Мониторинг эктомикоризных грибов в некоторых типах еловых лесов (подзона южной тайги, Пермский край, Россия)
")
Автор: Шишигин А.С., Боталов В.С.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микология
Статья в выпуске: 1, 2025 года.
Бесплатный доступ
Подведены итоги многолетнего мониторинга биоты эктомикоризных грибов в некоторых типах еловых лесов Пермского края. Исследования проводились стационарным методом на учетных площадях размером 50 × 20 м, заложенных по одной в исследуемых типах леса - ельнике приручьевом и ельнике кисличном. Работа осуществлялась в три периода: I - 1975-1977 гг., II - 1994-1996 гг., III - 2010-2012 гг. В каждый год наблюдений по 4 раза с интервалом в 10 дней собирались все плодовые тела грибов, учитывалось их число и воздушно-сухая биомасса. К настоящему времени выявлено от 80 (ельник кисличный) до 124 (ельник приручьевой) видов и внутривидовых таксонов эктомикоризных грибов. Большинство выявленных видов относится к семействвам Cortinariaceae, Russulaceae и Tricholomataceae, что характерно для бореальной зоны. Наибольшее число выявленных видов (72.6-75.0%) обнаруживалось от 2 до 9 раз, а 2-3% из них были постоянными, встречались ежегодно. Отмечена относительная стабильность видового состава сосудистых растений (коэффициенты Жаккара (J×100): J = 69-88) во времени и зафиксированы более значительные изменения видового состава микоризообразователей (J = 32-52). Продуктивность эктомикоризных грибов в исследуемых ценозах различается по годам и периодам наблюдений. Наибольшее число базидиом зафиксировано для ельника кисличного, а их наибольшая биомасса - для ельника приручьевого. Для биоты микоризообразователей как по числу (индекс Шеннона: H = 1.27), так и по биомассе (H = 1.54) базидиом за все периоды наблюдений более благоприятным являлся ельник приручьевой. Установлено, что повышение средней месячной температуры воздуха в июне благоприятствует «плодоношению» микоризообразователей в августе в ельнике кисличном (rs = 0.67; p function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Эктомикоризные агарикоидные базидиомицеты, мониторинг, ельник приручевой, ельник кисличный, экология грибов
Короткий адрес: https://sciup.org/147251169
IDR: 147251169 | УДК: 582.287.238 | DOI: 10.17072/1994-9952-2025-1-89-102
Monitoring of ectomycorrhizal fungi in some types of spruce forests (subzone of the southern taiga, Perm territory, Russia)
The results of long-term monitoring of ectomycorrhizal fungi biota in some types of spruce forests of Perm Krai are summarized. The studies were conducted using a stationary method on 50 × 20 m accounting plots, one in each of the studied forest types: spruce forest at the brook, sorrel spruce forest. The work was carried out in three periods: I - 1975-1977, II - 1994-1996, III - 2010-2012. In each year of observations, all fruiting bodies of fungi were collected 4 times with an interval of 10 days, their number and air-dry biomass were taken into account. To date, from 80 (sorrel spruce forest) to 124 (spruce forest at the brook) species and intraspecific taxa of ectomycorrhizal fungi have been identified in the studied biogeocenoses. Most of the identified 90 species belong to the families Cortinariaceae, Russulaceae and Tricholomataceae, which is typical of the boreal zone. The greatest number of identified species (72.6-75.0%) were found from 2 to 9 times, and 2-3% of them were constant and occurred annually. Relative stability of the species composition of higher vascular plants (Jaccard index×100: J = 69-88) over time was noted, and more significant changes in the species composition of mycorrhiza-forming fungi (J = 32-52) were recorded. The productivity of ectomycorrhizal fungi in the studied cenoses varies by years and observation periods. The largest number of basidiomes was recorded for wood sorrel spruce forest, and their largest biomass was recorded for spruce forest at the brook. The spruce forest at the brook was the most favorable for the biota of ectomycorrhizal fungi, both in number (Shannon index: H = 1.27) and in biomass (H = 1.54), during all the research, since the biota of ectomycorrhizal fung of the specified cenosis was more diverse and its components were most aligned. It was found that an increase in the average monthly air temperature in June favors the "fruiting" of mycorrhiza-forming fungi in August in the wood sorrel spruce forest (rs = 0.67; p function show_eabstract() { $('#eabstract1').hide(); $('#eabstract2').show(); $('#eabstract_expand').hide(); }
Текст научной статьи Мониторинг эктомикоризных грибов в некоторых типах еловых лесов (подзона южной тайги, Пермский край, Россия)
Агарикоидные базидиомицеты играют огромную роль в функционировании природных экосистем. Особую значимость из них представляют эктомикоризные грибы, вступающие с высшими растениями в консортивные отношения в форме микосимбиотрофизма. Микоризообразователи распространены в лесных экосистемах и составляют значительную часть от общего числа обитающих в них агарикоидных базидиомицетов [Переведенцева, 1999; Фомина, 2000, 2001; Straatsma, Ayer, Egli, 2001; Straatsma, Krisai-Greilhuber, 2003; Воронина, 2004; Шубин, 2009; Botalov, Perevedentseva, Shishigin, 2018, 2020; Шишигин, 2021]. «Плодоношение» эктомикоризных грибов является показателем дополнительного получения углеводов и повышения интенсивности фотосинтеза у древесных растений [Шубин, 2009]. Все микоризообразующие грибы оказывают существенное влияние на минеральное питание и водный баланс древесных растений, а также играют огромную роль в биохимических циклах важнейших химических элементов в лесных биогеоценозах. Они задействованы в круговороте азота, фосфора и калия [Селиванов, 1981; Шубин, 2010; Смит, Рид, 2012; Иванов, 2014, 2016].
Большой интерес представляют исследования эктомикоризных грибов в климаксных сообществах, которые имеют довольно стабильный видовой состав высших растений, в особенности древесных пород. В подзоне южной тайги Пермского края климаксными сообществами являются еловые леса. В связи с этим целью нашей работы является мониторинг эктомикоризных грибов в ельнике приручьевом и ельнике кисличном. Для достижения цели были поставлены следующие задачи: 1) выявление видового состава экто-микоризных грибов в еловых лесах; 2) проведение анализа таксономической структуры микоризообразова-телей; 3) определение доминирующих видов грибов по числу базидиом и их биомассе; 4) выявление зависимости «плодоношения» эктомикоризных грибов от количества осадков и температуры воздуха.
Материал и методы исследования
В Пермском крае (Добрянский городской округ, ООПТ «Верхняя Кважва») в лесных ценозах с 1975 г. проводится мониторинг агарикоидных базидиомицетов стационарным методом, позволяющим количественно оценить и выявить структуру грибного компонента, что необходимо для понимания развития природных экосистем. Микоэкологические наблюдения нами проводились в 2010–2012 гг. в ельнике приручь-евом и ельнике кисличном подзоны южной тайги Пермского края. Для выявления изменений, происходящих в структуре биоты эктомикоризных грибов за продолжительный период времени, полученные нами данные (III – период: 2010–2012 гг.) сравнивались с результатами предыдущих наблюдений, проведенных Л.Г. Переведенцевой [1999] (I – период: 1975–1977; II – период: 1994–1996 гг.). Геоботаническое описание было выполнено согласно В.Н. Сукачёву, Е.Н. Зонну [1961]. Латинские названия сосудистых растений приводятся по Иллюстрированному определителю растений Пермского края [2007].
