Морфофизиологическая характеристика нагульного лосося (Salmo salar L.) Онежского озера с использованием статистических методов
 Онежского озера с использованием статистических методов")
Автор: Дзюбук Ирина Михайловна, Клюкина Елена Александровна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 2 (123), 2012 года.
Бесплатный доступ
В статье представлены результаты исследований морфофизиологических характеристик нагульного лосося Онежского озера (возраст, масса, длина, высота, упитанность, пол, индексы органов) с использованием статистического анализа. Установлены различия между самками и самцами по индексу желудка, между особями из разных условий воспроизводства по индексам сердца и желудка. Выявлены закономерности изменений индексов внутренних органов, не зависящих от пола и условий воспроизводства.
Онежское озеро, пресноводный лосось, морфофизиологические показатели, компонентный анализ
Короткий адрес: https://sciup.org/14750088
IDR: 14750088 | УДК: 591.1
Morpho-physiological characteristics of Onego lake-feeding salmon (Salmo salar L.) obtained by use of statistical methods
Research results of the morpho-physiological characteristics of the lake-feeding salmon inhabiting Onego Lake (age, weight, length, height, fatness, gender, internal organ indexes) are presented in the article. The results were obtained with the use of the statistical analysis. Gender distinctions in salmon population were defined with the help of the stomach index. Distinctive differences in species coming from various habitats were determined by the use of the heart and stomach indexes. Consistent patterns of changes in the inner organs, independent upon gender and conditions of reproduction, were revealed.
Текст научной статьи Морфофизиологическая характеристика нагульного лосося (Salmo salar L.) Онежского озера с использованием статистических методов
Исследование лосося Онежского озера представляет определенный научный интерес, так как это ценный промысловый вид, численность которого неуклонно сокращалась [6]. Вследствие этого пресноводный лосось с 2001 года был занесен в Красную книгу Карелии. Меры по охране и воспроизводству этого вида привели к увеличению его численности [11].
С 1999 года сотрудниками Лаборатории экологических проблем Севера ПетрГУ проводится мониторинг юго-западного района Онежского озера, включающий и ихтиологические исследования. В ходе работ дается оценка популяций различных видов рыб, в том числе и пресноводного лосося. Результаты исследования нагульного лосося Онежского озера были представлены в работах Л. П. Рыжкова, И. М. Крупень с соавторами [12], [16], [17], [18]. Морфофизиологические показатели нагульного лосося оценивались с применением элементарных статистических приемов: вычисление средней, ее ошибки и критерия Стьюдента.
Цель настоящей работы – провести анализ данных по результатам морфофизиологического исследования нагульного лосося Онежского озера, используя современные методы математической статистики.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для статистического анализа послужили данные морфофизиологического исследования (размер, масса, индексы внутренних органов) лосося, выловленного в юго-западном районе Онежского озера (Шелтозеро - Брусно,
1999 год). Сбор и обработка ихтиологического материала осуществлялась по общепринятым методикам [14], [15], [19], [22].
Для анализа были использованы 50 особей (3+…7+), из них 30 самок и 20 самцов; 25 особей воспроизводились в заводских условиях («заводские»), 25 – в естественных («дикие»).
При математико-статистической обработке результатов исследования применялся дескриптивный (описательный) анализ, методы сравнения двух независимых выборок и компонентный анализ [1], [2]. Оценка достоверности различий между средними арифметическими осуществлялась по критерию Стьюдента (t-тест), а между дисперсиями - по критерию Фишера (F-тест) (при нормальном распределении признака) для уровня значимости б = 0,05 [4], [5], [13].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В ходе анализа мы учитывали два альтернативных признака: самка - самец и естественное воспроизводство - заводское воспроизводство.
Итак, 30 проб из 50 содержат самок (значение 1), 20 - не содержат (значение 0). То есть выборка состоит из 30 единиц и 20 нулей. Доля вариантов со значением 1 (доля самок) составляет: 30
p = 50 = 0,6 (60 %), тогда доля самцов q = 0,4 (40 %). Отметим, что наиболее точные границы доверительного интервала для альтернативного признака дает ц-преобразование Фишера [9]. Итак, найдем границы доли самок лосося p = 0,8 при уровне значимости б = 0,05: φ = 1,772 – 2,008 × лев.
Х 50= = 1,4889, фправ. = 1,772 + 2,008" В = 2,0551, pлев.(1,4889) = 45,9 %, pправ.(2,0551) = 73,3 %. То есть доля самок в генеральной совокупности (популяции лососей) составляет минимум 45,9 %, максимум 73,3 %.
Самки и самцы лосося были в возрасте от 3+ до 7+ лет. Наибольшее количество как самок, так и самцов было в возрасте 5+ или 2,3+ (то есть общий возраст – 5+, при этом рыбы 2-го года жили в реке и 3-го года в озере после ската). Масса (m), длина (D) и высота тела (h) лосося были в пределах 692,5-4325,0 г, 47,0-78,3 и 41,4-70,0 см соответственно.
Для альтернативного признака «условия воспроизводства» 25 проб из 50 содержат особей, воспроизведенных в естественных условиях (значение 1), и 25 - не содержат (значение 0). Значит, выборка состоит из 25 единиц и 25 нулей. Доля варианта со значением 1 составляет: p = ля = ця (50 %). Найдем с помощью ц-преоб-яц разования Фишера границы доверительного интервала при уровне значимости б = 0,05 для доли «диких» особей (p = 0,5: φлев.= 1,571 – 2,008 × х -1= = 1,288, Фправ. = 1,571 + 2,008 • -1= = 1,8541, 50 50
pлев. (1,288) = 36 %, pправ. (1,8541) = 64 %. Таким об-раз . ом, доля естествен . но воспроизведенных лососей в генеральной совокупности составляет минимум 36 %, максимум 64 %.
