Морфофункциональная оценка FAP+ и -SMA+-клеток в различные сроки токсического фиброза печени у крыс

Автор: Лебедева Е.И.
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 3 т.31, 2023 года.
Бесплатный доступ
Углубленное изучение источника фиброгенной популяции клеток применительно к этиологии и стадии фиброза, а также понимание молекулярных механизмов, регулирующих изменение фенотипа печеночных фибробластов, имеют первостепенное значение при разработке фармакологических препаратов. Цель исследования - морфофункциональная оценка активированных портальных фибробластов (FAP+) и жир-накапливающих клеток (α-SMA+) печени в различные сроки токсического фиброза печени крыс. Фиброз и цирроз печени у крыс-самцов Wistar индуцировали раствором тиоацетамида в течение 17 недель. Морфологическое исследование печени проводили на парафиновых срезах, окрашенных гематоксилином и эозином, методом Маллори, иммуногистохимическое исследование производили с помощью поликлональных кроличьих антител к антигену портальных фибробластов FAP и с помощью моноклональных мышиных антител к антигену α-SMA+ клеток. До наступления стадии фиброза печени F3/F4 с 3 по 7 неделю количество FAP+ и α-SMA+-клеток увеличивалось попеременно. На этапах трансформации фиброза в цирроз с 7 по 11 неделю повышение их количества происходило незначительно. На стадии неполного (F5) и до наступления достоверного цирроза (F6) с 11 по 15 неделю количество FAP+ и α-SMA+-клеток было непостоянным и происходило попеременное увеличение и уменьшение их количества. α-SMA+-клетки до начала процесса трансформации фиброза в цирроз (F4/F5) наблюдали в синусоидах и очагах некроза. Затем они выявлялись как в синусоидах, так и в соединительнотканных трабекулах. FAP+-клетки на стадии портального фиброза (F1) локализовались около междольковых сосудов и междольковых желчных протоков портальных зон, а с периода F2/F3 их выявляли в соединительнотканных трабекулах и синусоидах. В количественном отношении на всех стадиях фиброза преобладали α-SMA+-клетки. На основании полученных результатов можно предположить, что FAP+-клетки вносят основной вклад в развитие портального и начального этапа мостовидного фиброза. Их следует рассматривать как одну из популяций миофибробластов при тиоацетамид-индуцированном фиброгенезе печени.
Печень, крысы, фиброз, fap+ клетки, α-sma+ клетки
Короткий адрес: https://sciup.org/143181181
IDR: 143181181 | DOI: 10.20340/mv-mn.2023.31(3).794
The morphological and functional assessment of FAP+ and -SMA+-cells in different times of rat toxic liver fibrosis
Qualitative study of the source of the fibro-genic cell population in relation to the etiology and stage of fibrosis, as well as an understanding of the molecular mechanisms that regulate changes in the phenotype of hepatic fibroblasts, are of paramount importance in the development of pharmacological drugs. The purpose of the study was a morphological and functional assessment of activated portal fibroblasts (FAP+) and fat-accumulating cells (α-SMA+) of the liver at various stages of toxic liver fibrosis in rats. Liver fibrosis and cirrhosis in male Wistar rats were induced with thioacetamide solution for 17 weeks. Morphological examination of the liver was carried out on paraffin sections stained with hematoxylin and eosin using the Mallory method; immunohistochemical examination was carried out using polyclonal rabbit antibodies to the portal fibroblast antigen FAP and using monoclonal mouse antibodies to the α-SMA+ cell antigen. Before the onset of liver fibrosis stage F3/F4, from weeks 3 to 7, the number of FAP+ and α-SMA+ cells increased alternately. During the stages of transformation of fibrosis into cirrhosis from 7 to 11 weeks, their number increased slightly. At the stage of incomplete (F5) and before the onset of significant cirrhosis (F6) from weeks 11 to 15, the number of FAP+ and α-SMA+ cells were inconsistent and there was an alternating increase and decrease in their number. α-SMA+ cells before the start of the process of transformation of fibrosis into cirrhosis (F4/F5) were observed in sinusoids and foci of necrosis. Then they were detected both in sinusoids and in connective tissue trabeculae. FAP+ cells at the stage of portal fibrosis (F1) were localized near the interlobular vessels and interlobular bile ducts of the portal zones, and from the F2/F3 period they were detected in connective tissue trabeculae and sinusoids. In quantitative terms, α-SMA+ cells predominated at all stages of fibrosis. Based on the results obtained, it can be assumed that FAP+ cells make a major contribution to the development of the portal and initial stages of bridging fibrosis. They should be considered as one of the myofibroblast populations in thioacetamide-induced liver fibrogenesis.
