Морфофизиологические особенности форгутных инфузорий лося как система адаптаций к эндобионтному обитанию

Автор: Корчагина Т.А.
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Экология
Статья в выпуске: 1, 2010 года.
Бесплатный доступ
В статье приведены оригинальные данные по морфофизиологическим и адаптационным особенностям эндобионтных инфузорий, обитающих в желудке лося (Alces alces Linnaeus, 1758).
Короткий адрес: https://sciup.org/14719470
IDR: 14719470
Текст научной статьи Морфофизиологические особенности форгутных инфузорий лося как система адаптаций к эндобионтному обитанию
Инфузории пищеварительного тракта травоядных млекопитающих относятся к царству Protozoa Coldfuss, 1818, типу Ciliophora Doflein, 1901, классу Ciliata Petry, 1852. Они известны с середины XIX в. по работе Gruby и Delafond (1843). Все последующие исследования эндобионтных простейших во второй половине XIX в. содержали сведения об инфузорной фауне преимущественно сельскохозяйственных животных [5].
Первая половина XX в. была ознаменована важнейшими открытиями в изучении фауны эндобионтных инфузорий жвачных. В этот период были опубликованы работы отечественных протозоологов Г. Н. Тассовского, В. А. Догеля, А. А. Стрелкова и ряда зарубежных ученых, посвященные изучению эндобионтных инфузорий как сельскохозяйственных, так и диких травоядных млекопитающих [1 — 3].
В настоящее время в прото зоологической литературе периодически появляются описания новых видов и форм эндобионтных инфузорий, в основном из пищеварительного тракта копытных. Несколько работ посвящены новым инфузориям из рубца сайги Saiga tatarica Linnaeus, 1766 и лошадиных, антилопы «болотный козел» Kobus leche kafuensis Knottnerus-Meyer, 1907, буйвола Bubalus bubalis Linnaeus, 1758, американского лося Alces americana Linnaeus, 1758. Особый интерес представляет цикл работ, посвященных инфузориям пищеварительного тракта сумчатых, в значительной степени изменивших взгляды на филогению и систематику эндобионтных инфузорий. Описаны новые виды из кенгуру Macropus rufus Desmarest, 1822, валлаби Lagostrophus fasciatus Gray, 1821 и других австралийских сумчатых [4].
Отделы желудка лося A. alces и родственных ему других жвачных являются местом обитания различных по морфологии и систематическому положению видов ин- фузорий. Но до сих пор фауна и жизненные циклы большинства известных видов эндобионтных инфузорий, обитающих в желудке жвачных, и их влияние на организм хозяина исследованы неполно. Таким образом, изучение фауны, особенностей биологии и экологии инфузорий диких жвачных в разных регионах мира позволяет расширить наши представления об этих своеобразных и хорошо адаптированных к организму хозяина простейших.
Целью исследования стало изучение особенности морфологии инфузорий желудка лося как системы адаптации к эндо-бионтному обитанию.
Для достижения цели были поставлены следующие задачи:
-
1) изучить видовой состав инфузорий желудка лося, составить дифференциальные диагнозы;
-
2) выяснить особенности морфологии у ресничных инфузорий из разных отделов желудка лося с использованием разнообразных стандартных цитохимических методик;
-
3) при изучении некоторых физиологических процессов включить форгутных инфузорий лося в соответствующие «группы питания».
Материал был собран в 2002 — 2006 гг. из желудка лосей с территорий Челябинской, Тюменской и Омской областей. Из разных отделов желудка 12 особей лося было собрано 600 проб (по 50 из каждой особи). Дополнительно брали пробы из отделов кишечника каждой особи: слепой и прямой кишок и толстого кишечника. Пробы фиксировались 4 % формалином. Материалы были получены на двух марках световых микроскопов БиоМед-2 и МБИ-6 с фазовоконтрастным устройством. Рисунки инфузорий сделаны при помощи рисовального аппарата РА-4 (ок. Юх, об. 40х), а также по контурам фотографий. Определение видов проведено по таблицам офриосколецид [1;2].
