Мониторинг эктомикоризных грибов сосняка чернично-сфагнового в южно-таежных лесах Пермского края

Автор: Боталов В.С., Переведенцева Л.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
В Пермском крае с 1975 г. проводится мониторинг агарикоидных базидиомицетов в разных типах леса подзоны южной тайги. Исследования ведутся стационарным методом (пробные площади размером 50x20 м) в 3 этапа: I - 1975-1977 гг., II - 1994-1996 гг., Ш - 2010-2012 гг. Несмотря на высокую влажность в сосняке чернично-сфагновом, микориза у Pinus sylvestris L. и Betula pubescens Ehrh. хорошо развита, микоризные окончания сложного строения. К настоящему времени в сосняке чернично-сфагновом обнаружено 80 видов и внутривидовых таксонов агарикоидных базидиомицетов, 45 из которых (56.3 %) являются микоризообразователями, входящими в состав 7 семейств и 12 родов. Преобладают виды сем. Cortinariaceae и Russulaceae. Наибольшее число видов содержится в 4 родах: Cortinarius (21 вид), Lactarius (6), Hebeloma (4), Russula (3). Отмечено, что на фоне незначительных изменений видового состава сосудистых растений (коэффициенты Жаккара: JI-II = 83, JII-III = 82, JI-III = 75) и возрастания обводнения, произошла трансформация биоты микоризных грибов. Индексы общности, вычисленные по видовому составу грибов, колебались от 7 до 32. Микоризные грибы входят в состав доминантов по биомассе и числу базидиом. Изучена связь между разнообразием и продуктивностью микоризных грибов с основными климатическими факторами. Установлено отрицательное воздействие июньских осадков на плодоношение эктомикоризных грибов (rs = -0.70).
Эктомикоризные агарикоидные базидиомицеты, сосняк чернично-сфагновый, мониторинг, пермский край
Короткий адрес: https://sciup.org/147227006
IDR: 147227006 | УДК: 582.287.238
Monitoring of ectomycorrhizal fungi in the bilberry-sphagnum pine forest in the southern taiga of the Perm territory
The monitoring of agarics in the Perm Territory has been done since 1975 till the present time in the different types of forests (the southern taiga subzone). The research has been carried out by a stationary method (the test areas are 50x20 m) in 3 stages: I period - 1975-1977; II period - 1994-1996; Ш period - 2010-2012. Despite high humidity in the bilberry-sphagnum pine forest, mycorrhiza of Pinus sylvestris L. and Betula pubescens Ehrh. is well developed, mycorrhizal terminations are of complex structure. By the present time, 80 species and intraspecific taxa of agarics have been found in the bilberry-sphagnum pine forest, 45 of them (56.3 %) are ectomycorrhizal fungi. The mushrooms belong to 7 families and 12 genera. Families Cortinariaceae and Russulaceae prevail. The majority of species are in 4 genera: Cortinarius (41 species), Lactarius (6), Hebeloma (4), Russula (3). We have noted that biota of the mycorrhizal fungi has changed more than that of the higher plants (JI-II = 83, JII-III = 82, JI-III = 75). Jaccard index calculated by appearing mushrooms varies from 7 to 32. Mycorrhizal mushrooms are among dominant species in terms of their biomass and number. We have studied the connection between species diversity and productivity of agaricomycetes and the climate. We have found the negative impact of June rainfall on the > of ectomycorrhizal fungi (rs = -0.70).
Текст научной статьи Мониторинг эктомикоризных грибов сосняка чернично-сфагнового в южно-таежных лесах Пермского края
стоянием [Шубин, 1990; Переведенцева, 1999; Straatsma, Ayer, Egli, 2001].
Изучение биоты агарикоидных грибов в сообществах, не подверженных антропогенному воздействию, необходимо для познания их функционирования. Данную задачу позволяют решить многолетние стационарные наблюдения, отражающие динамику изменения микобиоты в биогеоценозах, однако подобные работы немногочисленны [Straatsma, Ayer, Egli, 2001; Lagana et al, 2002; Gange et al., 2018 и др.], а для такого обширного региона, как Урал – отсутствуют, что особенно актуально на фоне усиления антропогенного воздействия на природную среду.
В связи с этим, целью нашей работы является мониторинг биоты агарикоидных микоризообразующих грибов сосняка чернично-сфагнового. Для достижения цели были определены следующие задачи: 1) выявление видового состава эктомикориз-ных грибов сосняка чернично-сфагнового; 2) установление изменений их таксономической структуры во времени; 3) определение видов-доминантов по числу и биомассе базидиом; 4) выявление зависимости «плодоношения» грибов от количества осадков и температуры воздуха, в рамках изучения микобиоты.
Объекты и методы исследований
Изучение биоты агарикоидных базидиомицетов на территории Пермского края стационарным методом было начато в 1975 г. и продолжается до настоящего времени. Исследования проводятся в подзоне южной тайги в 10 типах леса, в том числе, в сосняке чернично-сфагновом (Добрянский административный район, окрестности ООПТ «Верхняя Кважва»; 58°38´ с.ш., 56°38´ в.д.). Первый период исследований был проведен в 1975–1977 гг., второй – в 1994–1996 гг., третий – в 2010–2012 гг. [Переведенцева, 2004; Грибные…, 2014].