Эколого-ценотическая характеристика исследуемых биогеоценозов
Ельник приручьевой (58°23‘6" с.ш.; 56°21‘55" в.д.) - коренное сообщество, возраст древостоя которого составляет 120–135 лет. Биогеоценоз расположен в долине лесной р. Кважевки. Состав леса 9E1П+Б. Сомкнутость крон 0.5. Подрост образован Picea obovata Ledeb и Abies sibirica Ledeb. Кустарниковый ярус представлен такими видами, как Lonicera xylosteum L., Sorbus aucuparia L., Ribes nigrum L. и R. spi-catum Robson. Проективное покрытие кустарничково-травяного яруса составляет 80–90%, где преобла- дают Dryopteris carthusiana Vill., Oxalis acetosella L. и Stellaria nemorum L. Моховой покров представлен зелёными мхами, растущими на стволах и около стволов деревьев, реже они встречаются на почве. Валежника много. Почва пойменная, аллювиально-слоистая, легкосуглинистая.
Ельник кисличный (58°23′47″ с.ш.; 56°22′9″ в.д.) – коренное сообщество, расположенное на равнинной местности, возраст деревьев которого составляет 135 лет. Состав древостоя 5Е2П2Лп1Б. Сомкнутость крон 0.6. Подрост состоит из Picea obovata , Abies sibirica , Tilia cordata Mill. и Betula pendula Roth . Кустарниковый ярус образован такими видами как Sorbus aucuparia , Lonicera xylosteum и Padus avium Mill. Проективное покрытие кустарничково-травяного яруса составляет 70–80%, где доминируют Oxalis acetosella , Dryopteris carthusiana , Stellaria nemorum и Gymnocarpium dryopteris L. Моховой покров состоит из зеленых мхов, растущих около стволов и на стволах деревьев, реже они встречаются на почве. Лишайники обнаружены на стволах деревьев, а также на валежнике, которого очень много. Почва песчаная, дерново-сильноподзолистая.
По периодам исследований в изучаемых биогеоценозах выявлялось от 39 (ельник кисличный) до 64 (ельник приручьевой) видов сосудистых растений. Их видовой состав со временем оставался относительно стабильным (ельник приручьевой: J I–II = 88, J II–III = 69, J I–III = 69; ельник кисличный: J I–II = 79, J II– III = 73, J I–III = 75). Видовой состав древесного яруса в исследуемых биогеоценозах с 1975 по 2012 гг. оставался неизменным.
Методы исследований
-
1) Учёт видового разнообразия, биомассы и числа плодовых тел грибов. В исследуемых биогеоценозах на пробных площадях размерами 50×20 м (1000 м2), заложенных ещё в 1975 г. Л.Г. Переведенцевой, в августе, один раз в декаду (три посещения с интервалом в 10 дней) проводился учет видового разнообразия, а также числа и биомассы плодовых тел агарикоидных грибов. Дополнительно, однократно, в сентябре изучался их видовой состав. Для учета числа и биомассы базидиом грибов проводился сбор всех плодовых тел на каждой учетной площади. Затем базидиомы каждого вида взвешивались и проводился пересчет на воздушно-сухую массу, учитывая, что воздушно-сухая масса составляет 10% от свежих ба-зидиом грибов.
-
2) Изучение микропризнаков и идентификация грибов. Идентификация грибов осуществлялась на световых микроскопах: ZEISS Axio Imager A2 и Olympus BX51M. В работе были использованы современные определители, монографии, атласы и справочные пособия зарубежных и отечественных авторов.
-
3) Мониторинг биоты эктомикоризных грибов. Для оценки сходства видового состава грибов по периодам наблюдений использовался коэффициент Жаккара ( J ×100) [Грейг-Смит, 1967; Леонтьев, 2008]:
J = —— X 100, a+i-c где J – индекс общности; c – число общих видов в двух сравниваемых биогеоценозах; a, b – число видов грибов в каждом из ценозов.
В качестве меры разнообразия сравниваемых биот исследуемых биогеоценозов использовался индекс Шеннона, описывающий два ее основных аспекта – богатство и сложность [Леонтьев, 2008]:
H = – ∑pilgpi , где H – индекс Шеннона; pi – относительное обилие каждого вида:
р i = n i / N , где n i – число базидиом (или биомасса базидиом) одного вида; N – общее число базидиом (или биомасса базидиом) в биогеоценозе.
Доминирующие виды грибов устанавливались по числу базидиом (шт./га) и по их воздушно-сухой биомассе (кг/га). Для выявления доминирующих видов грибов по биомассе и числу базидиом использовался индекс доминирования [Bochus, Babos, 1960]:
Д = а / b ×100, где Д – индекс доминирования; а – число базидиом (или биомасса) грибов данного вида; b – число бази-диом (или биомасса), собранных на всей учетной площади. К доминантам отнесены лишь те виды грибов, которые имеют индекс доминирования, равный 5 или более, что составляет 5 % или более от общего числа плодовых тел или их биомассы.
Принадлежность грибов к эколого-трофическим группам устанавливалась по шкале, предложенной А.Е. Коваленко [1980] с дополнениями некоторых авторов [Столярская, Коваленко, 1996; Морозова, 2001].
Влияние некоторых метеорологических показателей (по данным метеостанции г. Добрянки) на биоту эктомикоризных грибов изучалось с помощью корреляционного анализа. В качестве климатических показателей взяты: средняя месячная температура воздуха с мая по сентябрь (°С); сумма осадков (мм) по месяцам с мая по сентябрь; сумма осадков (мм) за май-сентябрь; сумма осадков (мм) и средняя температура воздуха (°С) по декадам августа. В качестве характеристик биоты эктомикоризных грибов взяты: 1) число видов за август-сентябрь; 2) число и биомасса базидиом грибов (за август в целом, а также по декадам августа). Математическая обработка осуществлялась при помощи программ Microsoft Office
Excel 2016 и StatSoft Statistica 10. Корреляционный анализ проводился с использованием коэффициента ранговой корреляции Спирмена ( r s ) [Трухачева, 2013].
Результаты и их обсуждение
Биота эктомикоризных грибов исследуемых еловых лесов довольно разнообразна. Далее приведен список видов, составленный по системе, принятой М. Moser [1983] с некоторыми дополнениями [Пере-веденцева, 1999]. Для каждого вида приводится латинское название. В скобках указаны синонимы грибов, соответствующие современной классификации [MycoBank Databases]. Римскими цифрами обозначены месяцы вегетационного периода. Хозяйственная значимость: съед. – съедобный, несъед. – несъедобный, яд. – ядовитый. Арабскими цифрами обозначен период выявления вида: 1 – в 1975–1977 гг., 2 – в 1994–1996 гг., 3 – в 2010–2012 гг. Место нахождения: ЕПР – ельник приручьевой, ЕК – ельник кисличный. Новые виды для Пермского края: ▲.