Инфузории желудка лося относятся к
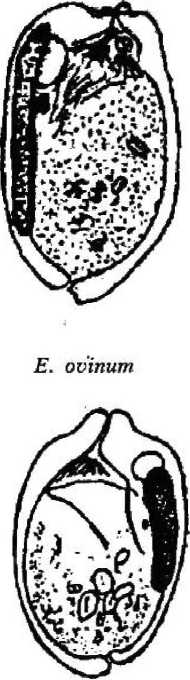
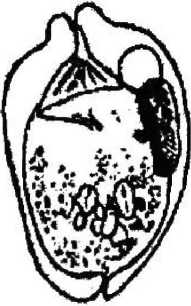
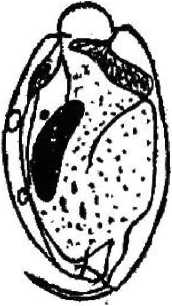
форгутным, т. е. населяющим только желудок. В пробах, взятых из различных отделов кишечника животного, они не обнаружены. Наиболее многочисленным по количеству видов является рубец, где нами обнаружено 15 видов инфузорий: Entodinium bursa Dogiel, 1925; Entodinium dices, Dehority, 1974; Entodinium nanellum Dogiel, 1923; Entodinium ovinum Dogiel, 1927; Entodinium simplex Dogiel, 1927; Entodinium caudatum Stein, 1859;
Entodinium simulans-dubardi Dogiel, 1925;
Entodinium exiguum Dogiel, 1925;
Entodinium longinucleatum Dogiel, 1925;
Diplodinium monacanthum Dogiel, 1927;
Diplodinium rangiferi Dogiel, 1929;
Diptodinium bubalidis ssp. bubalidis
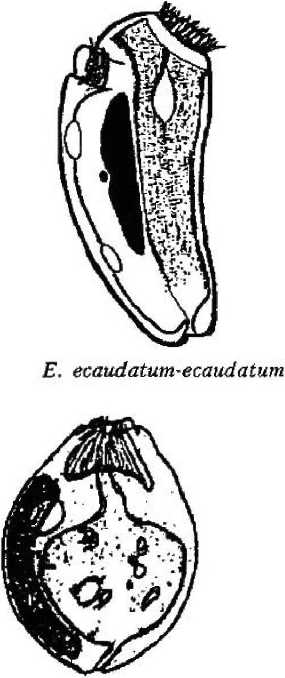
Latteur, 1970; Diplodinium bubalidis ssp. consors Dogiel, 1927; Epidinium ecaudatum-ecaudatum (Fior) Sharp, 1914; Dasytricha ruminantium (Schuberg, 1888) Dogiel, 1925 (рис. 1).
E. nanellum
E. longinucleatum
E. simplex
D. rangiferi
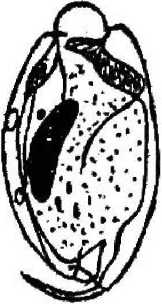
D. bubalidis ssp. consors
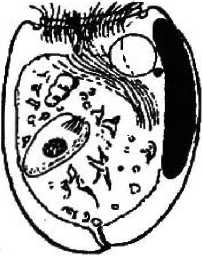
E. bursa
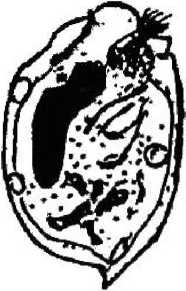
D. monocanthum
Рисунок 1
Видовое разнообразие эндобионтных инфузорий желудка лося
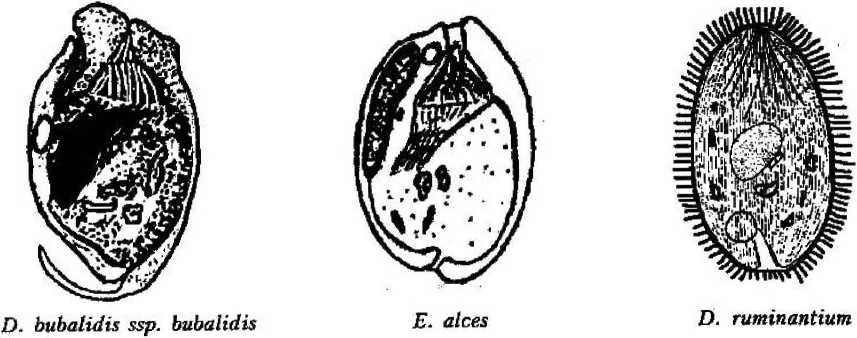
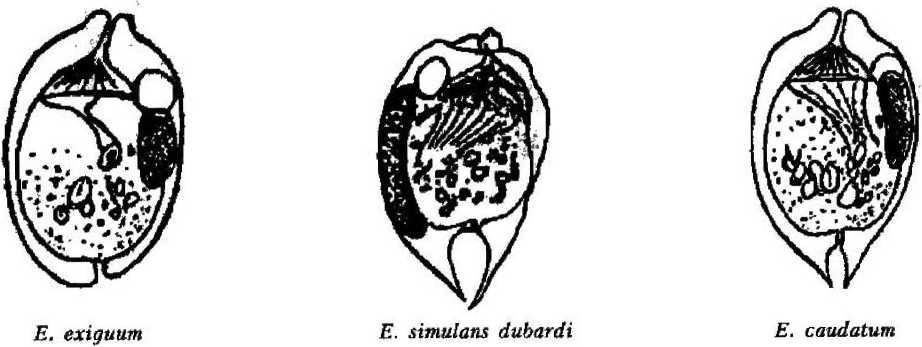
Рисунок / (продолжение)
Видовое разнообразие эндобионтных инфузорий желудка лося
Морфологические описания встреченных видов приводятся в работе в сопоставлении с данными литературы [2; 5; 8 — 10].