Возраст «заводских» особей был в пределах 3+…7+ лет, «диких» - 3+…5+. Наибольшее количество особей было возраста 5+ (2,3+).
При сравнении полученных результатов исследования нагульного лосося с данными Т. Н Александровой с соавторами, М. Б. Зборовской, Ю. А. Смирнова [3], [7], [20] можно заметить, что за многолетний период не произошло значимых изменений по следующим показателям: возраст, размер, масса, количественный состав в различных размерных группах, хотя нагульное стадо в настоящее время сформировано из «диких» и «заводских» рыб.
В дополнение имеющихся данных [18] был проведен анализ различий между самками и самцами нагульного лосося не только по массе и размерам тела, но и по индексам внутренних органов.
Анализ различий между самками и самцами нагульного лосося
Масса и размер тела. И для самцов, и для самок выборочные значения стандартизированных асимметрии и эксцесса массы тела, а также размеров тела попадают в нормальный диапазон (табл. 1). При этом доверительные интервалы на уровне надежности 95 % для выборочных средних для самок и самцов соответственно равны: 1) [1827,51; 2848,49] и [1852,65; 2736,68] по массе (m); 2) [59,68; 69,67] и [60,87; 68,07] по длине тела (D); 3) [56,88; 66,77] и [57,84; 64,94] по длине тела
Таблица 1
Общая статистическая характеристика для массы, размеров и индексов органов самок и самцов лосося (3+…7+), лето 1999 года
|
Параметр |
Статистика |
||||||
|
Среднее |
Дисперсия |
Коэффициент вариации, % |
Предел варьирования |
Стандартизир. асимметрия |
Стандартизир. эксцесс |
||
|
Масса (m) |
♂ |
2338,0 |
1,18975E6 |
46,65 |
400–5000 |
0,27 |
0,85 |
|
♀ |
2294,67 |
1,40122E6 |
51,59 |
400–5100 |
1,971 |
0,23 |
|
|
Длина тела (D) |
♂ |
64,68 |
114,06 |
16,51 |
43,5–81 |
-1,68 |
0,08 |
|
♀ |
64,47 |
92,88 |
14,95 |
41,5–82,5 |
-0,31 |
0,14 |
|
|
Длина тела без хвостового плавника (d) |
♂ |
61,83 |
111,80 |
17,10 |
40,5–77 |
-1,73 |
0,12 |
|
♀ |
61,39 |
90,41 |
15,49 |
39,0–79,5 |
-0,21 |
0,04 |
|
|
Высота тела (h) |
♂ |
58,20 |
97,85 |
17 |
38–72 |
-1,76 |
0,08 |
|
♀ |
57,73 |
75,94 |
15,09 |
37–75 |
-0,34 |
0,01 |
|
|
Индекс сердца |
♂ |
1,81 |
0,09 |
16,57 |
1,34–2,43 |
1,11 |
-0,42 |
|
♀ |
1,95 |
0,38 |
31,79 |
1,37–4,75 |
7,67 |
16,6 |
|
|
Индекс печени |
♂ |
12,63 |
16,81 |
32,46 |
1,32–20,58 |
-1,12 |
2,13 |
|
♀ |
13,63 |
17,32 |
30,52 |
8,77–30,25 |
5,11 |
8,94 |
|
|
Индекс селезенки |
♂ |
9,54 |
8,29 |
30,17 |
5,8–16,48 |
1,92 |
0,53 |
|
♀ |
9,08 |
3,52 |
20,68 |
5,86–13,51 |
1,21 |
0,52 |
|
|
Индекс жабр |
♂ |
40,65 |
165,17 |
31,62 |
4,64–71,51 |
-0,86 |
3,21 |
|
♀ |
40,78 |
138,2 |
28,82 |
4,88–70,62 |
-0,48 |
3,21 |
|
|
Индекс желудка |
♂ |
2,44 |
0,83 |
37,36 |
1,02–4,29 |
0,52 |
-0,73 |
|
♀ |
2,44 |
7,65 |
113,6 |
0,97–16,42 |
10,61 |
27,31 |
|
|
Индекс кишечника |
♂ |
27,77 |
34,07 |
21,02 |
20,54–48,7 |
4,73 |
8,04 |
|
♀ |
27,54 |
32,87 |
20,82 |
11,72–43,75 |
0,53 |
2,92 |
|
Примечание. ♂ – самцы, ♀ – самки.
без хвостового плавника (d); 4) [53,57; 62,82] и [54,48; 60,99] по высоте тела (h).
Проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения для массы (m) и всех размеров тела (D, d, h) самок и самцов: для альтернативной гипотезы вычисленные значения вероятности P (0,896; 0,943; 0,879; 0,862) больше 0,05, альтернативная гипотеза о неравенстве средних отвергается. Доверительные интервалы на уровне надежности 95 % для стандартных отклонений для самок и самцов соответственно равны: 1) [829,51; 1593,13] и [942,73; 1591,31] (m); 2) [8,12; 15,6] и [7,68; 12,96] (D); 3) [8,04; 15,44] и [7,57; 12,78] (d); 4) [7,52; 14,45] и [6,94; 11,71] (h).