Аннотированный список видов эктомикоризных грибовAGARICALES Clements
Amanitaceae R. Heim ex Pousar: Amanita battarrae (Boud.) Bon – VII–IX, съед., ЕПР (2, 3), ЕК (2); A. crocea (Quél.) Singer – VIII–IX, съед., ЕПР (1–3), ЕК (1, 3); A. fulva Fr. – VII–IX, съед., ЕПР (1–3), ЕК (1–3); A. muscaria (L.) Lam. – VII–IX, яд., ЕПР (1), ЕК (1, 3); A. porphyria Alb. & Schwein. – VIII–IX, яд., ЕПР (1–3), ЕК (1, 3); A. regalis (Fr.) Michael. – VIII–IX, яд., ЕПР (1, 3); A. rubescens Pers. – VIII–IX, съед., ЕПР (1), ЕК (3); A. rubescens f. annulosulfurea (Gillet) J.E. Lange – VIII–IX, съед., ЕПР (1); A. vaginata (Bull.) Lam. – VIII–IX, съед., ЕПР (1, 3), ЕК (1, 2).
Cortinariaceae R. Heim ex Pouzar: Cortinarius acutus (Pers.) Fr. – VIII–IX, несъед., ЕК (2, 3); C. albidus Peck – VIII–IX, несъед., ЕК (3).; C. alboviolaceus (Pers.) Fr. – VII–IX, съед., ЕК (3); C. anomalus (Fr.) Fr. – VIII, несъед., ЕПР (2, 3), ЕК (1); C. argentatus (Pers.) Fr. – VIII–IX, несъед., ЕПР (1, 3), ЕК (3); C. argutus Fr. (= Phlegmacium argutum (Fr.) Niskanen & Liimat.) – VIII, несъед., ЕПР (2); C. armeniacus (Schaeff.) Fr. – VII–IX, несъед., ЕПР (1–3), ЕК (1, 3); C. armillatus (Fr.) Fr. – VII–VIII, съед., ЕПР (1, 3); C. atropusillus Favre – VIII–IX, несьед., ЕПР (2, 3) ▲; C. betuletorum M.M. Moser ex M.M. Moser – VIII–IX, несъед., ЕПР (1, 3), ЕК (1, 3); C. bolaris (Pers.) Fr. – VIII, несъед., ЕПР (3); C. brunneus (Pers.) Fr. – VIII–IX, несъед., ЕПР (1–3), ЕК (3); C. candelaris Fr. – VIII–IX, несъед., ЕПР (3). C. castaneus (Bull.) Fr. – VIII–IX, несъед., ЕПР (1–3); C. cinnamomeoluteus P.D. Orton – VIII–IX, несъед., ЕПР (3); C. cinnamomeus (L.) Fr. – VIII–IX, не-съед., ЕПР (1, 2); C. collinitus (Sowerby) Gray – VIII–IX, съед., ЕПР (1); C. decipiens Fr. – VIII–IX, несъед., ЕК (3); C. decoloratus (Fr.) Fr. – VIII, несъед., ЕПР (3), ЕК (3); C. duracinus Fr. – VIII, несъед., ЕПР (1); C. gentilis (Fr.) Fr. – VIII, несъед., ЕПР (3), ЕК (3); C. glandicolor (Fr.) Fr (= C. brunneus var. glandicolor (Fr.) H. Lindstr. & Melot) – VIII–IX, несъед., ЕПР (1–3); C. hemitrichus (Pers.) Fr. – VII–IX, несъед., ЕПР (1–3), ЕК (1–3); C. hoeftii (Weinm.) Fr. – VIII–IX, несъед., ЕПР (3); C. impennis Fr. – VIII, несъед., ЕК (2); C. incisus (Pers.) Fr. – VIII–IX, несъед., ЕПР (1); C. jubarinus Fr. – VIII, несъед., ЕК (3); C. leucopus (Bull.) Fr. – VIII–IX, несъед., ЕК (3), ▲; C. megasporus Singer – VIII–IX, несъед., ЕПР (3), ЕК (2, 3); C. nemorensis (Fr.) J.E. Lange – VIII, несъед., ЕПР (1, 2); C. obtusus (Fr.) Fr. – VIII–IX, несъед., ЕПР (2); C. paragaudis Fr. – VIII, несъед., ЕПР (1); C. pholideus (Lilj.) Fr. – VIII, съед., ЕПР (1); C. privignoides Rob. Henry – VIII, не-съед., ЕК (3); C. rigidus (Scop.) Fr. – VIII–IX, несъед., ЕПР (3), ЕК (2, 3); C. sanguineus (Wulfen) Fr. – VIII– IX, несъед., ЕПР (1–3); C. scandens Fr. – IX, несъед., ЕПР (3); C. semisanguineus (Fr.) Gillet – VIII–IX, не-съед., ЕПР (1–3); C. torvus (Fr.) Fr. – VIII, несъед., ЕПР (3), ▲; C. trivialis J.E. Lange – VIII–IX, несъед., ЕПР (1, 3); C. tubulipes J. Favre – VIII, несъед., ЕК (3), ▲; C. uraceus Fr. – VIII–IX, несъед., ЕПР (1, 3), ЕК (2, 3); C. violaceus (L.) Gray – VII–VIII, несъед., ЕК (1, 3); Hebeloma claviceps (Fr.) Quél. – VIII–IX, несъед., ЕПР (2); H. crustuliniforme (Bull.) Quél. – VII–IX, несъед., ЕПР (1, 2), ЕК (2, 3); H. hiemale Bres. – VIII–IX, несъед., ЕПР (2, 3), ЕК (3); H. pusillum J.E. Lange – VIII, несъед., ЕПР (3), ЕК (2); H. sacchariolens Quél. – VIII, несъед., ЕПР (2); H. sinapizans (Paulet) Gillet – VII–IX, несъед., ЕПР (1, 3), ЕК (1); Inocybe assimilata Britzelm. – VII–IX, яд., ЕПР (1–3); I. bongardii (Weinm.) Quél. (= Inosperma bongardii (Weinm.) Matheny & Esteve-Rav .) – VII–VIII, несъед., ЕПР (1, 3); I. flocculosa Sacc. – VIII–IX, несъед., ЕПР (1), ЕК (1–3); I. geophylla (Sowerby) P. Kumm. – VIII–IX, яд., ЕПР (1–3), ЕК (1, 3); I. glabripes Ricken – VIII, не-съед., ЕПР (3); I. hirtella Bres. – VIII, несъед., ЕК (3), ЕПР (3); I. jacobi Kühner (= Inocybe rufoalba Sacc. ) – VIII, несъед., ЕПР (3); I. lanuginosa (Bull.) P. Kumm. – VIII, яд., ЕК (1–3); I. muricellata Bres. – IX, несъед., ЕПР (3); I. paludinella (Peck) Sacc. – VIII–IX, несъед., ЕПР (3); I. pseudodestricta Stangl & J. Veselský – VIII–IX, несъед., ЕПР (3); I. rimosa (Bull.) P. Kumm. (= Pseudosperma rimosum (Bull.) Matheny & Esteve-Rav.) – VII–IX, яд., ЕПР (1, 2), ЕК (1–3); I. whitei (Berk. & Broome) Sacc. W.G. Sm. – VII–IX, яд., ЕПР (1); Rozites caperatus (Pers.) P. Karst. (= Cortinarius caperatus (Pers.) Fr.) – VIII–IX, съед., ЕПР (1, 2).