С пробных площадей размером 50 × 20 м в течение августа (три посещения с интервалом в 10 дней) собирались все базидиомы агарикоидных грибов. Для каждого вида учитывалось их число и воздушно-сухая биомасса, отбирались образцы для последующей идентификации. Дополнительно в сентябре учитывался видовой состав грибов (одно посещение) [Переведенцева, 1985, 1999; Переве-денцева, Боталов, 2015].
Принадлежность к эколого-трофическим группам устанавливалась по литературным данным и наблюдениям в природе. За основу взята шкала эколого-трофических групп А.Е. Коваленко (с дополнениями) [Коваленко, 1980; Столярская, Коваленко, 1996]. К доминантам отнесены виды, составляющие 5% или более от общего числа плодовых тел или их суммарной биомассы за август [Bochus, Babos, 1960]. Геоботанические описания во все периоды выполнены согласно В.Н. Сукачеву и Е.В. Зонну [1961]. Латинские названия растений приводятся по «Иллюстрированному определителю растений Пермского края» [2007].
Для оценки сходства биогеоценозов по видовому составу грибов и сосудистых растений использовался коэффициент Жаккара [Шмидт, 1973]:
J =---c---xlOO, a + b - c где J – коэффициент сходства, c – число общих видов в двух сравниваемых ценозах; a, b – число видов в каждом из биогеоценозов. Пределы значений коэффициента Жаккара от 0 до 100%. При этом нулевое значение свидетельствует об отсутствии общих видов, а равное 100 – о полном сходстве видового состава.
Пробы корней древесных растений на исследование микосимбиотрофности были взяты один раз, в конце июля – начале августа у 5–10 экземпляров каждого вида. Обнаруженные нами типы микориз описывали согласно схеме структурно-функциональной классификации микориз и микоризоподобных образований, предложенной И.А. Селивановым (1981).
Для оценки влияния погодных условий на изменение продуктивности и числа выявляемых видов агарикоидных грибов, в рамках изучения микобиоты, проводился корреляционный анализ с использованием метода ранговой корреляции Спирмена ( r s ). Значение коэффициента корреляции ( r s ) от 0.3 до 0.7 означает среднюю положительную корреляцию между признаками; r s ≥ 0.70 – сильную положительную; отрицательное значение r s соответствует обратной корреляции. Результат считался статистически значимым при р-уровне меньше 0.05 [Трухачева, 2012]. Математическая обработка осуществлялась при помощи программ Microsoft Office Excel 2013 и StatSoft Statistica 10.
Рассчитывались корреляции климатических показателей со следующими характеристиками ми-коризообразователей: число базидиом и их суммарная биомасса за август; число видов за август–сентябрь, биомасса и число базидиом видов-доминантов, выявленных за август. В качестве климатических показателей использованы данные метеостанции г. Добрянка [Метеорологический…, 1975–1977, 1994–1996; Погода…, 2012]: сумма положительных температур (°C) за май–август, средняя месячная температура воздуха за май– сентябрь, сумма осадков (мм) по месяцам с мая по сентябрь, сумма осадков за май–август, сумма осадков по декадам августа.
Сосняк чернично-сфагновый располагается на равнинном плато, представляет собой облесенное верховое болото, коренное сообщество, возраст которого 95–105 лет. Состав древостоя 10С. Сомкнутость крон 0.4. Подрост состоит из Pinus sylvestris L. и Betula pubescens Ehrh. Подлесок не выражен. В кустарничково-травяном ярусе, проективное покрытие которого около 20%, преобладают Vaccinium vitis-idaea L. и Oxycoccus palustris
Pers., встречаются Chamaedaphne calyculata (L.) Moench и Ledum palustre L., различные виды осок. Моховой покров сплошной, образован сфагновыми мхами ( Sphagnum girgensohnii Russ., S. magellani-cum Brid. и др.). Валежника мало. Почва торфянистая, мощностью более 120 см.
С 1975 г. по настоящее время наблюдалось интенсивное развитие сфагновых мхов, что привело к возрастанию обводнения субстрата. Число видов высших сосудистых растений варьировало от 14 до 17, но их видовой состав по периодам исследований практически не изменялся ( J I–II = 83, J II–III = 82, J I–III = 75). Проективное покрытие кустарничковотравяного яруса сократилось от 50 (I период исследований) до 20% (II и III периоды). Доминирующие в I период Vaccinium myrtillus L., V. vitis-idaea L. и V. uliginosum L. во II и в III периоды остались лишь у стволов деревьев. Произошел прирост сфагновых мхов, застаивание воды и развитие разных видов осок. В III период продолжалось интенсивное нарастание сфагновых мхов.
