray – п. фиолетовый, VII– VIII, несъед. (1, 3); Hebeloma crustuliniforme (Bull.) Quél. – гебелома клейкая, ложный валуй, VII–IX, несъед. (2, 3); H. hiemale Bres. – г. зимняя, VIII, несъед. (3); H. pusillum J. E. Lange – г. малюсенькая, VIII, несъед. (2); H. sinapizans (Paulet) Gillet – г. редечная, VII–IX, несъед. (1); Inocybe flocculosa Sacc. – волоконница клочковатая, VIII– IX, несъед. (1-3); I . geophylla (Sowerby) P. Kumm. – в. земляная, VIII–IX, яд. (1, 3); I. hirtella Bres. – в. короткожестковолосая, VIII, несъед. (3); I. lanuginosa (Bull.) P. Kumm. – в. шерстистая, VIII, яд. (1–3); I. rimosa (Bull.) P. Kumm. – в. трещиноватая, VII–IX, яд. (1–3).
Entolomataceae Kotlaba & Pouzar – энтоломо-вые, розовопластинниковые: Clitopilus prunulus ( Scop.) P. Kumm. – клитопил сливовый, VIII, съед. (1, 2); Entoloma rhodopolium (Fr.) P. Kumm. – эн-толома дымчатая, VIII–IX, яд. (1, 2).
Tricholomataceae R. Heim ex Pousar– трихоломо-вые, рядовковые: Clitocybe odora (Bull.) P. Kumm. – говорушка душистая, VII–VIII, несъед. (1–3); Collybia butyracea var. asema (Fr.) Cetto – коллибия масляная, серая, VII–IX, съед. (1–3); C. butyracea var. butyracea (Bull.) Fr. (= Rhodocollybia butyracea (Bull.) Lennox) – коллибия масляная, VII–IX, съед. (1–3); Laccaria lac-cata (Scop.) Cooke – лаковица лаковая, VI–IX, съед. (1–3); Tricholoma flavobrunneum (Fr.) P. Kumm. (= T. fulvum (Fr.) Bigeard & H. Guill.) – рядовка. желто-бурая, VIII–IX, несъед. (3); T. lascivum (Fr.) Gillet – р. приятная, VIII, несъед. (1, 3).
BOLETALES J.-E. Gilbert – Болетальные
Boletaceae Chevall. – трубчатые, болетовые: Boletus edulis Bull. – белый гриб VII–IX, съед. (1, 3); Chalciporus piperatus (Bull.) Bataille – перечный гриб, VIII–IX, несъед. (1, 3); Leccinum auran- tiacum (Bull.) Gray – подосиновик, красноголовик, VII–VIII, съед. (1); L. scabrum (Bull.) Gray – подберезовик обыкновенный, VI–IX, съед. (1, 3); L. versipelle (Fr. & Hök) Snell – подосиновик желто-бурый, VIII–IX, съед. (1, 3); Tylopilus felleus (Bull.) P. Karst. – желчный гриб, VIII, несъед. (1, 3); Xerocomus chrysenteron (Bull.) Quél. (= Xerocomellus chrysenteron (Bull.) Šutara) – бо-лет, моховик трещиноватый, VII–IX, съед. (3); X. rubellus (Krombh.) Quél. – моховик краснеющий, VIII–IX, съед. (1–3); X. subtomentosus (L.) Fr. – м. зеленый, VI – IX, съед. (1, 3).
Paxillaceae Lotsy – паксилловые, свинушковые: Paxillus involutus (Batsch) Fr. – свинушка тонкая, VIII–IX, яд. (1, 2).