Проверка равенства дисперсий с помощью критерия Фишера показала, что гипотеза о равенстве дисперсий выборок по массе и размерам тела (D, d, h) не отвергается, так как P (0,722; 0,604; 0,592; 0,526) больше 0,05.
Таким образом, по размерно-весовым показателям различий между самками и самцами нагульного лосося не было выявлено.
Для анализа был проведен расчет индексов органов (в %о) лосося, которые широко используют в качестве морфофизиологических индикаторов состояния водной среды и организмов [15], [19], [21].
Индекс сердца. Значения стандартизированных асимметрии и эксцесса для самок лосося значительно превышают правую границу нормального диапазона от -2 до 2, что указывает на существенные отклонения от нормального распределения (табл. 1). У одной самки (3+) индекс сердца (4,8 %о) больше, чем у других самок выборки, для которых максимальный предел варьирования равен 2,8 %о. Исключив из рассмотрения данную особь, получили значения стандартизированных асимметрии и эксцесса для самок: 1,95 и 0,98 соответственно, которые принадлежат нормальному диапазону. Проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения по индексу сердца самок и самцов (P = 0,65 > 0,05); гипотеза о равенстве дисперсий выборок по критерию Фишера также подтверждается (P = 0,72 > 0,05).
Индекс жабр. Для обеих выборок значения стандартизированных эксцесса и асимметрии не попадают в нормальный диапазон, что указывает на существенные отклонения этих выборок от нормального (табл. 1). У 2 самок (2,1+ и 2,3+) индекс жабр (43,8 и 11,7 %о) не входит в пределы варьирования данного показателя у особей этой выборки - 19,6-36,3 %о. У одного из самцов (2,3+) индекс жабр (48,7 %о) больше максимального предела варьирования индекса у самцов этой группы - 34,8 %о. Исключив из рассмотрения выбросы, получили значения стандартизированных асимметрии и эксцесса для самок и самцов соответственно: 1,21 и 0,12; 0,97 и 1,05, принадлежащие нормальному диапазону. Проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения по индексу жабр самок и самцов (P = 0,45 > 0,05); критерий Фишера показал, что гипотеза о равенстве дисперсий выборок также подтверждается (P = 0,32 > 0,05).
Индекс печени. Выборочные значения стандартизированного эксцесса для обоих полов, а также выборочное значение стандартизированной асимметрии самок находятся за пределами нормального диапазона (табл. 1). Это указывает на существенные отклонения исследуемых выборок от нормального распределения. У 2 самок (2,2+ и 3,2+) индекс печени (30,3 и 19,9 %о) был больше максимального предела варьирования этого показателя (16,9 %о) у самок данной выборки. У 2 самцов (2,2+ и 2,3+) индекс печени (20,6 и 1,3%о) находится за пределами диапазона (8,8-18,4 %о) индекса органа у самцов изучаемой группы. Исключив из рассмотрения выбросы, получили соответственно значения стандартизированных асимметрии и эксцесса для индекса печени у самок (0,26 и -1,3) и у самцов (0,85 и 0,03), принадлежащие нормальному диапазону. Проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения индекса печени самок и самцов (P = 0,997 > 0,05); критерий Фишера подтвердил гипотезу о равенстве дисперсий выборок (P = 0,646 > 0,05).
Индекс селезенки. Значения стандартизированных асимметрии и эксцесса по индексам селезенки для самок находятся за пределами нормального диапазона, что указывает на существенные отклонения данной выборки от нормального распределения (табл. 1). Для самок максимальный предел варьирования индекса равен 3,8 %о, однако у одной самки (2,2+) он больше (16,4 %о). Исключив этот выброс, получили соответственно значения стандартизированных асимметрии и эксцесса (1,74 и -0,66), которые принадлежат нормальному диапазону. С помощью t-теста подтверждена гипотеза о равенстве среднего значения по индексу селезенки самок и самцов (P = 0,059 > 0,05), а критерий Фишера показал, что гипотеза о равенстве дисперсий выборок также подтверждается (P = 0,65 > 0,05).
Индекс желудка. Для самок и самцов выборочные значения стандартизированных асимметрии и эксцесса для индекса желудка находятся в пределах нормального диапазона (табл. 1). Критерий Фишера показал, что гипотеза о равенстве дисперсий выборок отвергается (P = 0,037 < 0,05). Таким образом, по индексу желудка самки имеют существенные отличия от самцов.
Индекс кишечника. Для обоих полов выборочные значения стандартизированного эксцесса для индекса кишечника не принадлежат нормальному диапазону от -2 до 2 (табл. 1). У 2 самок (3,2+ и 2,4+) индекс кишечника (70,6 и 4,9 %о) находится за пределами диапазона 21,8-61,9 %о, свойственного самкам исследованной выборки. Также у 2 самцов (2,2+ и 2,3+) индекс не входит в пределы от 29,3 до 51,92 ‰, которые отмечены у самцов исследуемой возрастной группы. Исключив выбросы, получили значения стандартизированного эксцесса для индекса кишечника самок и самцов: 0,01 и -1,14, принадлежащие нормальному диапазону. С помощью t-теста подтвердили гипотезу о равенстве среднего значения по индексу органа (P = 0,98 > 0,05), а критерий Фишера показал, что гипотеза о равенстве дисперсий выборок также подтверждается (P = 0,59 > 0,05).