По данным метеостанции г. Добрянка [Метеорологический…, 1975–1977, 1994–1996; Погода…, 2012] за время исследований (1975–1977, 1994– 1996 и 2010–2012 гг.) средние месячные температуры воздуха и количество осадков в летние месяцы отличались от средних многолетних данных. Для 2010–2012 гг., по сравнению с предыдущими наблюдениями, были зафиксированы более высокие температуры воздуха в июле и августе (особенно в 2010 и 2012 гг.). Больше всего осадков выпало в июле 1975 и 2010 гг., а меньше всего – в 1994 и 1996 гг. В остальные годы количество осадков за июль было близко к средним многолетним данным. В августе объем осадков чаще всего был меньше нормы, за исключением 1976, 1994 и 2010 гг. Таким образом, самыми засушливыми оказались 1975 и 2010 гг., а наиболее холодным и дождливым – 1994 г.
Результаты и их обсуждение
Видовой состав эктомикоризных агарикоидных базидиомицетов сосняка чернично-сфагнового не отличается большим разнообразием; далее приведен его список. Названия и объем родов, семейств указываются согласно системе, принятой М. Мозером [Moser, 1983], с некоторыми изменениями [Переведенцева, 1999]. Для каждого вида приводятся русское и латинское название. В скобках указаны синонимы видов в соответствии с современной классификацией [MycoBank Databases]. Римскими цифрами обозначены месяцы вегетационного периода. Хозяйственная значимость: съед. – съедобный, несъед. – несъедобный, яд. – ядовитый. Арабскими цифрами обозначен период выявления вида: 1 – 1975–1977 гг., 2 – 1994–1996 гг., 3 – 2010–2012 гг.
Аннотированный список видов грибов
AGARICALES Clements – Агарикальные
Amanitaceae R. Heim ex Pousar – мухоморовые: Amanita fulva Pers. – поплавок желто-коричневый, VII–VIII, съед., (1, 3); A. porphyria Alb. & Schwein. – мухомор порфировый, VIII–IX, яд., (1).
Cortinariaceae R. Heim ex Pouzar – паутинни-ковые: Cortinarius acutus (Pers.) Fr. – паутинник заостренный, под хвойными, VIII, несъед., (3); C. alboviolaceus (Pers.) Fr. – п. бело-фиолетовый, под лиственными, VII–VIII, съед., (1); C. armeniacus (Schaeff.) Fr. – п. абрикосовооранжевый, под хвойными и лиственными, VII– VIII, съед., (1); C. bataillei J. Favre – п. желто-огненноножковый, VIII–IX, несъед., (1, 2); C. brunneus (Pers.) Fr. – п. темно-бурый, под хвойными, VIII–IX, несъед., (1); C. candelaris Fr. – п. свечковидный, VIII, несъед., (2); C. cinnamomeoluteus P.D. Orton – п. коричневожелтый, под хвойными деревьями, VIII–IX, не-съед., (3); C. cinnamomeus (L.) Gray – п. темнокоричневый, VIII–IX, несъед., (3); C. collinitus (Sowerby) Gray – п. пачкающий, VIII–IX, съед., (1, 2); C. croceus (Schaeff.) Gray – п. шафрановый, VIII–IX, несъед., (1, 3); C. gentilis (Fr.) Fr. – п. благородный, под хвойными деревьями, VI–VIII, не-съед., (1); C. hemitrichus (Pers.) Fr. – п. полуопу-шенный, VII–VIII, несъед., (1, 3); C. huronensis Ammirati & A.H.Sm. – п. гуронский, VIII, несъед., (1, 2); C. jubarinus Fr. – п. сияющий, VIII, несъед., (3); C. leucopus (Bull.) Fr. – п. белоножковый, VIII, несъед., (3); C. malachius (Fr.) Fr. – п. малахитовый, VIII–IX, несъед., (1); C. semisanguineus (Fr.) Gillet – п. кроваво-красноватый, VIII–IX, несъед., (1, 2); C. sphagneti Singer – п. сфагновый, VIII–IX, несъед., (3); C. trivialis J.E. Lange – п. обыкновенный, под лиственными, VIII–IX, несъед., (1); C. tubarius Ammirati & A.H. Sm. – п. сфагновый, VIII–IX, несъед., (3); C. uliginosus Berk. – п. топяной, VIII–IX, несъед., (2); Hebeloma candidipes Bruchet – гебелома белоногая, VIII, несъед, (3); H. claviceps (Fr.) Quél. – г. булавовидноногая, VIII–IX, несъед., (1, 3); H. crustuliniforme (Bull.) Quél. – г. клейкая, ложный валуй, под xвойными и лиственными деревьями, VII–IX, несъед., (1, 2); H. longicaudum (Pers.) P. Kumm. – г. длиннонож-ковая, под хвойными и лиственными деревьями, VII–IX, несъед., (1); Rozites caperatus (Pers.) P. Karst. (= Cortinarius caperatus (Pers.) Fr.) – колпак кольчатый, под хвойными, VIII–IX, съед., (1).