Entolomataceae Kotlaba & Pouzar: Clitopilus prunulus (Scop.) P. Kumm. VIII–IX, съед., ЕПР (3), ЕК (3); Entoloma rhodopolium (Fr.) P. Kumm. – VIII–IX, яд., ЕПР (1–3), ЕК (1, 2).
Hygrophoraceae Lotsy: Hygrophorus olivaceoalbus (Fr.) Fr. – VIII–IX, несъед., ЕПР (1–3); H. piceae Kühner – VIII–IX, несъед., ЕПР (2).
Tricholomataceae R. Heim ex Pousar: Clitocybe odora (Bull.) P. Kumm. – VII–IX, несъед., ЕПР (1, 2), ЕК (1–3); Collybia butyracea var. asema (Fr.) Cetto – VII–IX, съед., ЕПР (1–3), ЕК (1–3); C. butyracea var. butyracea (Bull.) Fr. – VII–IX, съед., ЕПР (1–3), ЕК (1–3); Laccaria bicolor (Maire) P.D. Orton – VI–VIII, съед., ЕПР (2, 3); L. laccata (Scop.) Cooke – VI–IX, съед., ЕПР (1–3), ЕК (1–3); L. proxima (Boud.) Pat. – VIII, съед., ЕПР (3); Lepista nebularis (Batsch) Harmaja (= Clitocybe nebularis (Batsch) P. Kumm.) –VIII–IX, съед., ЕПР (3); Tricholoma album (Schaeff.) P. Kumm. – VIII, несъед., ЕПР (3); T. fulvum (Fr.) Bigeard & H. Guill. – VIII, несъед., ЕК (3); T. inamoenum (Fr.) Gillet – VIII–IX, несъед., ЕПР (1, 2); T. lascivum (Fr.) Gillet – VIII, несъед., ЕПР (1–3), ЕК (1, 3).
BOLETALES J.-E. Gilbert
Boletaceae Chevall.: Boletus edulis Bull. – VII–IX, съед., ЕПР (1, 3), ЕК (1, 3); Chalciporu s piperatus (Bull.) Bataille – VIII–IX, несъед., ЕПР (1, 3), ЕК (1, 3); Leccinum aurantiacum (Bull.) Gray – VII–VIII, съед., ЕК (1); L. scabrum (Bull.) Gray –VI–IX, съед., ЕПР (1, 3), ЕК (1, 3); L. variicolor Watling – VII–IX, съед., ЕПР (1); L. versipelle (Fr. & Hök) Snell – VIII–IX, съед., ЕПР (3), ЕК (1, 2); Suillus bovinus (L.) Roussel – VIII–IX, съед., ЕПР (1); S. luteus (L.) Roussel – VI–VIII, съед., ЕПР (1); Tylopilus felleus (Bull.) P. Karst. – VIII, несъед., ЕПР (1–3), ЕК (1, 3); Xerocomus chrysenteron (Bull.) Quél. (= Xerocomellus chrysen-teron (Bull.) Šutara) – VII–IX, съед., ЕК (3); X. rubellus (Krombh.) Quél. (= Hortiboletus rubellus (Krombh.) Simonini, Vizzini & Gelardi) – VIII–IX, съед., ЕПР (1, 3), ЕК (1, 3); X. subtomentosus (L.) Quél. – VI–IX, съед., ЕПР (1–3), ЕК (1, 3).
Gomphidiaceae Maire ex Jülich: Chroogomphus rutilus (Schaeff.) O.K. Mill. – VI–IX, съед., ЕПР (1).
Paxillaceae Lotsy: Paxillus involutus (Batsch) Fr. – VIII–IX, яд., ЕПР (1–3), ЕК (1, 2).
RUSSULALES Kreisel ex Kirk et al.
Russulaceae Lotsy: Lactarius aurantiacus (Pers.) Gray – VIII, съед., ЕПР (3); L. camphoratus (Bull.) Fr. – VIII–IX, несъед., ЕПР (2, 3), ЕК (3); L. deterrimus Gröger – VIII, съед., ЕПР (1, 3), ЕК (1); L. flexuosus (Pers.) Gray (= L. zonarius (Bull.) Fr.) – VII–VIII, съед., ЕПР (1, 2), ЕК (1, 3); L. fuliginosus (Fr.) Fr. – VIII–IX, съед., ЕПР (3); L. glyciosmus (Fr.) Fr. – VII–IX, несъед., ЕПР (1–3), ЕК (2); L. lignyotus Fr. – VII–VIII, несъед., ЕПР (1–3); L. necator (Bull.) Pers. – VIII–IX, съед., ЕПР (1, 2), ЕК (1, 3); L. rufus (Scop.) Fr. – VII–IX, съед., ЕПР (1–3), ЕК (1); L. theiogalus (Bull.) Gray (= L. tabidus Fr. ) – VIII–IX, съед., ЕПР (1–3), ЕК (1–3); L. torminosus (Schaeff.) Pers. – VIII–IX, съед., ЕПР (1), ЕК (1); L. trivialis (Fr.) Fr. – VII–IX, съед., ЕПР (1–3), ЕК (1, 3); L. vietus (Fr.) Fr. – VIII–IX, съед., ЕПР (1–3), ЕК (1–3); Russula acrifolia Romagn. – VIII–IX, съед., ЕПР (2, 3); R. aeruginea Lindbl. ex Fr. – VII–VIII, съед., ЕПР (1–3), ЕК (1); R. atropurpurea (Krombh.) Britzelm. – VII–IX, несъед., ЕПР (1–3); R. betularum Hora – VIII–IX, несъед., ЕПР (1–3), ЕК (1–3); R. chamaeleontina (Lasch) Fr. (= Russula risigallina (Batsch) Sacc . ) – VII–IX, съед., ЕПР (1), ЕК (1–3); R. claroflava Grove – VII–IX, съед., ЕПР (1–3), ЕК (1–3); R. consobrina (Fr.) Fr. – VIII–IX, съед., ЕПР (1); R. decolorans (Fr.) Fr. – VII–VIII, съед., ЕПР (1–3), ЕК (1, 2); R. delica Fr. – VII–IX, съед., ЕПР (1–3), ЕК (1); R. elaeodes (Bres.) Bon (= R. clavipes Ve-len.) – VIII–IX, съед., ЕПР (1–3); R. emeticicolor (Jul. Schäff.) Singer – VII–VIII, съед., ЕПР (2, 3), ЕК (1, 2); R. foetens Pers. – VII–IX, съед., ЕПР (1–3), ЕК (3); R. fragilis Fr. – VIII–IX, несъед., ЕПР (1–3), ЕК (1–3); R. grisea Fr. – VII–VIII, съед., ЕПР (1, 3); R. integra (L.) Fr. – VIII–IX, съед., ЕПР (1–3); R. laurocerasi Melzer – VIII, съед., ЕПР (2); R. mustelina Fr. – VIII, съед., ЕПР (1, 2), ЕК (1); R. lutea (Huds.: Fr.) Gray – VII–VIII, съед., ЕПР (1); R. nauseosa (Pers.) Fr. – VIII–IX, съед., ЕПР (2, 3), ЕК (1, 2); R. ochroleuca Fr. – VII–IX, съед., ЕПР (3), ЕК (1, 2); R. puellaris Fr. – VII–IX, съед., ЕПР (1–3), ЕК (1–3); R. queletii Fr. – VIII–IX, несъед., ЕПР (1); R. vesca Fr. – VII–VIII, съед., ЕПР (1–3), ЕК (1); R. vinosa Lindblad – VIII, съед., ЕПР (1–3); R. xerampelina (Schaeff.) Fr. – VII–VIII, съед., ЕПР (1–3), ЕК (1, 3).