Анализ различий между «дикими» и «заводскими» особями
Масса и размер тела. Для «диких» и «заводских» рыб (3+…5+) выборочные значения стандартизированных асимметрии и эксцесса массы, а также размеров тела попадают в нормальный диапазон (табл. 2). При этом доверительные интервалы на уровне надежности 95 % для выборочных средних «диких» и «заводских» особей соответственно равны: 1) [1635,23; 2543,23] и [1524,56; 2193,84] по массе; 2) [58,15; 67,46] и [57,64; 65,48] по длине тела (D); 3) [55,49; 64,55] и [54,53; 62,36] по длине тела без хвостового плавника (d); 4) [52,15; 60,58] и [52,15; 60,58] по высоте тела (h).
Проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения для массы тела и всех размеров (D, d, h) этих выборок: для альтернативной гипотезы о неравенстве средних вычисленные значения вероятности P (0,40; 0,69; 0,61; 0,66) больше 0,05, то есть альтернативная гипотеза отвергается. Доверительные интервалы на уровне надежности 95 % для стандартных отклонений для «диких» и «заводских» рыб соответственно равны: 1) [538,73; 1240,17] и [633,02; 1127,81] по массе; 2) [5,52; 12,72] и [7,42; 13,22] по длине тела (D); 3) [5,38; 12,38] и [7,40; 13,19] по длине тела без хвостового плавника (d); 4) [5,0; 11,51] и [6,92; 12,33] по высоте тела (h). Критерий Фишера показал, что гипотеза о равенстве дисперсий выборок по массе и размерам тела (D, d, h) не отвергается, так как значения вероятности P (0,81; 0,45; 0,4; 0,39) больше 0,05.
Также были исследованы отличия между естественно воспроизведенными и воспроизведенными на заводе особями (3+…5+) по индексам органов (табл. 2).
Индекс сердца. Для индекса сердца вычисленные значения стандартизированных асимметрии и эксцесса для «диких» и «заводских» особей в несколько раз превышают правую границу нормального диапазона от -2 до 2, что ука-
Таблица 2
Общая статистическая характеристика для массы, размеров и индексов органов лосося разных условий воспроизводства (3+…5+), лето 1999 года
|
Параметр |
Статистики |
||||||
|
Среднее |
Дисперсия |
Коэффициент вариации, % |
Предел варьирования |
Стандартизир. асимметрия |
Стандартизир. эксцесс |
||
|
Масса (m) |
Е |
2089,23 |
564424 |
35,96 |
800–3300 |
-0,32 |
-0,65 |
|
З |
1859,2 |
657233 |
43,60 |
400–3200 |
-0,66 |
-0,68 |
|
|
Длина тела (D) |
Е |
62,81 |
59,36 |
12,27 |
47,5–72 |
-0,81 |
-0,26 |
|
З |
61,56 |
90,34 |
15,44 |
41,5–76 |
-1,82 |
-0,06 |
|
|
Длина тела без хвост. плавника (d) |
Е |
60,02 |
56,22 |
12,49 |
45,5–69,5 |
-0,67 |
-0,41 |
|
З |
58,45 |
89,92 |
16,22 |
39,0–74 |
-1,61 |
-0,14 |
|
|
Высота тела (h) |
Е |
56,37 |
48,64 |
12,37 |
43,5–65,5 |
-0,60 |
-0,56 |
|
З |
55,12 |
78,55 |
16,08 |
37,0–69,0 |
-1,66 |
-0,18 |
|
|
Индекс сердца |
Е |
1,77 |
0,13 |
20,11 |
1,43–2,75 |
3,03 |
3,48 |
|
З |
1,99 |
0,39 |
31,43 |
1,34–4,75 |
7,78 |
17,41 |
|
|
Индекс печени |
Е |
14,91 |
30,0 |
36,74 |
9,59–30,25 |
2,96 |
3,64 |
|
З |
13,18 |
13,81 |
28,2 |
1,32–20,58 |
-1,95 |
3,61 |
|
|
Индекс селезенки |
Е |
2,21 |
0,99 |
45,0 |
1,07–3,65 |
0,54 |
-1,22 |
|
З |
2,88 |
8,54 |
101,5 |
1,21–16,42 |
9,11 |
21,74 |
|
|
Индекс жабр |
Е |
27,84 |
20,34 |
16,2 |
19,59–36, 25 |
0,4 |
0,07 |
|
З |
28,03 |
50,88 |
25,45 |
11,72–48,7 |
1,97 |
3,33 |
|
|
Индекс гонад |
Е |
2,73 |
4,8 |
80,36 |
0,33–6,24 |
0,64 |
-1,13 |
|
З |
2,01 |
2,76 |
82,8 |
0,25–5,25 |
1,07 |
-1,31 |
|
|
Индекс желудка |
Е |
9,94 |
3,8 |
19,61 |
7,35–13.51 |
1,24 |
-0,11 |
|
З |
8,92 |
4,39 |
23,5 |
5,8–15,08 |
2,26 |
2,14 |
|
|
Индекс кишечника |
Е |
38,62 |
218,59 |
38,29 |
4,64–70,62 |
-0,22 |
2,25 |
|
З |
45,1 |
92,2 |
21,29 |
29,81–71,51 |
1,5 |
1,14 |
|
Примечание. Е – лосось естественного воспроизводства, З – лосось заводского воспроизводства.