Tricholomataceae R. Heim ex Pousar– трихоло-мовые, рядовковые: Collybia butyracea var. butyracea (Bull.) Fr. (= Rhodocollybia butyracea (Bull.) Lennox) – коллибия масляная, VII–IX, съед., (1); Laccaria laccata (Scop.) Cooke – лакови-ца лаковая, VI–IX, съед., (2); L. proxima (Boud.) Pat. – л. рыжеватая, VIII, съед., (2).
BOLETALES J.-E. Gilbert – Болетальные
Boletaceae Chevall. – трубчатые, болетовые: Leccinum scabrum (Bull.) Gray – подберезовик обыкновенный, VI–IX, съед., (1, 2); Suillus flavidus (Fr.) Singer – масленок желтоватый, болотный, VIII, съед., (2); S. variegatus (Sw.) Kuntze – м. желто-бурый, VIII–IX, съед., (1, 2).
Gomphidiaceae Maire ex Jülich – мокруховые: Chroogomphus rutilus (Schaeff.) O.K. Mill. – хро-огомф пурпуровый, VI–IX, съед., (1, 2).
Paxillaceae Lotsy – паксилловые, свинушковые: Paxillus involutus (Batsch) Fr. – свинушка тонкая, под хвойными и лиственными, иногда на гнилой древесине, VIII–IX, яд., (1–3).
RUSSULALES Kreisel ex Kirk et al. – Руссу-лальные
Russulaceae Lotsy – сыроежковые: Lactarius flexuosus (Pers.) Gray (= L. zonarius (Bull.) Fr.) – млечник извилистый, серушка, под лиственными и хвойными деревьями, VII–VIII, съед., (1); L. helvus (Fr.) Fr. – м. серо-розовый, под хвойными деревьями, VIII–IX, несъед., (1); L. rufus (Scop.) Fr. – горькушка, под хвойными и лиственными деревьями, VII–IX, съед., (1–3); L. theiogalus (Bull.) Gray – м. серно-млечный, золотистый, VIII– IX, съед., (1); L. trivialis (Fr.) Fr. – м. обыкновенный, гладыш, под хвойными, VII–VIII, съед., (1); L. vietus (Fr.) Fr. – м. блеклый, под лиственными деревьями, VIII–IX, съед., (1); Russula decolorans (Fr.) Fr. – сыроежка сереющая, Mr, VII–VIII, съед., (1); R. emetica (Schaeff.) Pers. – с. жгучеедкая, VII–IX, несъед., (1, 2); R. paludosa Britzelm. – с. болотная, под хвойными деревьями, VII–VIII, съед., (1, 2).
Анализ таксономической структуры
Несмотря на высокую влажность в сосняке чернично-сфагновом, микориза у древесных растений развита хорошо. Микоризные окончания находятся в верхних слоях торфа, а также в слаборазложив-шихся сфагновых мхах. Характерной особенностью является то, что микоризные окончания сложного строения, имеют большую массу. Нередки гроздевидные, коралловидные окончания у Pinus sylvestris, а булавовидные встречаются реже [Переведенцева, 1999].
У сосны обыкновенной 55% микоризных окончаний имеют чехлы подтипа В1. Довольно много чехлов сложного строения F1, F3 (30%). Мицелиальные чехлы, образованные Cenococcum geophi-lum Fr., составляют 11%, то есть, в условиях сильного увлажнения большую роль в микоризообра-зовании играют другие виды грибов, в частности, агарикоидные базидиомицеты, например виды рода Lactarius . Следует отметить еще одну особенность в образовании мицелиальных чехлов у сосны обыкновенной в сосняке чернично-сфагновом. Только в переувлажненных местообитаниях у нее обнаружены чехлы подтипа J. У Betula pubescens в образовании микориз Cenococcum geophilum принимает меньшее участие, чем у сосны, так как выявлено около 10% чехлов подтипа G. Другие микоризные грибы образуют чехлы подтипа В1 (82%). Небольшая часть чехлов, имеющих сложное строение, относится к подтипу F3.
В сосняке чернично-сфагновом за все время исследований (1975–1977, 1994–1996, 2010–2012 гг.) выявлено 80 видов и внутривидовых таксонов ага-рикоидных грибов, из которых 45 являются мико-ризообразователями, относящимися к 12 родам и 7 семействам (таблица). В изучаемом биогеоценозе микоризообразователи являются преобладающей эколого-трофической группой, к которой относится 56.3% всех выявленных видов агарикоидных грибов. Большей частью, это симбионты сосны обыкновенной и березы пушистой [Шубин, 1990; Смит, Рид, 2012]. Преобладание в составе микобиоты микоризообразователей характерно для всей лесной зоны Голарктики [Нездойминого, 1996; Moser, 1983; Funga Nordica, 2012]. В III период наблюдений выявлено 8 видов грибов, новых для изучаемого ценоза, из которых Cortinarius leucopus впервые обмечен на территории Пермского края.