Анализ таксономической структуры эктомикоризных грибов
В исследуемых еловых лесах за все время наблюдений (1975–1977, 1994–1996, 2010–2012 гг.) выявлено 336 видов и внутривидовых таксонов агарикоидных базидиомицетов, 139 из которых являются ми-коризообразователями, входящими в состав 23 родов и 9 семейств (табл. 1).
Эктомикоризные грибы в исследуемых биогеоценозах являются преобладающей экологотрофической группой, к которой относится 37.4 (ельник кисличный) – 43.5% (ельник приручьевой) всех выявленных видов агарикоидных грибов. Преобладание микоризообразователей в составе микобиоты характерно для всей лесной зоны Голарктики. Видовой состав микоризообразователей в целом за все периоды наблюдений (1975–2012 гг.) ельника приручьевого достиг 124 видов (24 рода, 9 семейств), а ельника кисличного – 80 видов (18 родов, 7 семейств). По периодам число видов микоризообразователей варьируется, что вероятно связано с цикличностью образования плодовых тел эктомикоризных грибов. В третий период наблюдений впервые в еловых лесах обнаружено 30 видов микоризообразователей, из числа которых 4 вида оказались новыми для Пермского края: Cortinarius atropusillus, C. leucopus, C. torvus C. tubulipes.
Таблица 1
Таксономический состав эктомикоризных грибов еловых лесов [Taxonomic composition of ectomycorrhizal fungi of spruce forests]
|
Семейство (число родов/видов) |
Роды (число видов) |
Ельник приручьевой |
Ельник кисличный |
|
Amanitaceae (1/9) |
Amanita (9) |
9 |
7 |
|
Cortinariaceae (4/63) |
Cortinarius (43), Hebeloma (6), Inocybe (13), Rozites (1) |
52 |
30 |
|
Entolomataceae (2/2) |
Clitopilus (1), Entoloma (1) |
2 |
2 |
|
Tricholomataceae (5/11) |
Clitocybe (1), Collybia (2), Laccaria (3), Lepista (1), Tricholoma (4) |
10 |
6 |
|
Hygrophoraceae (1/2) |
Hygrophorus (2) |
2 |
– |
|
Boletaceae (6/12) |
Boletus (1), Chalciporus (1), Leccinum (4), Suillus (2), Tylopilus (1), Xerocomus (3) |
9 |
9 |
|
Gomphidiaceae (1/1) |
Chroogomphus (1) |
1 |
– |
|
Paxillaceae (1/1) |
Paxillus (1) |
1 |
1 |
|
Russulaceae (2/38) |
Lactarius (13), Russula (25) |
38 |
25 |
|
9 семейств |
23 рода (139 видов) |
124 |
80 |
Ведущими по числу видов в исследуемых биогеоценозах оказались сем.: Cortinariaceae (37.5–41.9%), Russulaceae (30.6–31.3%), Tricholomataceae (7.5–8.1%), Amanitaceae (7.3–8.8%) и Boletaceae (7.3–11.3%) к которым относится 95.2–96.4% всех выявленных микоризообразователей (рис. 1). Остальные семейства включали менее одного процента видов микоризных грибов. Наиболее крупными родами по числу видов эктомикоризных грибов являются: Cortinarius (9.8–11.6% видов), Russula (7.0–8.8%), Lactarius (4.6–4.7%) и Inocybe (2.3–4.2%), что характерно для лесных ценозов подзоны южной тайги.
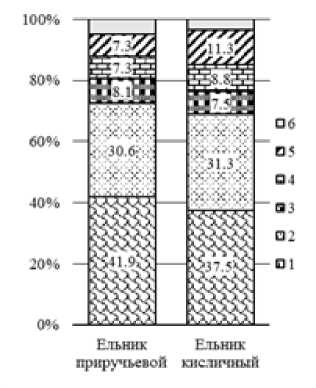
Рис. 1. Соотношение ведущих семейств эктомикоризных грибов в исследуемых биогеоценозах за все время наблюдений (в процентах от общего числа видов мико-ризообразователей за 1975–2012 гг. для каждого из ценозов):
1 - Cortinariaceae; 2 - Russulaceae ; 3 - Tricholomataceae; 4 - Amanitaceae ; 5 - Boletaceae ; 6 - остальные семейства
[The ratio of the leading families of ectomycorrhizal fungi in the investigate biogeocenoses for the entire time of observations (as a percentage of the total number of mycorrhizal species for 1975-2012 for each of the cenoses):
1 – Cortinariaceae; 2 – Russulaceae; 3 – Tricholomataceae; 4 – Amanita-ceae: 5 - Boletaceae: 6 - other families ]
Наличие или отсутствие базидиом эктомикоризных грибов может зависеть от различных факторов, поэтому в ходе ежегодных наблюдений выявляется только часть реально существующих в экосистеме видов грибов [Новожилов и др., 2016]. В связи с этим, все обнаруженные виды микоризообразователей в еловых лесах включались в общий список. В результате отмечено увеличение скрытого видового разнообразия микоризных грибов по годам наблюдений, хотя ежегодно выявляемое число видов сократилось во втором периоде наблюдений, а к концу третьего периода вновь увеличилось (рис. 2).
Под скрытым разнообразием понимается, что в зависимости от различных факторов грибы периодически формируют свои базидиомы, поэтому некоторые виды грибов не встречаются на учетной площади, хотя мицелий, вероятно, существует и при благоприятных условиях вновь формирует плодовые тела. Например, в 2010 г. в ельнике приручьевом базидиомы формировали всего лишь 24 вида микоризообра-зователей, а в целом к этому году в нем было обнаружено 107 видов. Кривые накопления выявляемого разнообразия эктомикоризных грибов в еловых лесах линейные, сохраняют восходящий тренд. В ельнике приручьевом по годам наблюдений наименьшее число эктомикоризных грибов зафиксировано в 2010 г. (24 вида), а наибольшее – в 2012 г. (75). В ельнике кисличном наименьшее число микоризообра-зователей зафиксировано в 2010 г. (10 видов), а наибольшее – в 1977 и 2012 гг. (46). Следовательно, появление базидиом эктомикоризных грибов в исследуемых биогеоценозах имеет волнообразный характер.