зывает на существенные отклонения от нормального распределения. Заметим, что среди естественно воспроизведенных особей у одной (2,2+) индекс сердца (2,8 ‰) больше, чем у рыб изучаемой группы, для которых максимальный предел варьирования равен 2,18. Среди «заводских» особей у одной (2,1+) индекс сердца (4,8 ‰) также больше, чем у других особей изучаемой группы, для которых максимальный предел варьирования равен 2,43. Исключив из рассмотрения данные особи, получили следующие значения стандартизированных асимметрии и эксцесса соответственно: 1,83 и 1,61 для «диких», 0,42 и 0,28 для «заводских» особей, которые принадлежат нормальному диапазону. Критерий Фишера показал, что гипотеза о равенстве дисперсий выборок подтверждается на данном уровне значимости (P = 0,497 > 0,05), однако проверка с помощью t-теста отвергла гипотезу о равенстве среднего значения по индексу сердца воспроизведенных в разных условиях особей (P = 0,029 < 0,05). Следовательно, между особями естественного и заводского воспроизводства существуют достоверные различия по индексу сердца.
Индекс жабр. Значение стандартизированного эксцесса для индекса жабр воспроизведенных на заводе особей не принадлежит нормальному диапазону от -2 до 2 (табл. 2). Заметим, что для выборки «заводских» особей (3+…5+) есть три выброса (11,7, 48,7, 43,8 ‰), индексы жабр которых отличаются от относительного веса органа остальных особей, у которых индекс жабр варьирует от 20,5 до 34,8 ‰. Исключив из рассмотрения данные выбросы, получили следующие значения стандартизированных асимметрии и эксцесса для «заводских» рыб соответственно: 0,24 и -0,37, которые принадлежат нормальному диапазону. Критерий Фишера показал, что гипотеза о равенстве дисперсий выборок подтверждается (P = 0,317 > 0,05), проверка с помощью t-теста также подтвердила гипотезу о равенстве среднего значения по индексу жабр «диких» и «заводских» особей (P = 0,597 > 0,05).
Индекс селезенки. Для индекса селезенки значения стандартизированных асимметрии и эксцесса для «заводских» рыб в несколько раз превышают правую границу нормального диапазона от -2 до 2, что указывает на существенные отклонения от нормального распределения. У одной особи (2,2+) индекс селезенки (16,4 ‰) больше, чем у рыб этой группы, для которых максимальный предел варьирования равен 3,8. Исключив выброс, получили значения стандартизированных асимметрии и эксцесса для воспроизведенных на заводе особей (0,52 и -1,2 соответственно), принадлежащие нормальному диапазону. При этом критерий Фишера показал, что гипотеза о равенстве дисперсий выборок подтверждается (P = 0,301 > 0,05), а проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения по индексу селезенки особей, воспроизведенных в разных условиях (P = 0,724 > 0,05).
Индекс печени. Значения стандартизированных асимметрии и эксцесса для «диких» особей по индексу печени, а также стандартизированного эксцесса для «заводских» рыб превышают правую границу нормального диапазона, что указывает на существенные отклонения от нормального распределения. У одной особи естественного воспроизводства (2,2+) индекс печени (30,3 ‰) больше, чем у остальных «диких» особей, для которых максимальный предел варьирования равен 19,9 ‰. У одной особи заводского воспроизводства (2,3+) индекс печени (1,3 ‰) меньше, чем у «заводских» особей изучаемой группы, для которых минимальный предел варьирования равен 9,3 ‰. Исключив из рассмотрения выбросы, получили значения стандартизированных асимметрии и эксцесса соответственно: 0,87 и -0,06 для «диких» особей; 1,28 и 0,17 для «заводских» особей, принадлежащие нормальному диапазону. Критерий Фишера показал, что гипотеза о равенстве дисперсий выборок подтверждается (P = 0,697 > 0,05), а проверка с помощью t-теста подтвердила гипотезу о равенстве среднего значения по индексу печени воспроизведенных в разных условиях особей (P = 0,962 > 0,05).
Индекс желудка. Значения стандартизированных асимметрии и эксцесса для индекса желудка «заводских» рыб немного превышают правую границу нормального диапазона. У одной особи заводского воспроизводства (2,1+) индекс желудка (15,1 ‰) больше, чем у «заводских» рыб, для которых максимальный предел варьирования равен 12,9 ‰. Исключив из рассмотрения выброс, получили значения стандартизированных асимметрии и эксцесса «заводских» особей соответственно: 0,83 и 0,44, принадлежащие нормальному диапазону. Критерий Фишера показал, что гипотеза о равенстве дисперсий выборок подтверждается (P = 0,54 > 0,05), однако проверка с помощью t-теста отвергла гипотезу о равенстве среднего значения по индексу желудка особей разных условий воспроизводства (P = 0,045 < 0,05).
Индекс кишечника. Для индекса кишечника вычисленное значение стандартизированного эксцесса для «диких» особей превышает правую границу нормального диапазона. У двух особей (3,2+ и 2,3+) индекс кишечника (70,6 и 4,6 ‰) выходит за границы диапазона 30,7–51,3 ‰, характерного для остальных рыб этой группы. Исключив из рассмотрения выбросы, получили значения стандартизированных асимметрии и эксцесса, которые принадлежат нормальному диапазону: 0,72 и -0,35 соответственно. Критерий Фишера показал, что гипотеза о ра- венстве дисперсий выборок подтверждается (P = 0,229 > 0,05), а проверка с помощью t-теста отвергла гипотезу о равенстве среднего значения по индексу кишечника (P = 0,057 > 0,05). Следовательно, между особями разных условий воспроизводства существуют достоверные различия по индексу кишечника.