Таксономический состав эктомикоризных агарикоидных грибов сосняка чернично-сфагнового
|
Порядок |
Семейство (число родов/видов) |
Роды (с указанием числа видов и внутривидовых таксонов) |
|
Boletales |
Boletaceae (2/3) |
Leccinum (1), Suillus (2) |
|
Paxillaceae (1/1) |
Paxillus (1) |
|
|
Gomphidiaceae (1/1) |
Chroogomphus (1) |
|
|
Agaricales |
Tricholomataceae (2/3) |
Laccaria (2), Collybia (1) |
|
Amanitaceae (1/2) |
Amanita (2) |
|
|
Cortinariaceae (3/26) |
Cortinarius (21), Hebeloma (4), Rozites (1) |
|
|
Russulales |
Russulaceae (2/9) |
Russula (3), Lactarius (6) |
|
Всего |
7 семейств |
12 родов (45 видов) |
Наиболее крупным по числу видов микоризных грибов является порядок Agaricales (31 вид). Ведущими по числу видов оказались семейства:
Cortinariaceae (26 видов) и Russulaceae (9), к которым относится 77.8 % всех выявленных видов микоризообразователей. Наибольшим числом ви- дов представлены роды: Cortinarius (46.7% от всех видов микоризных грибов), Lactarius (13.4%), Hebeloma (9.0%) и Russula (6.7%). Преобладание в составе микобиоты представителей сем. Cortinariaceae, а также родов Cortinarius, Russula и Lactarius характерно для сосновых лесов Пермского края [Переведенцева, 1999, 2008; Переве-денцева, Боталов, 2015].
Наличие или отсутствие базидиом разных видов агарикоидных грибов зависит от различных факторов, поэтому в ходе ежегодных наблюдений выявляется только некоторая часть реально существующих в биогеоценозе видов [Переведенцева, 1999; Новожилов и др., 2016]. В связи с этим, все виды грибов, обнаруживаемые на учетной площади, включались в общий список. В результате отмечено увеличение общего выявленного разнообразия микоризообразователей по годам наблюдений, хотя ежегодно выявляемое число их видов со II периода сократилось (рис. 1). Установлено несоответствие наблюдаемого и скрытого разнообразия биоты эктомикоризных грибов.
Анализируя видовое разнообразие грибов по годам наблюдений, отметим, что минимальное число видов выявлено в засушливый 2011 г. (4 вида, или 8.9% от общего числа видов микоризных грибов), максимальное – в 1977 г. (25 видов, или 55.6%) и в 1976 г. (24 вида, или 53.3%).
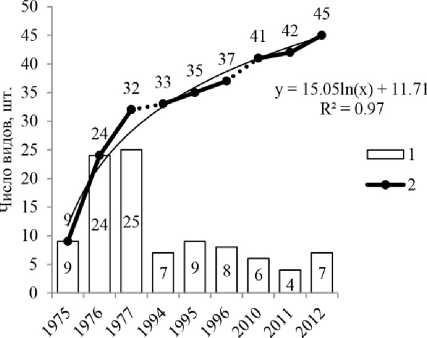
Рис. 1 . Изменение видового разнообразия биоты эктомикоризных грибов по годам наблюдений:
1 – число видов за год наблюдений; 2 – накопление общего выявленного разнообразия (аппроксимация логарифмической функцией)
Новый для Пермского края, Cortinarius leucopus, обнаружен в 2010 г. Показательно, что за всё время наблюдений, 20 видов грибов (44.4% от общего числа микоризных грибов) были зафиксированы только один раз в какой-либо сезон (рис. 2).
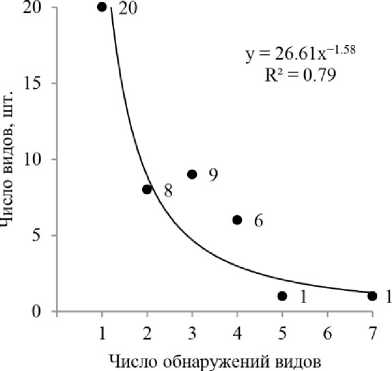
Рис. 2. Выявление видов эктомикоризных грибов в сосняке чернично-сфагновом за все время исследований (аппроксимация степенной функцией)
Постоянными для сосняка чернично-сфагнового являются Lactarius rufus (1975–1977, 1995, 1996, 2011, 2012 гг.) и Suillus variegatus (1976, 1977, 1994–1996 гг.). Ежегодно встречающихся видов не было выявлено.
Сравнение видового состава микоризообразователей по периодам наблюдений
В сосняке чернично-сфагновом со временем происходили изменения в составе ведущих семейств грибов (рис. 3) . Отметим, что в каждый период лидирующее положение занимали виды сем. Cortinariaceae и Russulaceae. Другие семейства были представлены в различной степени. Так, в I период доли сем. Boletaceae и Tricholomataceae составляли 6.3 и 3.1% соответственно, во II период их было по 11.7% от числа видов за период. В 2010–2012 гг., на фоне усиления болотообразовательного процесса, представителей указанных семейств не выявлено. Остальные семейства в различные периоды составляли 11.7–14.2%.