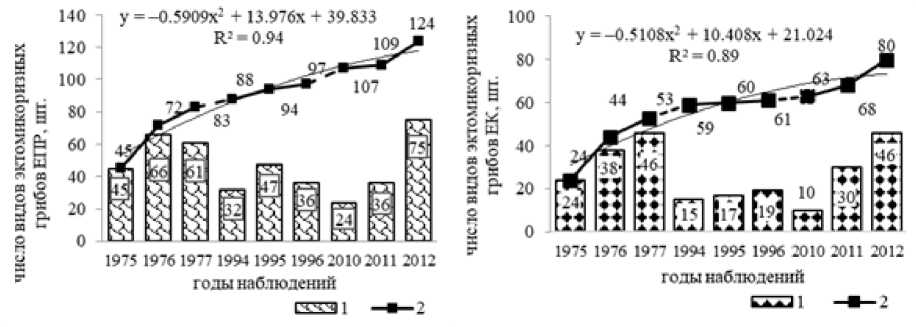
Рис. 2. Изменение видового разнообразия эктомикоризных грибов в исследуемых биогеоценозах по годам наблюдений:
ЕПР – ельник приручьевой; ЕК – ельник кисличный; 1 – число видов эктомикоризных грибов за год наблюдений; 2 – общее выявленное разнообразие эктомикоризных грибов
[Changes in the species diversity of ectomycorrhizal fungi in the investigate biogeocenoses by years of observation:
EПР – spruce forest at the brook; EK – sorrel spruce forest; 1 – the number of ectomycorrhizal fungi species per year of observations; 2 – the total revealed diversity of ectomycorrhizal fungi]
В исследованных еловых лесах за 9 лет наблюдений большинство видов (72.6–75.0%) обнаруживалось от 2 до 9 раз, с разными интервалами, что подтверждает гипотезу о наличии мицелиального континуума во времени. Некоторые виды эктомикоризных грибов (2–3%) были постоянными и встречались ежегодно. Как в ельнике приручьевом так и в кисличном, к ним относились Laccaria laccata и Lactarius theiogalus. Часть видов (25.0–27.4%) выявлена только один раз, в какой-либо сезон (рис. 3).
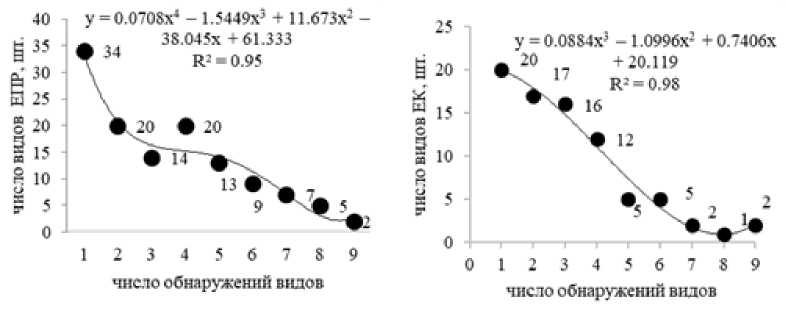
Рис. 3. Частота выявления видов эктомикоризных грибов в исследуемых биогеоценозах:
ЕПР – ельник приручьевой; ЕК – ельник кисличный
[Frequency of detection of ectomycorrhizal fungi species in the investigate biogeocenoses:
EПР – spruce forest at the brook; EK – sorrel spruce forest]
Вычислив индексы общности по видовому разнообразию эктомикоризных грибов в разные периоды, мы установили, насколько интенсивно произошло изменение биоты эктомикоризных грибов. В отличие от растений-хозяев, видовой состав которых практически остался прежним (за 1975–2012 гг.), биота эк-томикоризных грибов подвергалась значительным изменениям по периодам исследования (табл. 2).
Индексы общности (по Жаккару: J ×100) по грибам между периодами колебались от 32 до 52. Наиболее стабильным во времени оказался видовой состав микоризообразователей ельника приручьевого ( J I– II = 52, J II–III = 46, J I–III = 47), а менее стабильным – видовой состав грибов ельника кисличного ( J I–II = 39, J II–III = 32, J I–III = 46). Появление базидиом эктомикоризных грибов в климаксных сообществах не отличается стабильностью и зависит в большей степени не от растения-хозяина, а от внешних факторов. С течением времени общее (скрытое) биоразнообразие увеличивается. При сравнении эктомикоризных грибов между исследуемыми биогеоценозами выявлено сходство их видового состава. Коэффициенты Жаккара по микоризообразователям между ценозами по периодам наблюдений варьировали от 28 до 45 ( J I = 45, J II = 28, J III = 34).
Таблица 2
Число видов микоризообразователей и коэффициенты сходства по видовому составу по периодам наблюдений в исследуемых биогеоценозах
[Number of mycorrhizal fungi species and similarity coefficients by species composition by observation periods in the investigate biogeocenoses]
83 64 I 91 I 124 I 53 I 32 I 39 I 80
коэффициенты сходства по видовому составу грибов по Жаккару ( J ×100)
|
I–II |
II–III |
I–III |
I–II |
II–III |
I–III |
|
52 |
46 |
47 |
39 |
32 |
46 |
Примечание: I – 1975–1977 гг., II – 1994–1996 гг., III – 2010–2012 гг.
В целом, за все периоды наблюдений коэффициент сходства между исследуемыми ценозами был довольно высокий ( J = 47). Следовательно, по мере выявления новых видов эктомикоризных грибов увеличивается число их общих видов, т. е. происходит сближение биогеоценозов по скрытому видовому разнообразию грибов. Таким образом, видовое сходство микоризообразователей между исследуемыми сообществами составляло почти 50%, несмотря на разное число видов и разные экологические условия.
Доминирующие виды эктомикоризных грибов
Доминирующие виды грибов характеризуются наибольшими показателями по числу и биомассе ба-зидиом в биогеоценозе, поэтому данные характеристики биоты могут рассматриваться в качестве индикаторов экосистемной роли различных видов. Образование базидиом у грибов может быть связано с различными факторами: температурой, влажностью воздуха и почвы, метеорологическими условиями, и др. В благоприятных условиях грибы массово формируют базидиомы. Виды доминирующих грибов как по числу, так и по биомассе базидиом определялись по результатам сбора грибов, собранных в течение августа, в период массового «плодоношения» практически всех видов грибов. В каждом исследуемом биогеоценозе доминирующие виды грибов, как по числу, так и по биомассе базидиом, создают свой «микологический облик».
В еловых лесах число доминирующих видов эктомикоризных грибов как по числу базидиом, так и по биомассе базидиом значительно варьировало (табл. 3). Их доля в составе биоты всех микоризообразова-телей составляла более 50%.