Оценка степени различия с помощью компонентного анализа
Степень различия между самками и самцами оценивалась по массе тела (m), длине тела (D), длине тела без хвостового плавника (d), высоте тела (h), индексам сердца (m1), печени (m2), селезенки (m3), жабр (m4), желудка (m5) и кишечника (m6) (табл. 3).
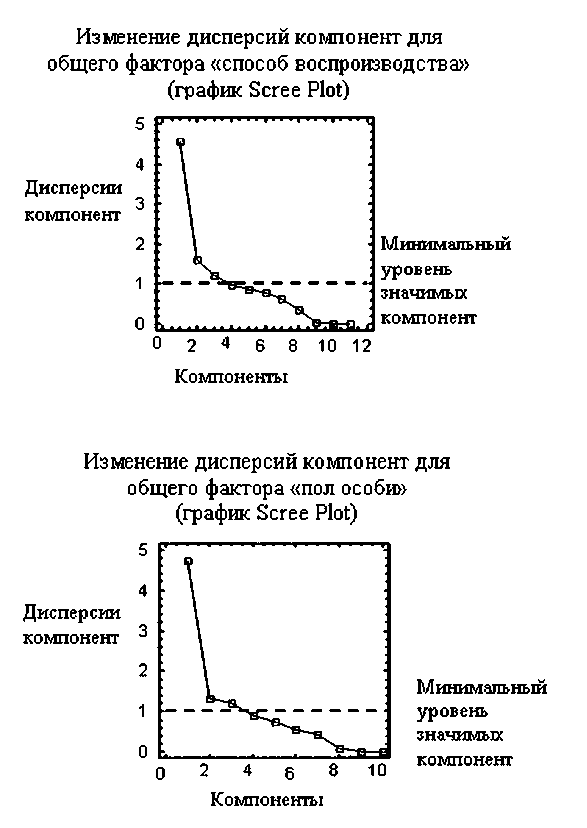
Заметим, что первая компонента учитывает большую долю исходной изменчивости (ее дисперсия равна SГ2К1 = 4,72 из 10, относительный вес составляет 47,2 %). Информативность второй компоненты выражается дисперсией SГ2К2 = 1,321 и относительным весом, равным 13,21 %. Информативность третьей компоненты выражается дисперсией SГФКш = йсФФ и относительным весом, равным 12,23 %. Дисперсии остальных компонент меньше единицы, что указывает на их низкую информативность, поскольку по умолчанию минимальное значение дисперсии главной компоненты (Minimum Eigenvalue) принимается равным 1. График Scree Plot отражает изменение дисперсий компонент (сплошная линия) и минимальный уровень компонент (пунктир) (см. рисунок).
Значения факторных нагрузок для первых трех главных компонент рассматриваемых морфологических признаков представлены в табл. 4.
Результаты расчетов компонентного анализа
Таблица 3
|
Номер компоненты |
Значение дисперсий главных компонент (Eigenvalue) |
Относительный вес главных компонент (Percent of variance) |
Суммативный вес главных компонент (Cumulative percentage) |
Значение дисперсий главных компонент (Eigenvalue) |
Относительный вес главных компонент (Percent of variance) |
Суммативный вес главных компонент (Cumulative percentage) |
|
Для общего фактора «пол особи» |
Для общего фактора «способ воспроизводства» |
|||||
|
1 |
4,720 |
47,20 |
47,20 |
4,589 |
41,72 |
41,72 |
|
2 |
1,321 |
13,21 |
60,41 |
1,608 |
14,62 |
56,34 |
|
3 |
1,223 |
12,23 |
72,64 |
1,213 |
11,03 |
67,37 |
|
4 |
0,912 |
9,12 |
81,75 |
0,961 |
8,73 |
76,10 |
|
5 |
0,756 |
7,56 |
89,32 |
0,839 |
7,62 |
83,72 |
|
6 |
0,560 |
5,60 |
94,92 |
0,772 |
7,02 |
90,74 |
|
7 |
0,431 |
4,31 |
99,23 |
0,603 |
5,48 |
96,22 |
|
8 |
0,068 |
0,68 |
99,91 |
0,357 |
3,25 |
99,47 |
|
9 |
0,008 |
0,08 |
99,99 |
0,046 |
0,42 |
99,89 |
|
10 |
0,001 |
0,01 |
100,00 |
0,012 |
0,11 |
99,99 |
|
11 |
– |
– |
– |
0,001 |
0,01 |
100,00 |
Таблица значений факторных нагрузок в главных компонентах
Таблица 4
|
Признаки |
Компонента |
|||||
|
123 |
123 |
|||||
|
Для общего фактора «пол особи» |
Для общего фактора «способ воспроизводства» |
|||||
|
Масса тела (m) (г) |
0,441 |
-0,099 |
0,188 |
0,439 |
-0,111 |
0,059 |
|
Длина тела (D) (см) |
0,439 |
-0,103 |
0,211 |
0,452 |
-0,060 |
0,143 |
|
Длина тела без хвостового плавника (d) (см) |
0,437 |
-0,115 |
0,213 |
0,451 |
-0,063 |
0,166 |
|
Высота тела (h) |
0,432 |
-0,095 |
0,140 |
0,450 |
-0,067 |
0,166 |
|
Индекс сердца (m1) (мг) |
-0,248 |
0,259 |
0,508 |
-0,278 |
0,033 |
0,524 |
|
Индекс печени (m2) (мг) |
-0,222 |
-0,535 |
0,211 |
-0,170 |
-0,525 |
-0,005 |
|
Индекс селезенки (m3) (мг) |
-0,107 |
-0,172 |
-0,352 |
-0,073 |
-0,006 |
-0,427 |
|
Индекс жабр (m4) (мг) |
-0,177 |
0,387 |
0,561 |
-0,176 |
0,238 |
0,646 |
|
Индекс желудка (m5) (мг) |
-0,173 |
-0,533 |
0,253 |
-0,156 |
-0,525 |
0,020 |
|
Индекс кишечника (m6) (мг) |
-0,226 |
-0,374 |
0,223 |
-0,132 |
-0,451 |
0,177 |
|
Индекс гонад (m7) (мг) |
– |
– |
– |
0,113 |
-0,403 |
0,123 |
Изменение дисперсий компонент
Пример уравнения расчета значений первой компоненты:
Component 1 = 0,432 ⋅ m + 0,441 ⋅ D + 0,439 ⋅ d + + 0,437 ⋅ h – 0,248 ⋅ m 1 – 0,222 ⋅ m 2 – 0,107 ⋅ m 3 – – 0,177 ⋅ m 4 – 0,173 ⋅ m 5 – 0,226 ⋅ m 6.