RUSSULALES Kreisel ex Kirk et al. – Руссулальные
Russulaceae Lotsy – сыроежковые: Lactarius camphoratus (Bull.) Fr. – млечник камфарный, VIII–IX, несъед. (3); L. deterrimus Gröger – рыжик еловый, VIII, съед. (1); L. flexuosus (Pers.) Gray (= L. zonarius (Bull.) Fr.) – млечник извилистый, серушка, VII–VIII, съед. (1, 3); L. glyciosmus (Fr.) Fr. – м. пахучий, VII–IX, несъед. (2); L. necator (Bull.) Pers. – груздь черный, VIII–IX, съед. (1, 3); L. rufus (Scop.) Fr. – горькушка, VII–IX, съед. (1); L. theiogalus (Bull.) Gray – м. серно-млечный, золотистый, VIII–IX, съед. (1–3); L. torminosus (Schaeff.) Pers. – волнушка, VIII–IX, съед. (1); L. trivialis (Fr.) Fr. – м. обыкновенный, гладыш,
VII–VIII, съед. (1, 3); L. vietus (Fr.) Fr. – м. блеклый, VIII–IX, съед. (1–3); Russula aeruginea Lindbl. ex Fr. – сыроежка зеленая, VII–VIII, съед. (1); R. betularum Hora – с. березовая, VIII–IX, не-съед. (1–3); R. chamaeleontina (Lasch) Fr. (= R. risigallina (Batsch) Sacc.) – с. переменчивая, VII–VIII, съед. (1–3); R. claroflava Grove – с. светло-желтая, VII–VIII, съед. (1–3); R. decolorans (Fr.) Fr. – с. сереющая, VII–VIII, съед. (1, 2); R. delica Fr. – подгруздок белый, VII–VIII, съед. (1); R. emeticicolor (Jul. Schäff.) Singer – с. красная, матовая, VII–VIII, съед. (1, 2); R. foetens Pers. – валуй, VII–VIII, съед. (3); R. fragilis Fr. – с. хрупкая, VIII–IX, несъед. (1–3); R. mustelina Fr. – с. гладкокожая, VIII, съед. (1); R. nauseosa (Pers.) Fr. – с. отвратительная, VIII, съед. (1, 2); R. ochroleuca Fr. – с. бледно-желтая, VII–VIII, съед. (1, 2); R. puellaris Fr. – с. девичья, VII–VIII, съед. (1–3); R. vesca Fr. – с. пищевая, VII–VIII, съед. (1); R. xerampelina (Schaeff.) Fr. – с. селедочная, VII–VIII, съед. (1, 3).
Анализ таксономической структуры
В ельнике кисличном за все время исследований (1975–1977 гг., 1994–1996 гг., 2010–2012 гг.) выявлено 214 видов и внутривидовых таксонов агарикоидных базидиомицетов, 80 из которых являются микоризообразователями, входящими в состав 18 родов и 7 семейств (таблица).
Таксономический состав эктомикоризных агарикоидных базидиомицетов ельника кисличного
|
Порядок |
Семейство (число ро-дов/видов) |
Роды (число видов) |
|
Agaricales |
Amanitaceae (1/7) |
Amanita (7) |
|
Cortinariaceae (3/30) |
Cortinarius (21), Hebeloma (4), Inocybe (5) |
|
|
Entolomataceae (2/2) |
Clitopilus (1), Entoloma (1) |
|
|
Tricholomataceae (4/6) |
Clitocybe (1), Collybia (2), Laccaria (1), Tricholoma (2) |
|
|
Boletales |
Boletaceae (5/9) |
Boletus (1), Chalciporus (1), Leccinum (3), Tylopilus (1), Xerocomus (3) |
|
Paxillaceae (1/1) |
Paxillus (1) |
|
|
Russulales |
Russulaceae (2/25) |
Lactarius (10), Russula (15) |
|
Всего |
7 семейств |
18 родов (80 видов) |
В изучаемом биоценозе микоризообразователи являются преобладающей эколого-трофической группой, к которой относится 37.4% всех выявленных видов агарикоидных грибов. Преобладание микоризообразователей в составе микобиоты естественных лесных ценозов характерно для всей лесной зоны Голарктики [Морозова, 2001, Переве-денцева, Шилкова, 2013; Грибные сообщества…, 2014, 2018].
Ведущими по числу видов оказались семейства: Cortinariaceae (30 видов), Russulaceae (25) и Bole-taceae (9), включающие 80% всех выявленных ми-коризообразователей. Наиболее крупными родами по числу видов эктомикоризных грибов являются: Cortinarius (21 вид), Russula (15), Lactarius (10) и Amanita (7), что характерно для лесных ценозов подзоны южной тайги [Морозова, 2001; Грибные сообщества…, 2012, 2014, 2018].
В третий период наблюдений впервые в ельнике кисличном обнаружено 19 видов микоризных грибов, из числа которых 2 вида ( Cortinarius leucopus, C. tubulipes ) оказались новыми для Пермского края .