Основными морфологическими признаками эндобионтных инфузорий являются:
-
1) форма цитостомального (переднего) и цитопроктального (заднего) концов тела;
-
2) вооружение заднего конца тела и его модификации;
-
3) длина и ширина тела инфузорий;
-
4) строение ресничного аппарата и его расположение на теле;
-
5) строение ротового аппарата;
-
6) строение цитоскелета;
-
7) строение ядерного аппарата и локализация ядер в клетке;
-
8) строение сократительной вакуоли и локализация в клетке.
Покров клетки представлен пелликулой, имеющей идентичное с другими ресничными инфузориями строение: плазмалемма с подстилающими ее уплощенными альвеолами. Неспособность всех обнаруженных нами в желудках лосей видов изменять форму тела, вероятно, свидетельствует о наличии у них, как и у свободноживущих форм, хорошо развитого кортекса, или ци тоскелета. Тем не менее покровы клетки большинства видов имеют специфичные только для данной группы инфузорий черты строения. Прежде всего у большинства видов пелликула гладкая и часто практически лишена ресничного покрова;, либо содержит рудиментарные группы ресничек (род Entodinium) (см. рис. 1). Если имеются реснички, то они собраны в пучки (Е. bursa, Е. ecaudatum-ecaudatum, D. то-nacanthum). Лишь вид D, ruminautium имеет хорошо развитые реснички, покрывающие клетку ровными продольными рядами (см. рис. 1);
Отсутствие сплошного ресничного покрова, вероятно, связано с обитанием инфузорий в пищеварительном тракте, где реснички как локомоторные органеллы теряют значимость. Утрату ресничек можно рассматривать как морфофизиологическую адаптацию к жизни в среде с ограниченным пространством для плавания либо с его отсутствием.
Второй особенностью инфузорий является наличие разнообразных выростов (шипов), обычно расположенных на заднем конце тела (Е. bursa, Е. alces, Е. cauda- turn, D. monacanthum). У некоторых представителей рода Diplodinium задний конец тела имеет длинный, часто подвижный, серповидно изогнутый хвостовой отросток (D. bubalidis ssp. bubalidis, D. bubalidis ssp. censors) (см. рис. 1). Можно предположить, что.при отсутствии у инфузорий специальных прикрепительных органелл именно выросты, или шипы, препятствуют выносу эндобионтов из желудка в кишечник, т. е. являются морфологическими адаптациями, направленными на удержание инфузорий не только в организме хозяина, но и в специфичном для них отделе пищеварительного тракта — желудке.
Форма тела многих видов эндобионт-ных инфузорий желудка лосей удлиненноовальная, лишь у вида Е. ecaudatum-ecaudatum клетка имеет веретеновидную форму (см. рис. 1). В той или иной степени передний конец клетки расширен и уплощен, а у некоторых видов косо срезан (D. bubalidis ssp. bubalidis). У других передний конец сужен (Е. отпит, Е. cauda-tum). Задний конец тела всегда закруглен. Эти инфузории ведут пристеночный образ жизни, в толще пищевого комка не встречаются, и форма тела отвечает условиям существования и способствует передвижению в заполненном пищей желудке.
У всех видов описанных инфузорий на переднем конце тела открывается цитостом, или перистом. Можно говорить о наличии у некоторых видов перистомального диска. У найденных эндобионтных инфузорий отмечены две формы цитостомов:
-
1) цитостом в виде сифона, представляющий собой трубку, проходящую в продольном направлении почти до конца клетки;
-
2) сложный цитостом, имеющий отверстие, открывающееся на переднем конце тела, а на заднем конце — порошицу («анальное отверстие»).
Строение цитостома, несомненно, является морфофизиологической адаптацией к питанию клетчаткой. Его длина позволяет осуществлять долгий процесс расщепления. Наличие порошицы позволяет инфузориям пропускать растительные волокна через тело.