В ельнике приручьевом за три периода наблюдений доминантами по числу базидиом являлись 9 видов грибов, а по биомассе базидиом – 10 видов. В ельнике кисличном за все время исследований выявлено 9 видов эктомикоризных грибов, доминирующих по числу базидиом и 12 видов – по биомассе бази-диом. В разные периоды доминировали: Boletus edulis, Collybia asema, Cortinarius brunneus, Laccaria lac-cata, Lactarius theiogalus, Leccinum scabrum, Paxillus involutus, Russula claroflava, R. ochroleuca, R. xerampelina и некоторые другие. Видовой состав доминантов к III периоду наблюдений существенно изменяется. В большей степени это касается доминантов по биомассе. Индексы общности, вычисленные для доминантов разных периодов исследования по числу базидиом, варьировали от 0 до 50, а по биомассе базидиом колебались от 0 до 33. Сравнивая индексы общности по доминантам с индексами, вычисленными по всему биоразнообразию эктомикоризных грибов, отметим, что индексы по доминантам чаще всего ниже, чем по общему видовому разнообразию микоризообразователей.
«Плодоношение» эктомикоризных грибов
Большое значение при изучении агарикоидных грибов имеют данные об их «урожайности», которые позволяют судить об объеме вторичной продукции биогеоценоза. По сезонам наблюдений, по данным метеостанции г. Добрянки, погодные условия отличались от средних многолетних данных повышенным фоном температур воздуха (относительно нормы) и довольно значительными колебаниями количества осадков. Самыми неблагоприятными для развития грибов следует считать засушливые 1975 г. (среднемесячная температура воздуха ниже нормы на 0.56–0.65 С, а количество осадков за июль–август ниже нормы на 26–31 мм) и 2010 г. (среднемесячная температура воздуха в июле–августе выше нормы на 2.0– 3.2 С, а количество осадков в июле ниже нормы на 64 мм, в августе все их количество выпало в конце третьей декады), а также 1994 г., отличающийся большим количеством осадков и пониженными температурами воздуха в июле-августе (среднемесячная температура воздуха ниже нормы на 1.0–2.6 С, а количество осадков за июль–август выше нормы на 27–83 мм).
По годам наблюдений также отмечаются значительные вариации «урожая» грибов, что связано с погодными условиями не только текущего года, но предыдущих лет. Например, резкое увеличение «урожайности» грибов по биомассе и количеству базидиом наблюдалось в 1976 г. после засушливого 1975 г. (рис. 4).
Таблица 3
Число доминантов, их процентные доли в составе биоты эктомикоризных грибов в исследуемых биогеоценозах и коэффициенты сходства по Жаккару ( J ×100)
[The number of dominants, their percentage shares in the composition of the biota of ectomycorrhizal fungi in the investigate biogeocenoses and the Jaccard similarity coefficients ( J ×100)]
|
cравниваемые периоды |
I–II |
25 |
17 |
50 |
33 |
|
II–III |
25 |
14 |
17 |
0 |
|
|
I–III |
13 |
0 |
0 |
8 |
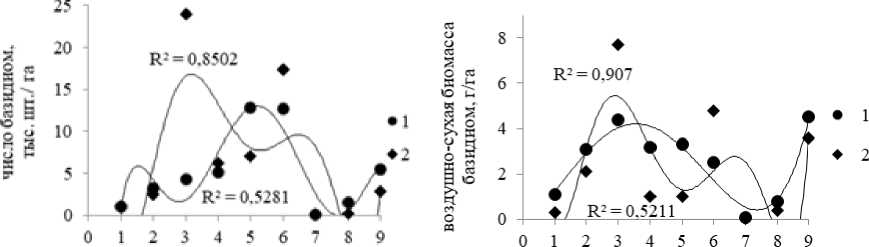
годы наблюдений годы наблюдений
Рис. 4. Динамика числа и биомассы базидиом эктомикоримзных грибов в исследуемых ценозах по годам наблюдений:
1 – ельник приручьевой, 2 – ельник кисличный
[Dynamics of the number and biomass of basidiomes of ectomycorrhizal fungi in the studied cenoses by years of observations:
1 – spruce forest at the brook; 2 – sorrel spruce forest]
Максимальные показатели «урожайности» микоризообразователей по числу базидиом в ельнике при-ручьевом выявлены в 1977, а в ельнике кисличном – в 1996 г. Наибольшая биомасса базидиом эктомико-ризных грибов в исследуемых еловых лесах зафиксирована в 1977 г. Самыми «низкоурожайными» были засушливые 1975, 2010, и 2011 гг. Таким образом, по сезонам наблюдений в исследуемых биогеоценозах число и биомасса базидиом эктомикоризных грибов значительно варьируются, что связано с погодными условиями текущего года.
По периодам (сравнение во времени) в исследуемых биогеоценозах число и биомасса базидиом экто-микоризных грибов также варьируются. Во всех биогеоценозах по периодам наблюдений, по числу бази-диом максимальные «урожаи» выявлены во II период исследований (31 тыс. шт./га в каждом из ельни- ков). В этот период зафиксировано массовое развитие грибов как с крупными плодовыми телами (Amanita muscaria, Boletus edulis, Leccinum scabrum, Tylopilus felleus и др.), так и с мелкими (Laccaria laccata и др.). По биомассе базидиом максимальные показатели «урожайности» зафиксированы в I (ельник кисличный – 10.1 кг/га) и II периоды (ельник приручьевой – 9.0 кг/га) наблюдений. В целом за все периоды наблюдений наибольшее число базидиом зафиксировано для ельника кисличного (62 тыс. шт./га), а наибольшая биомасса базидиом – для ельника приручьевого (22.8 кг/га). Наименьшие показатели числа и биомассы базидиом в исследуемых биогеоценозах отмечены в III период, что, вероятно, связано с высокими температурами воздуха и низким уровнем осадков в течение периода наблюдений (табл. 4).
Таблица 4
Число и воздушно-сухая биомасса базидиом грибов, значения индексов Шеннона по периодам наблюдений в каждом из биогеоценозов
[Number and air-dry biomass of fungal basidiomes, Shannon indexes values on observations periods in each of the biogeocenoses]
|
Ельник приручьевой |
Ельник кисличный |
||||||
|
периоды |
за все периоды |
периоды |
за все периоды |
||||
|
I |
II |
III |
I |
II |
III |
||
|
число базидиом, шт./га |
|||||||
|
9 |
30 |
1 7 1 |
46 1 |
28 1 |
31 1 |
3 1 |
62 |
|
воздушно-сухая биомасса, кг/га |
|||||||
|
8.5 |
9.0 |
5.3 |
22.8 |
10.1 |
6.7 |
3.9 |
20.7 |
|
значения индексов Шеннона ( H ) по числу |
базидиом |
||||||
|
1.52 |
0.96 |
1.17 |
1.27 |
0.79 |
0.77 |
1.35 |
0.92 |
|
значения индексов Шеннона ( H ) по биомассе базидиом |
|||||||
|
1.34 |
1.29 |
1.18 |
1.54 |
1.23 |
0.91 |
1.16 |
1.36 |
Примечание: I – 1975–1977 гг., II – 1994–1996 гг., III – 2010–2012 гг.
Значения индексов Шеннона по числу и биомассе базидиом микоризообразователей (за август) по периодам наблюдений варьировались. Наибольшие их значения как по числу, так и по биомассе базидиом грибов в ельнике приручьевом отмечены в I период. Однако в ельнике кисличном по числу базидиом максимальные их значения отмечены в III период, а по биомассе базидиом – в I период наблюдений. В целом за три периода наблюдений наибольшие значения индексов Шеннона как по числу, так и по биомассе базидиом отмечены в ельнике приручьевом. Следовательно, за все периоды наблюдений биота эктомикоризных грибов ельника приручьевого являлась более разнообразной и ее составляющие наиболее выровнены.