Со временем выявлены изменения в спектре ведущих родов. В I и II периоды наибольшее число видов относилось к роду Cortinarius (37.5 и 35.4% от числа видов микоризных грибов за период). В I период в спектре ведущих еще были роды – Lactarius (18.8%), Hebeloma и Russula (по 9.4%), а во II период – Russula, Laccaria и Suillus (по 11.8%). В III период также преобладал род Cortinarius (64.4 %), на втором месте был род Hebeloma (14.3%). Роды Russula, Laccaria и Suillus в III период выпали из состава микобиоты. В каждый период наблюдений выявлялись эврибионт-ные виды: Paxillus involutus, Lactarius rufus , Suillus variegatus и некоторые другие, массово встречающиеся в разных типах сосновых лесов, как в пере-
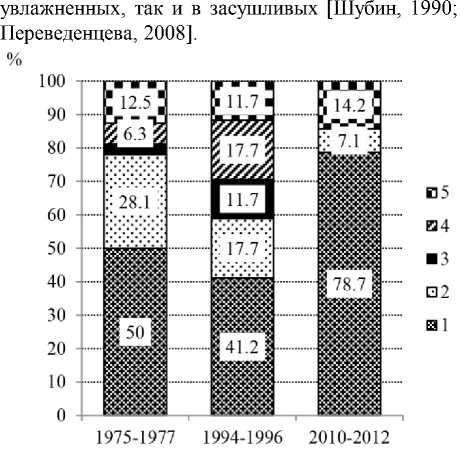
Рис. 3 . Соотношение ведущих семейств эктоми-коризных грибов в сосняке чернично-сфагновом по периодам наблюдений (в процентах от числа видов микоризообразователей за период):
1 – Cortinariaceae , 2 – Russulaceae , 3 – Tricholo-mataceae , 4 – Boletaceae , 5 – остальные семейства
Проводя сравнение видового состава эктомико-ризных грибов по периодам наблюдений, установили, что с увеличением обводнения число видов микоризных грибов уменьшается. Так, в I период было выявлено 32 вида (71.1% от общего числа видов микоризообразователей). Во II – число видов сократилось до 17 (37.8%), а в III период – до 14 (31.1%). Но тем не менее, в каждый период наблюдений микоризообразователи составляли 52.5– 63.7% от всех видов, то есть доли их в составе микобиоты различных периодов колебались примерно на 11%, хотя видовой состав между периодами существенно различается ( J I–II = 32; J II–III = 7; J I–III = 15). Постепенное снижение числа выявляемых видов грибов, смена видового состава микоризообра-зователей, выпадение из состава микобиоты в III период представителей семейств Boletaceae и Tricholomataceae , а также изменение спектра ведущих родов, в значительной степени связаны с усилением болотообразовательного процесса.
Доминирующие виды грибов
Доминанты имеют наибольшие показатели количественной представленности в ценозе, поэтому данный признак рассматривают в качестве индикатора экосистемной роли различных видов грибов, а изменение их видового состава может свидетельствовать об устойчивости видов к факторам среды, о происходящих в биогеоценозе сукцессионных процессах [Переведенцева, 1999; Леонтьев, 2008; Bochus, Babos, 1960].
В изучаемом ценозе за все время исследований выявлено 10 видов микоризных грибов, доминирующих по числу базидиом (58.8% от общего количества доминантов по числу базидиом), и 13 видов – по биомассе базидиом (76.5% от общего числа доми-нантов по биомассе). Микоризные грибы преобладают не только по числу видов, но и по биомассе ба-зидиом, которая составляла 77.5% от биомассы ба-зидиом всех доминирующих видов грибов. Как по биомассе, так и по числу базидиом, в разные годы, доминировали: Cortinarius cinnamomeoluteus, C. croceus, C. tubarius, Russula emetica, R. paludosa, Lactarius rufus, L. helvus, Suillus variegatus и другие. Число видов-доминантов микоризообразова-телей по биомассе базидиом варьировало по периодам от 3 до 8, по числу базидиом – от 2 до 5.
Выявлено, что видовой состав доминантов-микоризообразователей по числу базидиом со временем изменяется ( J I–II = 20; J II–III = 0; J I–III = 29). Доминантами по биомассе базидиом, чаще всего, были одни и те же виды ( J I–II = 50; J II–III = 38; J I–III = 75).
«Плодоношение» эктомикоризных агарикоидных грибов
Микоризные грибы составляют 88.2% от общей биомассы базидиом и 27.3% – от общего числа бази-диом. Самым «урожайным» по биомассе базидиом был I период (15.0 кг/га), когда наблюдалось массовое развитие грибов с крупными плодовыми телами: Lactarius helvus, L. rufus, Russula emetica, R. paludosa , и другие. В остальные периоды происходит постепенное снижение продуктивности микоризных грибов из-за продолжающегося заболачивания ценоза. По числу базидиом максимальные «урожаи» были в I период (16080 шт./га), минимальные – во II период (76 шт./га). Самыми неблагоприятными для развития микоризных грибов оказались 2010 и 2011 гг., а самыми «урожайными» – 1976 и 1977 гг.