Для большинства видов характерна удлиненная (колбасовидная) форма макро-нуклеуса, лежащего вдоль спинной стенки тела (Е. nanellum, Е. ovinum, Е. caudatum, Е. simulans-dubardi, Е. dices, Е. ecaudatum-ecaudatum). У некоторых видов макронуклеус по форме ближе к округлому (Е. exiguum, D. ruminantium). У видов рода Diplodinium форма макронуклеуса специфична: он имеет расширенную переднюю часть и суженную — заднюю. У большинства инфузорий макронуклеусы крупные, что связано с их функцией регуляции всех процессов жизнедеятельности клетки за исключением полового размножения.
Микронуклеус у всех видов имеет небольшие размеры и у большинства прилежит к срединной части макронуклеуса. У диплодиниумов округлый микронуклеус находится в выемке макронуклеуса. Микронуклеус — это генеративное ядро, которое принимает участие только в процессе полового размножения инфузорий.
Сократительная вакуоль инфузорий рода Diplodinium представлена парными образованиями, причем их локализация зависит от вида простейших: D. monacanthum — сократительные вакуоли располагаются в середине клетки, D. rangiferi — одна вакуоль располагается ближе к переднему концу клетки и лежит над макронуклеусом, другая — располагается почти посередине клетки под макронуклеусом, у D. bubalidis ssp. bubalidis сократительные вакуоли лежат на противоположных концах клетки. Для Е. ecaudatum-ecaudatum характерны чечевицеобразные парные сократительные вакуоли, причем одна лежит немного позади переднего конца макронуклеуса, а другая — несколько впереди его заднего конца. У большинства видов эндобионтных инфузорий лося одна сократительная вакуоль, чаще округлой формы, расположенная в передней части клетки вблизи макронуклеуса (род Entodinium, D. bubalidis ssp. censors). Для D. ruminantium также характерна одна сократительная вакуоль, находящаяся вблизи цитостома. Сократительные вакуоли инфузорий — это еще и осморегулирующие вакуоли, поскольку они регулируют осмотическое давление простейших. В сравнении со свободноживущими видами у эндобионтов строение сократительных вакуолей упрощено, что связано с образом жизни последних. Сократительные вакуоли работают ритмически, то попеременно расширяясь и медленно заполняясь жидкостью, то сокращаясь и выталкивая содержимое наружу.
Эндобионтные инфузории желудка лося A. dices, обитающего на территории Челябинской, Тюменской и Омской областей, как, впрочем, и во всей Евразии, прежде не были изучены. Эти своеобразные простейшие, обнаруженные в пищеварительном тракте различных травоядных млекопитаю- щих и у некоторых видов хозяев отличающиеся многообразием видов (например, у лошадей количество видов превышает 100), у лосей имеют низкое видовое разнообразие, всего (15 видов).
Эндобионтные ресничные инфузории в общих чертах строения и функционирования соответствуют дифференциальному диагнозу класса: пелликула, .реснички, цитостом, дуализм ядер, конъюгация. Но в связи с обитанием в условиях желудка хозяина эндобионты приобрели ряд признаков и свойств, отличающих их от других представителей класса и указывающих на их специфичность и пищевую специализацию.
Важным аспектом биологии инфузорий является их питание. На основе микроскопирования организмов, а также пищевых частиц, находящихся в цитостомах и пищеварительных вакуолях фиксированных инфузорий, мы выявили три группы видов (рис. 2):
-
1) растительноядные инфузории — преимущественно питающиеся растительными волокнами или отдельными клетками растительных тканей;
-
2) крахмалоядные инфузории — предпочитающие в питании зерна крахмала;
-
3) хищные инфузории — питающиеся другими инфузориями и отчасти бактериями.
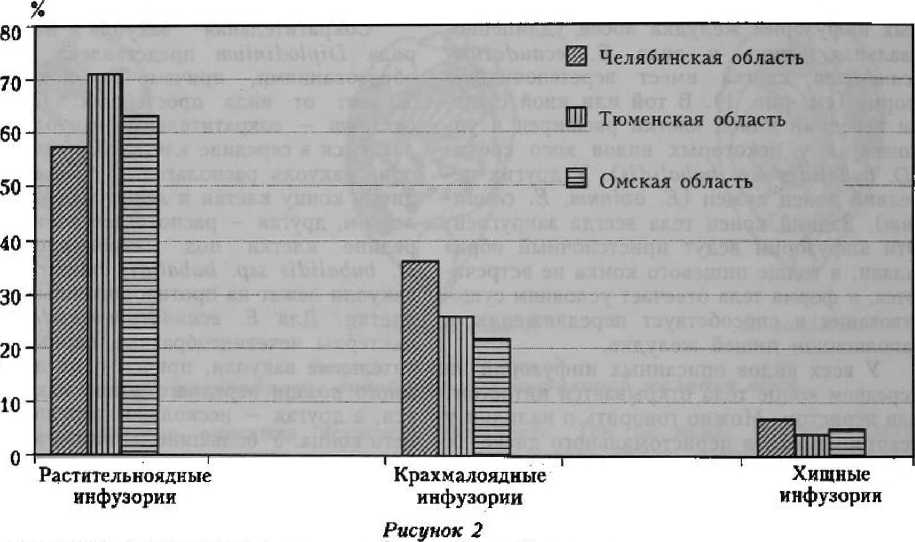
Процентное соотношение групп инфузорий, выделенных по характеру питания
На основе полученных данных выделять отдельную группу бактериоядных инфузорий нецелесообразно, как это делают некоторые исследователи, так как все инфузории поедают бактерий как дополнение к основной пище.