Наличие или отсутствие базидиом эктомико-ризных грибов может зависеть от различных факторов, поэтому в ходе ежегодных наблюдений вы- является только часть реально существующих в экосистеме видов грибов [Переведенцева, 1999; Новожилов и др., 2016; Боталов, Переведенцева, 2018]. В связи с этим, все обнаруженные виды эк-томикоризных грибов в ельнике кисличном вклю- чались в общий список. С учетом скрытого видо-
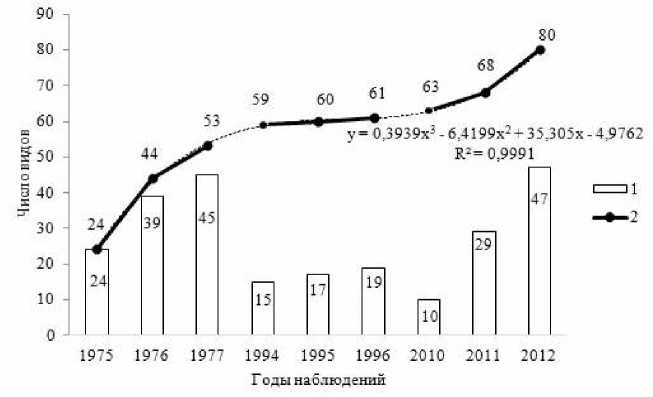
Рис. 1 . Изменение видового разнообразия биоты эктомикоризных грибов по годам наблюдений:
1 – число видов за год наблюдений, 2 – накопление общего выявленного разнообразия (аппроксимация полиномиальной функцией)
Анализируя разнообразие эктомикоризных грибов по годам наблюдений, отметим, что минимальное число видов было выявлено в 2010 г. (10 видов, или 12.5% от общего числа видов микоризных грибов), максимальное – в 2012 г. (47 видов, или 58.8%).
Примечательно, что из 80 видов эктомикориз-
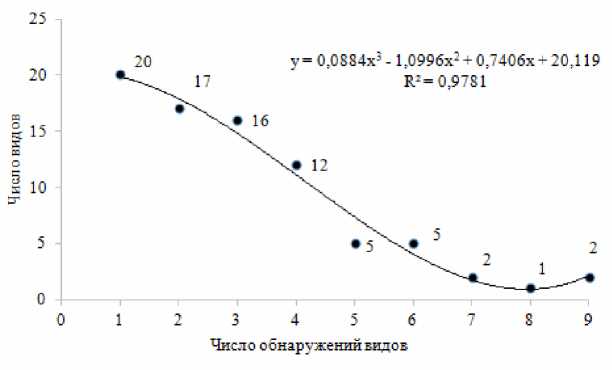
Рис. 2 . Выявление видов эктомикоризных грибов ельника кисличного за все время исследований (аппроксимация полиномиальной функцией)
В ельнике кисличном на протяжении трех периодов исследований происходили существенные изменения в составе ведущих семейств. В первый и второй периоды наблюдений лидирующее положение занимали представители сем. Russulaceae, а к третьему периоду наибольшее число видов относилось к сем. Cortinariaceae (рис. 3).
вого разнообразия отмечено увеличение числа видов микоризообразователей по годам наблюдений, хотя ежегодно выявляемое число видов сократилось во втором периоде наблюдений, а к концу третьего периода вновь увеличилось (рис.1).
ных грибов, выявленных в ельнике кисличном за все три периода, только 2 вида – Laccaria laccata и Lactarius theiogalus – были постоянными и встречались ежегодно, то есть 9 раз . Наибольшее число, 20 видов, обнаруживали только 1 раз в какой-либо один год наблюдений (рис. 2).
Преобладание данных семейств указывает на бореальный характер исследуемой территории [Морозова, 2001; Грибные сообщества…, 2012, 2014, 2018].
Наиболее многочисленными по числу видов в I-й период наблюдений были роды: Russula (14 видов) и Lactarius (8), во II-й – Russula (9) и Corti- narius (5), а III-й период характеризуется преобладанием родов Cortinarius (19) и Russula (7).