Все факторные нагрузки первых четырех признаков положительны и больше 0,3, остальные - отрицательны и по модулю меньше 0,3, то есть их влияние наименее значимо. Первую компоненту можно назвать «Увеличение размеров лосося при увеличении его массы». Для второй компоненты 8 из 10 факторных нагрузок отрицательны, причем для индексов печени, желудка и кишечника их величина по модулю меньше 0,3; индекс жабр и индекс сердца имеют положительную факторную нагрузку, но только для индекса жабр ее величина больше 0,3. Поэтому вторую компоненту назовем «Увеличение индекса жабр при уменьшении индексов печени, желудка и кишечника». Третью компоненту можно назвать «Уменьшение индекса селезенки при увеличении индексов жабр и сердца», поскольку она объясняется в основном за счет переменных «индекс селезенки», «индекс жабр» и «индекс сердца», их факторные нагрузки соответственно равны -0,352, 0,561 и 0,508.
Отметим, что для всех трех главных компонент и для самцов, и для самок численные ха- рактеристики принимают как положительные, так и отрицательные значения, то есть для этих компонент знак численной характеристики особи не зависит от пола.
Степень различия между особями разных условий воспроизводства оценивалась по m, D, d, h, m1, m2, m3, m4, m5, m6, m7 (индекс гонад) (табл. 3).
Как и в первой задаче, первые три компоненты накопили в себе основную содержательную информацию об отличиях особей по способу воспроизводства, заменяя собой в данном случае 11 исходных признаков. Дисперсия первой компоненты S 2 = 4 589 из 11, относительный ГК 1
вес составляет 41,72 %. Информативность второй компоненты выражается дисперсией S Г 2 К 2 = 1,608 и относительным весом, равным 14,62 %. Информативность третьей компоненты выражается дисперсией S Г 2 К 3 = 1,213 и относительным весом, равным 11,03 %. График Scree Plot отражает изменение дисперсий компонент (сплошная линия) и минимальный уровень компонент (пунктир) (см. рисунок).
Значения факторных нагрузок для первых трех главных компонент рассматриваемых морфологических признаков представлены в табл. 4. Так, уравнение расчета значений первой компоненты следующее:
Component 1 = 0,439 ⋅ m + 0,452 ⋅ D + 0,451 ⋅ d + + 0,45 ⋅ h – 0,278 ⋅ m 1 – 0,17 ⋅ m 2 – 0,073 ⋅ m 3 – – 0,176 ⋅ m 4 – 0,156 ⋅ m 5 – 0,132 ⋅ m 6 + 0,113 ⋅ m 7.
Все факторные нагрузки первых четырех признаков положительны и больше 0,3. Все остальные факторные нагрузки по модулю меньше 0,3, то есть их влияние наименее значимо, причем все они, кроме последней, отрицательны. Таким образом, первую компоненту, как и в первой задаче, назовем «Увеличение размеров лосося при увеличении его массы». Для второй компоненты 9 из 11 факторных нагрузок отрицательны (положительны только факторные нагрузки индексов сердца и жабр), причем только для индексов печени, желудка, кишечника и гонад их величина по модулю больше 0,3. Поэтому вторую компоненту можно назвать «Уменьшение индекса гонад при уменьшении индексов печени, желудка и кишечника». Третью компоненту, как и в первой задаче, можно назвать «Уменьшение индекса селезенки при увеличении индексов жабр и сердца», поскольку она объясняется в основном за счет переменных «индекс селезенки», «индекс жабр» и «индекс сердца», их факторные нагрузки соответственно равны -0,427, 0,646 и 0,524.
Отметим, что численные характеристики особей, выраженные в трех главных компонентах и для естественно воспроизведенных лососей, и для особей, воспроизведенных на заводе, принимают как положительные, так и отрицательные значения. Таким образом, для этих компонент знак численной характеристики особи не зависит от способа воспроизводства.
ЗАКЛЮЧЕНИЕ
Анализ морфофизиологических характеристик нагульного лосося юго-западного района Онежского озера (Шелтозеро - Брусно) с использованием современных методов математической статистики позволил существенно дополнить имеющиеся материалы [12], [16], [17], [18] новыми статистически достоверными (P ≥ 0,95) результатами.