Таким образом, в ельнике приручьевом, по сравнению с ельником кисличным, для большего числа видов эктомикоризных грибов были более благоприятные и разнообразные условия для развития их ба-зидиом, а в ельнике кисличном – лишь для некоторых видов, которые массово появлялись в каждый из периодов исследований.
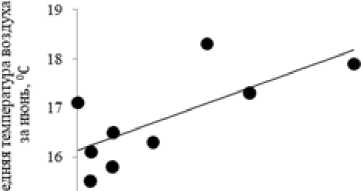
В изучаемом биогеоценозе по годам наблюдений выявлено немного статистически значимых корреляций характеристик микобиоты эктомикоризных грибов с погодными условиями текущего года. Так, с ростом средней месячной температуры воздуха в июне прослеживается увеличение суммарной биомассы базидиом микоризных грибов в августе ( r s = 0.67; p <0.05) (рис. 5). Средняя температура воздуха и сумма осадков за август не влияет на количество и биомассу базидиом эктомикоризных грибов в разные годы наблюдений.
" is ■
О 2000 4000 6000 8000
воздушно-сухая биомасса базидием микоризных грибов, кг та
Рис. 5. Диаграмма рассеяния средней температуры воздуха за июнь и воздушно-сухой биомассы базидиом эктомикоризных грибов, кг/га ( r s = 0.67; p <0.05)
[Scatter diagram of average air temperature for June and air-dry biomass of basidiomes of ectomycorrhizal fungi, kg/ha ( r s = 0.67; p <0.05)]
В результате анализа данных по декадам наблюдений (август), т. е. во время учета базидиом, установлено, что метеорологические показатели по декадам августа оказывали слабое влияние на биоту эк-томикоризных грибов во всех исследуемых еловых лесах. По декадам исследований были отмечены корреляции только средней силы ( r s от 0.30 до 0.70; или r s от -0.30 до -0.70; при p <0.05), а сильных статистически значимых корреляций по декадам наблюдений не было выявлено во всех исследуемых ценозах (табл. 5).
Таблица 5
Коэффициенты корреляции Спирмена по декадам августа в исследуемых ценозах
[Spearman correlation coefficients on the decades of August in the investigate cenoses]
|
Характеристики биоты эктомикоризных грибов по декадам августа |
к! ей .__ Н « а й а П 5 Я й и 2 а о и н о |
к! « f Д 1—| ® а a S U 2 м й о О ь д о |
к! те н й Ь И ей U 2 д й О о У идо |
о Й h п 9 |
!^а о W Н « й |
£ М ей о |
|
ельник приручьевой |
||||||
|
Число базидиом, шт./га |
–0.34 |
–0.41 |
0.07 |
0.55 |
0.26 |
–0.58 |
|
Биомасса базидиом, г/га |
0.03 |
–0.33 |
–0.07 |
0.33 |
0.43 |
–0.20 |
|
ельник кисличный |
||||||
|
Число базидиом, шт./га |
–0.29 |
–0.60 |
–0.23 |
0.58 |
0.10 |
–0.38 |
|
Биомасса базидиом, г/га |
–0.07 |
–0.44 |
–0.25 |
0.21 |
0.09 |
–0.39 |
Примечание: жирным цветом отмечены коэффициенты с уровнем статистической значимости p < 0.05.
Анализируя пищевую ценность эктомикоризных грибов в еловых лесах, мы выявили, что 69 видов являлись съедобными, они обычно имели крупные плодовые тела. Несъедобными были 60 видов, имеющих небольшие размеры базидиом, либо обладающих неприятным запахом и вкусом. Ядовитых грибов обнаружено 10 видов.
Заключение
В еловых лесах за все время исследований (1975–1977, 1994–1996, 2010–2012 гг.) выявлено 139 видов микоризообразователей. Видовой состав эктомикоризных грибов ельника приручьевого достиг 124 видов (24 рода, 9 семейств), а ельника кисличного – 80 видов (18 родов, 7 семейств). Лидирующими по числу видов эктомикоризных грибов за все время исследований были сем. Cortinariaceae , Russulaceae , Tricholomataceae , Amanitaceae и Boletaceae , что характерно для бореальной зоны. Наиболее крупными родами по числу видов грибов являются: Cortinarius , Russula , Lactarius и Inocybe , что характерно для лесных ценозов подзоны южной тайги. Новыми для еловых лесов в третий период наблюдений являлись 30 видов эктомикоризных грибов, из них 4 вида оказались новыми для Пермского края.
Видовой состав эктомикоризных грибов с течением времени меняется в большей степени ( J = 32–52), чем видовой состав сосудистых растений ( J = 69–88). Во всех исследуемых сообществах в течение трех периодов наблюдений большинство видов (от 72.6 до 75.0%) встречалось от 2 до 9 раз с разными интервалами, что подтверждает гипотезу о наличии мицелиального континуума во времени. Некоторые виды эктомикоризных грибов (2–3%) были постоянными и встречались ежегодно. Следовательно, для выявления всех видов грибов необходимы длительные и стационарные наблюдения. По мере обнаружения новых видов эктомикоризных грибов между биогеоценозами происходит сближение их видового состава ( J = 47), что свидетельствует о наличии мицелиального континуума в пространстве.
В еловых лесах по периодам наблюдений число доминирующих видов эктомикоризных грибов варьировало как по числу базидиом (от 2 до 7 видов), так и по биомассе базидиом (от 3 до 7 видов),. Их доля в составе биоты всех микоризообразователей за все время наблюдений всегда составляла более 50% как по числу, так и по биомассе базидиом. Видовой состав доминантов по периодам наблюдений как по числу ( J = 0–50), так и по биомассе базидиом ( J = 0–33) подвергался большей трансформации, в отличие от всей биоты эктомикоризных грибов.
Продуктивность эктомикоризных агарикоидных грибов в исследуемых ценозах различается по годам наблюдений и зависит от изменения погодных условий текущего года. В целом за все периоды наблюдений наибольшее число базидиом зафиксировано для ельника кисличного, а наибольшая биомасса бази-диом – для ельника приручьевого. Для биоты эктомикоризных грибов как по числу, так и по биомассе базидиом за все периоды наблюдений более благоприятным являлся ельник приручьевой, т. к. биота ми-коризообразователей ельника приручьевого была более разнообразна и ее составляющие наиболее вы- ровнены (H = 1.27; 1.54). Установлено, что повышение средней месячной температуры воздуха в июне благоприятствует «плодоношению» эктомикоризных грибов в августе (rs = 0.67; p <0.05). Средняя месячная температура воздуха и сумма осадков за август по годам наблюдений в рамках нашего исследования не влияет на «продуктивность» микоризообразователей данных ценозов.
Среди 139 видов эктомикоризных грибов, выявленных в еловых лесах 69 являются съедобными, 60 – несъедобны, и 10 – ядовиты.