При сравнении видового состава эктомикориз-ных грибов по периодам наблюдений было установлено, что число видов микоризных грибов существенно различается. Так, в I-й период было выявлено 53 вида (66.3% от общего числа видов микоризообразователей). Во II-й период число видов сократилось до 32 (40%), а в III-й – увеличилось до 59 видов (73.8%). Вычислив индексы общности по видовому разнообразию эктомикоризных грибов в разные периоды, мы определили, насколько интенсивно произошло изменение биоты эктомикоризных грибов. В отличие от растений-хозяев, видовой состав которых практически остался прежним (за 1975–2012 гг.), биота микоризных грибов подвергалась значительным изменениям по периодам исследования. Индексы общности (по Жаккару: J×100) по грибам между периодами наблюдений колебались от 32 до 46. Появление базидиом эктомикоризных грибов в климаксных сообществах не отличается стабильностью и зависит в большей степени не от растения-хозяина, а от внешних факторов. Но с течением времени общее (скрытое) биоразнообразие увеличивается, а также происходит сближение видового состава эктоми-коризных грибов (J I–II = 39, J II–III = 32, J I–III = 46), что, вероятно, объясняется наличием мицелиаль-
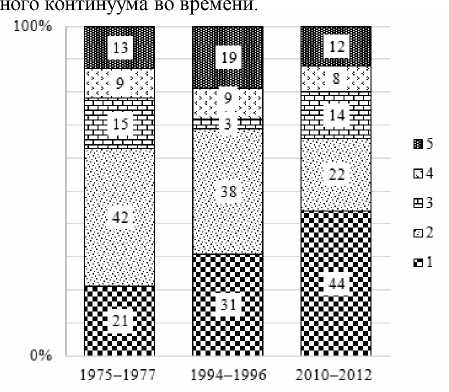
Рис. 3 . Соотношение ведущих семейств экто-микоризных грибов в ельнике кисличном по периодам наблюдений (в процентах, от числа видов микоризообразователей за период):
1 – Cortinariaceae, 2 – Russulaceae, 3 – Boletaceae, 4 – Amanitaceae, 5 – остальные сем-ва
Доминирующие виды эктомикоризных грибов
Доминирующие виды грибов определялись отдельно по числу и по биомассе базидиом. Образование плодовых тел грибов связано с различными факторами: температурой, влажностью воздуха и почвы, наличием субстрата, метеорологическими условиями, наличием достаточного количества питательных веществ в мицелии и т.д. Количество и биомасса базидиом свидетельствуют об активности мицелия и оптимальности условий окружающей среды [Переведенцева, 1999]. В ельнике кисличном за все время исследований выявлено 7 видов эктомикоризных грибов, доминирующих по числу базидиом (43.8 % от общего количества до-минантов по числу базидиом), и 14 видов – по биомассе базидиом (66.6 % от общего числа доми-нантов по биомассе). То есть, микоризообразова-тели в изучаемом ценозе преобладали не только по числу видов, но и имели наибольшие показатели количественной представленности по числу бази-диом и их суммарной биомассе. В разные периоды доминировали: Boletus edulis, Collybia asema, Cortinarius uraceus, Lactarius theiogalus, Leccinum scabrum, Paxillus involutus, Russula claroflava, R. ochroleuca, R. xerampelina и некоторые другие. Число доминантов по биомассе базидиом варьировало по периодам от 4 до 9, по числу базидиом – от 3 до 5. Во все периоды доминировал только один вид Laccaria laccata. Видовой состав доми-нантов к III периоду наблюдений существенно изменился. В большей степени это касается доми-нантов по биомассе. Индексы общности, вычисленные для доминантов разных периодов исследования по числу базидиом, варьировали от 14 до 60 (JI–II = 60, JII–III = 14, JI–III = 20). Индексы общности, установленные для грибов, доминирующих по биомассе, еще ниже и колебались от 0 до 25 (JI– II= 25, JII–III = 0, JI–III = 18). Сравнивая индексы общности по доминантам с индексами, вычисленными по всему биоразнообразию эктомикоризных грибов, отметим, что индексы по доминантам чаще всего ниже, чем по общему разнообразию видов микоризообразователей.
«Плодоношение» эктомикоризных грибов
Большое значение при изучении эктомикориз-ных агарикоидных грибов имеют данные об их урожайности, которые позволяют судить об объеме вторичной продукции биогеоценоза [Грибные сообщества…, 2018]. На «плодоношение» эктомико-ризных грибов оказывают влияние различные факторы. Чаще всего колебания обилия и разнообразия микоризообразователей объясняют погодными условиями вегетационного периода [Переве-денцева, 1999; Шубин, 2009; Иванов, 2016; De la Varga, et al., 2013; Andrew, 2016].