Установлено, что самцы и самки лосося (3+…7+) достоверно различаются (P ≥ 0,95) по индексу желудка. Для остальных морфофизиологических признаков (масса, длина тела, длина тела без хвостового плавника, высота тела, индексы сердца, печени, селезенки, жабр, кишечника), исключая особи с патологией, существенных различий между самками и самцами не обнаружено (P < 0,05; при нормальном характере распределения выборок). Также для нагульного лосося были выявлены следующие закономерности, которые не зависят от пола рыбы: увеличение размеров лосося при увеличении массы; увеличение индекса жабр при уменьшении индексов печени, желудка и кишечника; уменьшение индекса селезенки при увеличении индексов жабр и сердца.
Показано, что естественно воспроизведенные и воспроизведенные на заводе особи (3+…5+) достоверно различаются (P ≥ 0,95) по индексам сердца и желудка. Для остальных морфофизиологических признаков (масса, длина тела, длина тела без хвостового плавника, высота тела, индексы печени, селезенки, жабр, кишечника, гонад), исключая особи с патологией, существенных различий не обнаружено (P < 0,05; при нормальном характере распределения выборок). Также для лосося (3+…5+) были выявлены следующие закономерности, которые не зависят от способа воспроизводства лосося: увеличение размеров лосося при увеличении массы; уменьшение индекса гонад при уменьшении индексов печени, желудка и кишечника; уменьшение индекса селезенки при увеличении индексов жабр и сердца.
Список литературы Морфофизиологическая характеристика нагульного лосося (Salmo salar L.) Онежского озера с использованием статистических методов
- Айвазян С. А., Енюков И. С., Мешалкин Л. Д. Основы моделирования и первичная обработка данных. М.: Финансы и статистика, 1983. 471 с.
- Айвазян С. А., Енюков И. С., Мешалкин Л. Д. Прикладная статистика: Классификация и снижение размерности. М.: Финансы и статистика, 1989. 607 с.
- Александрова Т. Н., Гуляева А. М., Кудерский Л. А. О нагульном лососе Онежского озера//Труды Карельского отдела ГосНИОРХ. Т. 5. Вып. 1. Петрозаводск, 1968. С. 274-286.
- Боровиков В. П., Боровиков И. Д. Statistica. Статистический анализ и обработка данных в среде Windows. М.: Информационно-издательский дом «Филин», 1997. 608 с.
- Гублер Е. В., Генкин А. А. Применение непараметрических критериев статистики в медико-биологических исследованиях. Л.: Медицина, 1973. 142 с.
- Жаков Л. А., Меншуткин В. В. Практические занятия по ихтиологии. Ярославль: Ярославский госуниверситет, 1982. 112 с.
- Зборовская М. Б. Материалы по промыслу и биологии лосося Онежского озера//Труды КНИРС. Т. 1. Л., 1935. С. 257-281.
- Ивантер Д. Э., Рыжков Л. П. Рыбы. Петрозаводск: Изд-во ПетрГУ, 2004. 176 с.
- Коросов А. В. Специальные методы биометрии: Учеб. пособие. Петрозаводск: Изд-во ПетрГУ, 2007. 364 с.
- Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных: Метод. пособие. Петрозаводск: Изд-во ПетрГУ, 2010. 84 с.
- Красная книга Республики Карелия. Петрозаводск: Карелия, 2007. С. 229-230.
- Крупень И. М., Рыжков Л. П. Морфофизиологическая индикация нагульного лосося Онежского озера//Современные проблемы физиологии и биохимии водных организмов: Материалы междунар. конф. 6-9 сентября 2004. Петрозаводск, 2005. С. 86-91.
- Малета Ю. С., Тарасов В. В. Непараметрические методы статистического анализа в биологии и медицине. М.: Изд-во МГУ, 1982. 178 с.
- Правдин И. Ф. Руководство по изучению рыб. М.: Пищ. пром-сть, 1966. 376 с.
- Применение метода морфофизиологических индикаторов для оценки качественного состава рыб: Методические указания. Петрозаводск: Изд-во ПетрГУ, 1997. 20 с.
- Рыжков Л. П., Крупень И. М. Значение искусственного воспроизводства в формировании структуры нагульной части популяции онежского лосося//Проблемы воспроизводства, кормления и борьбы с болезнями рыб при выращивании в искусственных условиях: Материалы науч. конф. 14-18 октября 2002. Петрозаводск, 2002. С. 102-107.
- Рыжков Л. П., Крупень И. М. Морфофизиологические особенности нагульного лосося Онежского озера//Атлантический лосось: Биология, охрана и воспроизводство. Петрозаводск, 2003. С. 158-164.
- Рыжков Л. П., Крупень И. М. Пресноводный лосось Онежского озера. Петрозаводск: Изд-во ПетрГУ, 2004. 152 с.
- Смирнов В. С., Божко А. М., Рыжков Л. П., Добринская Л. А. Применение метода морфофизиологических индикаторов в экологии рыб//Труды СевНИОРХ. Т. 7. Петрозаводск, 1972. 190 с.
- Смирнов Ю. А. Лосось Онежского озера. Л.: Наука, 1971. 143 с.
- Строганов Н. С. Экологическая физиология рыб. М.: Изд-во АН СССР, 1962. 444 с.
- Чугунова Н. И. Методика изучения возраста и роста рыб. М.: Советская наука, 1952. 115 с.