Выводы
-
1. Видовой состав эндобионтных инфузорий желудка лося представлен 15 видами, относящимися к четырем родам семейств Ophryoscolecidae и Isotrichidae. Основу фауны эндобионтных инфузорий лося по видовому разнообразию, частоте встречаемости и численности составляют 9 видов рода Entodinium,
-
2. Все найденные виды инфузорий являются форгутными, встречающимися только в желудке, так как в других отделах пищеварительного тракта лося инфузории не обнаружены.
-
3. Эндобионтные ресничные инфузории в общих чертах строения и функционирования соответствуют дифференциальному диагнозу класса, но в связи с обитанием в условиях желудка хозяина эндобионты приобрели ряд признаков и свойств, отличающих их от других представителей класса и указывающих на их специфичность и пищевую специализацию.
-
4. Инфузории, обитающие в желудке лося, составляют три «группы питания*: растительноядные, крахмалоядные и хищные. Растительноядные инфузории имеют наиболее высокую частоту встречаемости в пробах — от 57,3 до 74,1 %, крахмалоядные виды — от 21,5 до 36,0 %, а хищники — от 3,3 до 6,7 %.
Благодарность. Автор работы выражает искреннюю благодарность за научные консультации и помощь в проведении исследования доктору биологических наук, профессору С. Ф. Лихачеву, доктору био логических наук, профессору С. Н. Гаше-ву.
Список литературы Морфофизиологические особенности форгутных инфузорий лося как система адаптаций к эндобионтному обитанию
- Гассовский Г. Н. К микрофауне кишечника лошади/Г. Н. Гассовский//Труды Петроградского общества естествоиспытателей. -Л., 1918. -C. 20-37, 65-69.
- Догель В. А. Простейшие -Protozoa. Малоресничные инфузории -Infusoria Oligotricha. Сем. Ophryoscolecidae: определитель по фауне СССР/В. А. Догель. -Л: Изд.-во АН СССР, 1929. -96 с.
- Стрелков А. А. Паразитические инфузории из кишечника непарнокопытных сем. Equidae/А. А. Стрелков//Учен. записки ЛГПИ им. А. И. Герцена, фак. естеств. наук, каф. зоологии. -1939. -С. 314-318.
- Cameron S. L. The ultrastructure of Amylovorax dehorityi comb. nov. and erection of the Amylovoracidae fam. nov. (Ciliophora: Trichostomata)/S. L. Cameron, P. J. O'Donaghue//Eur. Jou rn. Protistology. -2002. -P. 29-44.
- Dehority B. A. A new family of entodiniomorph protozoa from the marsupial forestomach, with descri ptions of a new genus and five new species/B. A. Dehority//Journ. of Eucaryotic Microbiology. -1996. -№ 43. -P. 285-295.
- Fiorentini A. Su r les Protistes de l'estomac des Bovides/A. Fiorentini//Journ. de Micrographie. -1890. -P. 14.
- Hsiung T. S. A monograph on the protozoan fauna of the large intestine of the horse/T. S. Hsiung//Iowa St. Coll. J Sci. -1930. -№ 64. -P. 359-423.
- Lubinsky G. Studies on the evolution of the Ophryoseolecidae: some evolutionary trends on the genus Entodinium/G. Lubinsky//Canad. J. Zool. -1957. -Vol. 35, № 1. -P. 111-133.
- Lubinsky G. Studies on the evolution of the Ophryoscolecidae. On the origin of the higher O phryoscolecidae/G. Lubinsky//Canad. Journ. Zool. -1957. -Vol. 35, № 1. -P. 135-140.
- Lubinsky G. Studies on the evolution of the Ophryoscolecidae. Phylogeny of the O phryoscolecidae based on their comparative morphology/G. Lubinsky//Canad. Journ. Zool. -1957. -Vol. 35, № 1. -P. 141-159.