Число базидиом эктомикоризных грибов и их суммарная воздушно-сухая биомасса в разные периоды исследований значительно варьируются. В I-й период наблюдений отмечена наибольшая биомасса грибов (10.1 кг/га). Зафиксировано массовое развитие грибов с крупными плодовыми телами: Boletus edulis, Lactarius theiogalus, Leccinum scabrum, Russula ochroleuca и другие. В остальные периоды происходит постепенное снижение продуктивности эктомикоризных грибов.
По числу базидиом максимальные «урожаи» были во II-й период исследований (30.7 тыс. шт/га), минимальные – в III-й период (3 тыс. шт/га).
По годам наблюдений также отмечаются большие вариации «урожая» грибов, что связано с по- годными условиями не только текущего года, но предыдущих лет. Например, резкое увеличение «урожайности» грибов по биомассе и количеству наблюдалось в 1977 г. после засушливого 1975 г.
(рис. 4). Максимальные показатели «урожайности» микоризообразователей по числу и биомассе бази-диом выявлены в 1977 г. и 1996 г. Самыми «низкоурожайными» были засушливые 1975 г., 2010 г., и 2011 г. В изучаемом биогеоценозе по годам наблюдений выявлено немного статистически значимых корреляций характеристик микобиоты эк-томикоризных грибов с погодными условиями текущего года. Так, с ростом средней месячной тем- пературы воздуха в июне прослеживается увеличение суммарной биомассы базидиом микоризных грибов в августе (rs = 0.67; p < 0.05) (рис. 5).
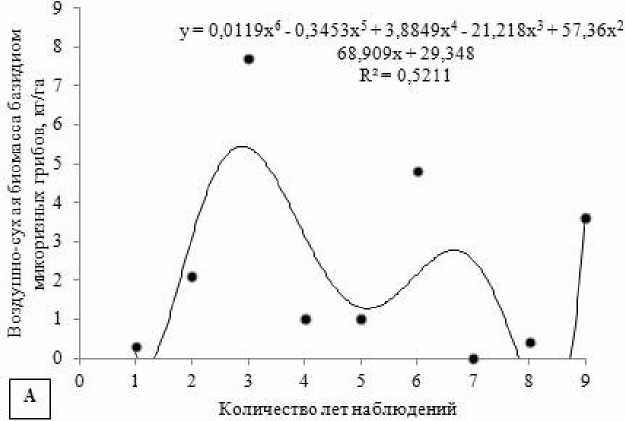
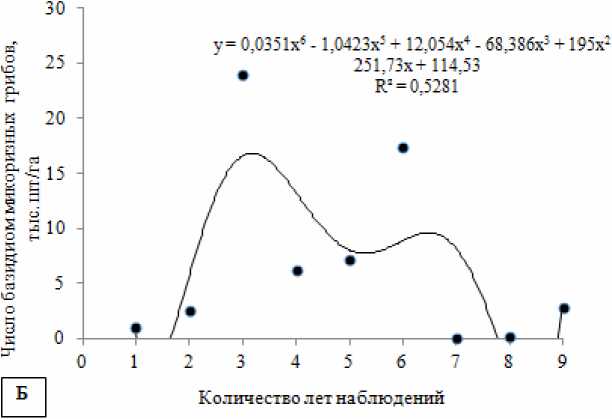
Рис. 4. Динамика «плодоношения» эктомикоризных грибов в течение всего срока наблюдений (аппроксимация полиномиальной функцией):
А – динамика воздушно-сухой биомассы эктомикоризных грибов кг/га в течение всего срока наблюдений; Б – динамика числа базидиом эктомикоризных грибов тыс. шт/га в течение всего срока наблюдений
зидиом эктомикоризных грибов в разные годы на-
Средняя температура воздуха и сумма осадков блюдений.