На плодоношение агарикоидных грибов оказывают влияние различные факторы. Чаще всего колебания обилия и разнообразия агарикоидных грибов объясняют погодными условиями вегетационного периода [Шубин, 1990; Музыка, Музыка, 2015; Иванов, 2016; Straatsma, Ayer, Egli, 2001; Büntgen et al., 2015; Tahvanainen et al., 2016]. В изучаемом биогеоценозе выявлено немного значимых корреляций характеристик микобиоты с погодными условиями текущего года. Так, с увеличением количества осадков в июне, сокращается число базидиом микоризных грибов в августе (rs = –0.70), а также число их видов за август–сентябрь (rs = –0.72) (рис. 4), что связано с аккумуляцией дождевой воды сфагновыми мхами [Wieder, Vitt, 2006]. Осадки во время учета числа и биомассы базидиом (август), в рамках нашего исследования, не влияют на продуктивность микоризообразова-телей данного ценоза. Повышение средней месячной температуры воздуха в июле способствует снижению «урожаев» микоризных грибов как по числу базидиом (rs = –0.45), так и по их биомассе (rs = –0.60). Для видов-доминантов выявлены закономерности, аналогичные общим характеристикам микобиоты.
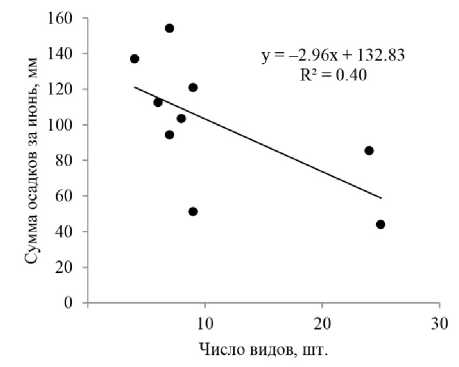
Рис. 4 . Диаграмма рассеяния суммы осадков за июнь и числа видов микоризообразователей за август-сентябрь ( r s = –0.72; p = 0.028)
Многие микоризные грибы известны как съедобные и обычно имеют крупные базидиомы. В сосняке чернично-сфагновом 19 видов относятся к съедобным грибам. Ядовитыми грибами являются Amanita porphyria и Paxillus involutus , а 24 вида грибов относятся к несъедобным. Они имеют либо небольшие размеры плодовых тел, либо обладают неприятным вкусом и запахом, либо их пищевые свойства неизвестны [Переведенцева, 2008].
Заключение
В сосняке чернично-сфагновом за все время исследований выявлено 45 видов и внутривидовых таксонов агарикоидных микоризообразующих грибов, относящихся к 12 родам и 7 семействам, что составляет 56.3% от всей выявленной биоты ага-рикоидных базидиомицетов. Лидирующими по числу видов микоризных грибов как за все время исследований, так и в каждый период, были сем. Cortinariaceae и Russulaceae . Наибольшее число видов грибов отмечено в родах: Cortinarius , Russula и Lactarius .
На фоне относительной стабильности видового состава высших сосудистых растений ( J I–II = 83; J II– III = 82; J I–III = 75) и усиления болотообразовательного процесса выявлена трансформация биоты ми-коризообразователей ( J I–II = 32; J II–III = 7; J I–III = 15), а также изменение структуры ведущих семейств и родов.
Микоризные грибы составляют 58.8% от числа видов-доминантов по числу базидиом и 76.5% – от числа видов-доминантов по биомассе базидиом. Число видов-доминантов микоризообразователей по биомассе базидиом варьировало по периодам от 3 до 8, по числу базидиом – от 2 до 5. Видовой состав доминантов по числу базидиом со временем изменяется (JI–II = 20; JII–III = 0; JI–III = 29). По биомассе базидиом доминантами, чаще всего, были одни и те же виды (JI–II = 50; JII–III = 38; JI–III = 75).
Зафиксировано снижение «урожаев» микоризо-образователей во II и III периоды наблюдений. Ежегодно выявляемое число видов и продуктивность микоризных грибов различается по годам наблюдений и зависит от изменения погодных условий текущего года. При увеличении количества осадков в июне установлено снижение «урожаев» микоризообразователей в августе, а также числа их видов в августе-сентябре ( r s ≈ –0.70; p < 0.05). Повышение средней месячной температуры воздуха в июле также ослабляет их «плодоношение». Среди микоризных грибов 19 видов являются съедобными, 24 вида несъедобны и 2 – ядовиты.