за август не влияют на количество и биомассу ба-
Отмечено влияние на микобиоту метеорологических показателей по декадам наблюдений. Так, при повышении количества осадков в первую декаду августа выявлено увеличение числа базидиом микоризных грибов ( r s = 0.59; p < 0.05). Напротив, обильные осадки в течение трех декад августа
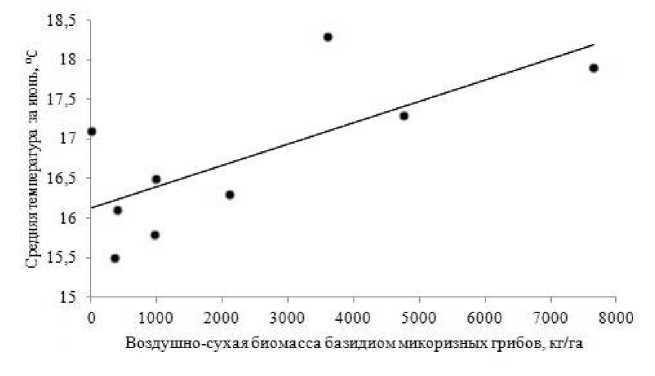
Рис. 5 . Диаграмма рассеяния средней температуры за июнь и воздушно-сухой биомассы базидиом эктомикоризных грибов, кг/га ( r s = 0.67; p < 0.05)
Анализируя пищевую ценность эктомикориз-ных грибов в ельнике кисличном, мы выявили, что 44 вида являлись съедобными. Эти грибы обычно имели крупные плодовые тела. Несъедобными были 29 видов, имеющих небольшие размеры бази-диом, либо обладающих неприятным запахом и вкусом. Ядовитых грибов обнаружено 7 видов.
Заключение
В ельнике кисличном за все время исследований выявлено 80 видов и внутривидовых таксонов агарикоидных микоризообразующих грибов, относящихся к 18 родам и 7 семействам, что составляет 37.4% от всей выявленной биоты агарикоидных базидиомицетов ельника кисличного. Лидирующими по числу видов эктомикоризных грибов за все время исследований были сем. Cortinariaceae , Russulaceae и Boletaceae , что подчеркивает бореальный характер исследуемой территории . Наибольшее число видов микоризных грибов отмечено в родах: Cortinarius , Russula , Lactarius и Amanita , что характерно для подзоны южной тайги. Новыми для ельника кисличного в третий период наблюдений являлись 19 видов микоризных грибов, из них 2 вида оказались новыми для Пермского края.
Видовой состав эктомикоризных грибов с течением времени меняется в большей степени (J= 32– 46), чем видовой состав высших сосудистых растений. Появление базидиом эктомикоризных грибов в климаксных сообществах не отличается стабильностью и зависит в большей степени не от растения-хозяина, а от внешних факторов. Особенно это заметно на доминантах грибов. Уста- снижают суммарную биомассу базидиом микоризных грибов к третьей декаде исследований (rs = -0.39; p < 0.05). При повышении средней месячной температуры воздуха в июле (rs = -0.49; p < 0.05) и августе (rs = -0.52; p < 0.05) наблюдается снижение числа базидиом эктомикоризных грибов.
новлено, что с течением времени происходит сближение видового состава эктомикоризных грибов разных периодов исследований, что свидетельствует о наличии мицелиального континуума во времени.
В изучаемом биоценозе выявлено 7 видов ми-коризообразователей, доминирующих по числу ба-зидиом, и 14 видов, доминирующих по биомассе. Видовой состав доминантов варьировал как по числу базидиом (J = 14–60), так и по их биомассе (J = 0–25). Ежегодно выявляемое число видов и продуктивность эктомикоризных грибов различается по годам наблюдений и зависит, чаще всего, от изменения погодных условий текущего года. Наибольшее число базидиом и их суммарная биомасса зафиксированы в 1977 и 1996 гг. Самыми «низкоурожайными» годами были 1975, 2010 и 2011 гг., так как являлись самыми засушливыми. Установлено, что повышение средней месячной температуры воздуха в июне благоприятствует «плодоношению» эктомикоризных грибов в августе ( r s = 0.67; p < 0.05). Выявлено, что средняя месячная температура воздуха и сумма осадков за август по годам наблюдений, в рамках нашего исследования, не влияет на «продуктивность» мико-ризообразователей данного ценоза.
Среди 80 видов эктомикоризных грибов, обнаруженных в ельнике кисличном, 44 вида являются съедобными, 29 – несъедобны, и 7 – ядовиты.
Список литературы Мониторинг эктомикоризных грибов ельника кисличного в южнотаежных лесах Пермского края
- Боталов В.С., Переведенцева Л.Г. Мониторинг эктомикоризных грибов сосняка чернично-сфагнового в южнотаежных лесах Пермского края // Вестник Пермского университета. Сер. Биология. 2018. Вып. 2. С. 107-116.
- Воронина Е.Ю. Симбиотрофные макромицеты и эктомикоризы основных пород в сложных ельниках на территории лесного массива Звенигородской биологической станции Московского государственного университета // Микология и фитопатология. 2004. Т. 38, вып. 3. С. 10-18.