Список литературы Мониторинг эктомикоризных грибов сосняка чернично-сфагнового в южно-таежных лесах Пермского края
- Грибные сообщества лесных экосистем / ред. В.Г. Стороженко, А.В. Руоколайнен. М.; Петрозаводск, 2014. Т. 4. 145 с
- Иванов А.И. Плодоношение агарикомицетов (Agaricomycetes) в природных сообществах Пензенской области в связи с циклами солнечной активности и погодными условиями // Микология и фитопатология. 2016. Т. 50, вып. 4. С. 219-229
- Иллюстрированный определитель растений Пермского края / ред. С.А. Овеснов. Пермь: Кн. мир, 2007. 743 с
- Коваленко А.Е. Экологический обзор грибов из порядков Polyporales s. str., Boletales, Agaricales s. str., Russulales в горных лесах Центральной части Северо-Западного Кавказа // Микология и фитопатология. 1980. Т. 14, вып. 4. С. 300314
- Леонтьев Д.В. Флористический анализ в микологии. Харьков: ПП Ранок-НТ, 2008. 110 с
- Метеорологический ежемесячник. Уральское УГМС, Свердловск. 1975-1977, 1994-1996. Вып. 9
- Музыка В.А., Музыка С.М. Метеозависимость в развитии микоризных макромицетов и прогнозирование ежегодных урожаев съедобных грибов на примере Северного Присаянья // Вестник ИрГСХА. 2015. № 67. С. 72-78
- Нездойминого Э.Л. Определитель грибов России. Порядок Агариковые. Вып. 1. Семейство Пау-тинниковые. СПб.: Наука, 1996. 408 с
- Новожилов Ю.К. и др. Скрытое разнообразие грибов и грибообразных протистов в природных экосистемах: проблемы и перспективы // Биосфера. 2016. Т. 8, № 2. С. 202-215.
- Переведенцева Л.Г. Микоризные грибы и мико-симбиотрофизм растений в сосновых лесах Центрального Прикамья // Микориза и другие формы консортивных связей в природе. Пермь, 1985. С. 59-66
- Переведенцева Л.Г. Биота и экология агарикоидных базидиомицетов Пермской области: авто-реф. дис.... д-ра биол. наук. М., 1999. 48 с
- Переведенцева Л.Г. Агарикоидные микоризообра-зующие грибы Пермского Прикамья // Вестник Пермского университета. 2004. Вып. 2. Биология. С. 14-18
- Переведенцева Л.Г. Мониторинг видового состава агарикоидных микоризных грибов Пермского Прикамья // Высшие базидиальные грибы: индивидуумы, популяции, сообщества: материалы юбилейной конф., посвящ. 100-летию со дня рождения М.В. Горленко. М., 2008. С. 143-148
- Переведенцева Л.Г., Боталов В.С. Мониторинг видового состава агарикоидных базидиомице-тов в некоторых типах сосновых лесов Пермского края (подзона южной тайги) // Проблемы лесной фитопатологии и микологии: материалы 9-й Междунар. конф. Минск, 2015. С. 163-166
- Погода и климат [Электронный ресурс]. URL: http://www.pogodaiklimat.ru (дата обращения: 01.12.2012)
- Селиванов И.А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 231 с
- Смит С.Э., Рид Д.Дж. Микоризный симбиоз. М.: КМК, 2012. 776 с
- Столярская М.В., Коваленко А.Е. Грибы Нижнесвир-ского заповедника. Вып. 1. Макромицеты: аннотированные списки видов. СПб., 1996. 59 с
- Сукачев В.Н., Зонн Е.В. Методические указания к изучению типов. М.: Изд-во АН СССР, 1961. 144 с
- Трухачева Н.В. Математическая статистика в медико-биологических исследованиях с применением пакета Statistica. М.: ГЭОТАР-Медиа, 2012. 384 с
- Шмидт В.М. Математические методы в ботанике. Л.: Наука, 1973. 263 с
- Шубин В.И. Макромицеты лесных фитоценозов таежной зоны и их использование. Л.: Наука, 1990. 195 с
- Bochus G., Babos M. Coenology of terricolous macroscopic fungi of deciduous forests. Contributions to our knowledge of their behavior in Hungary // Botanische Jahrbucher fur Systematik, Pflan-zengeschichte und Pflanzengeographie. 1960. B. 80, № 1. P. 1-100
- Buntgen U. et al. Drought-induced changes in the phenology, productivity and diversity of Spanish fungi // Fungal Ecology. 2015. Vol. 16. P. 6-18
- Funga Nordica / Edit. H. Knudsen & J. Vesterholt. Copenhagen: Nordsvamp. 2012. Vol. 1, 2
- Gange A.C. et al. Trait-dependent distributional shifts in fruiting of common British fungi // Ecography. 2018. Vol. 41, I. 1. P. 51-61
- Lagana A. et al. Macrofungi as long-term indicators of forest health and management in central Italy // Cryptogamie Mycologie. 2002. Vol. 23, № 1. P. 39-50
- Moser M. Die Rohrlinge und Blatterpilze (Polypora-les, Boletales, Agaricales, Russulales). Kleine Kryptogamenflora. Bd. 2b/2. Stuttgart, New York: Fischer, 1983. 533 S
- MycoBank Database [Электронный ресурс]. URL: http//www.mycobank.org (дата обращения: 15.03.2018)
- Straatsma G., Ayer F., Egli S. Species richness, abundance, and phenology of fungal fruit bodies over 21 years in a Swiss forest plot // Mycological Research. 2001. Vol. 105, № 5. P. 515-523
- Tahvanainen, V. et al. Modelling the yields of marketed mushrooms in Picea abies stands in eastern Finland // Forest Ecology and Management. 2016. Vol. 362. P. 79-88
- Wieder R.K., Vitt D.H. Boreal Peatland Ecosystems. Berlin: Springer, 2006. 188 p