- Грейг-Смит П. Количественная экология растений. М.: Мир, 1967. 359 с.
- Грибные сообщества лесных экосистем. М.; Петрозаводск, 2012. Т. 3. 192 с.; 2014. Т. 4. 145 с.; 2018. Т. 5. 163 с.
- Иванов А.И. Плодоношение агарикомицетов (Agaricomycetes) в природных сообществах Пензенской области в связи с циклами солнечной активности и погодными условиями // Микология и фитопатология. 2016. Т. 50, вып. 4. С. 219-229.
- Иванов А.И. Агарикомицеты Приволжской возвышенности. Порядок Boletales. Пенза, 2014. 178 с.
- Иллюстрированный определитель растений Пермского края / под ред. С.А. Овеснова. Пермь: Кн. мир, 2007. 743 с.
- Коваленко А.Е. Экологический обзор грибов из порядков Polyporales s. str., Boletales, Agaricales s. str., Russulales в горных лесах центральной части Северо-Западного Кавказа // Микология и фитопатология. 1980. Т. 14, вып. 4. С. 300-314.
- Леонтьев Д.В. Флористический анализ в микологии. Харьков: ПП РанокНТ, 2008. 110 с.
- Морозова О.В. Агарикоидные базидиомицеты подзоны южной тайги Ленинградской области: автореф. дис. … канд. биол. наук. СПб., 2001. 27 с.
- Новожилов Ю.К. и др. Скрытое разнообразие грибов и грибообразных протистов в природных экосистемах: проблемы и перспективы // Биосфера. 2016. Т. 8, № 2. С. 202-215.
- Переведенцева Л.Г. Биота и экология агарикоидных базидиомицетов Пермской области: автореф. дис. … д-ра биол. наук. М., 1999. 48 с.
- Переведенцева Л.Г., Шилкова Т.А. Микоризные агарикоидные базидиомицеты лесопарка "Черняевский" (г. Пермь) // Вестник Пермского университета. Сер. Биология. 2013. Вып. 3. С. 31-33.
- Смит С.Э., Рид Д.Дж. Микоризный симбиоз. М.: КМК, 2012. 776 с.
- Столярская М.В., Коваленко А.Е. Грибы Нижнесвирского заповедника. Вып. 1. Макромицеты: аннотированные списки видов. СПб., 1996. 59 с.
- Сукачев В.Н., Зонн Е.В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
- Трухачёва Н.В. Математическая статистика в медико-биологических исследованиях с применением пакета Statistica. М.: ГЭОТАР-Медиа, 2013. 384 с.
- Фомина Е.А. Эктомикоризные грибы еловых лесов Карельского перешейка (Ленинградская область). Видовое разнообразие // Микология и фитопатология. 2001. Т. 35, вып. 1. С. 43-51.
- Шубин В.И. О плодоношении эктомикоризных грибов // Хвойные бореальной зоны. 2009. Т. 26, № 1. С. 29-32.
- Andrew C., et al. Climate impacts on fungal community and trait dynamics // Fungal Ecology. 2016. Vol. 22. P. 17-25.
- Bochus G., Babos M. Coenology of terricolous macroscopic fungi of deciduous forests. Contributions to our knowledge of their behavior in Hungary // Bot. Jahrb. System. Pflanzengeschichte und Pflanzengeographie. 1960. Bd. 80, № 1. S. 1-100.
- De la Varga H., et al. Seasonal dynamics of Boletus edulis and Lactarius deliciosus extraradical mycelium in pine forests of central Spain // Mycorrhiza. 2013. Vol. 23, № 5. P. 391-402.
- Moser M. Die Rohrlinge und Blätterpilze (Polyporales, Boletales, Agaricales, Russulales) // Kleine Kryptogamenflora. Stuttgart; New York, 1983. Bd. 2b. 2. 533 S.
- MycoBank Database [Электронный ресурс]. URL: http//www.mycobank.org (дата обращения: 08.04.2019).
- Straatsma G., Ayer F., Egli S. Species richness, abundance, and phenology of fungal fruit bodies over 21 years in a Swiss forest plot // Mycological Research. 2001. Vol. 105, № 5. P. 515-523.
- Straatsma G., Krisai-Greilhuber I. Assemblage structure, species richness, abundance, and distribution of fungal fruit bodies in a seven-year plot-based survey near Vienna // Mycological Research. 2003. Vol. 107, № 5. P. 632-640.