Текст научной статьи Морфофункциональная оценка FAP+ и -SMA+-клеток в различные сроки токсического фиброза печени у крыс
Lebedeva EI. The morphological and functional assessment of FAP+ and α-SMA+-cells in different times of rat toxic liver fibrosis. Morfologicheskie Vedomosti – Morphological newsletter. 2023;31(3):794. (3).794
Введение. При хронических заболеваниях печени разрастание фиброзной соединительной ткани связано с их этиологией, стадией фиброза и обусловлено гетерогенностью популяцией миофибробластов, вовлеченных в процесс, жиронакапливающих клеток, портальных фибробластов, циркулирующих фиброцитов, гемопоэтических и мезенхимальных стволовых клеток костного мозга [1-4]. Жиронакапливающие клетки (далее - ЖК) считают основной клеточной популяцией, синтезирующей межклеточное вещество [4]. В литературе они имеют разные названия: клетки Ито, липоциты печени, перисинусоидные клетки, перициты и звездчатые клетки. Эмбриональное происхождение ЖК не установлено, но известно, что они экспрессируют белки энтодермы и мезодермы. В здоровой печени ЖК располагаются в перисинусоидном пространстве, контактируя с гепатоцитами, звездчатыми макрофагами и эндотелиоцитами, проявляя не активированный фенотип [5]. В норме ЖК неправильной формы, с округлым ядром, накапливают витамин А и жиры, синтезируют цитокины, матриксные металлопротеиназы и их тканевые ингибиторы; они практически не экспрессируют гладкомышечный альфа-актин (a-SMA). ЖК регулируют кровоток в синусоидах, функционируя как перициты, обладают низкой пролиферативной активностью и способностью к секреции коллагенов [4-5]. Токсические и вирусные поражения печени стимулируют активацию и трансдифференцировку ЖК в миофибробластический фенотип [6-7]. Этот процесс сопровождается гиперплазией и гипертрофией шероховатой эндоплазматической сети, исчезновением в цитоплазме липидных капель, изменением организации цитоскелета, избыточной экспрессией a-SMA, коллагенов и цитокинов. Механизмы активации ЖК, особенно в фазу ее инициации, весьма сложны и не до конца изучены. Предполагается, что пул покоящихся и активированных ЖК пополняется также за счет клеток костного мозга [4,8].
Портальные фибробласты (далее -ПФ) или перидуктулярные мезенхимальные клетки, являются второй популяцией клеток, вырабатывающих межклеточное вещество при фиброзе печени [1]. Сведений о функциях ПФ в норме недостаточно. Расположены они в соединительной ткани портальных зон, преимущественно вокруг междольковых желчных протоков. Происхождение и роль ПФ в развитии холестатического фиброза до сих пор остаются дискуссионными. Окончательно не установлено, являются ли ПФ основной популяцией миофибробластов и на каких этапах фиброгенеза их вклад наиболее значителен, синтезируют ли ПФ фибриллярные коллагены, типичные для фиброза или они выполняют другие роли в фиброге-незе [9-10]. Одни авторы предполагают, что активированные ПФ принимают участие в секреции межклеточного вещества только при хронических холестатических заболеваниях печени [1]. В экспериментальных работах с использованием репортерных Col-GFP и Mdr2-/- мышей на начальных стадиях холестатического фиброгенеза источником миофибробластов выступали активированные ПФ. При этом дальнейшее прогрессирование фиброза приводило к трансдифференцировке ЖК в фиброгенный фенотип [10]. Научные работы последних лет указывают, что при холестатическом фиброзе одновременно идентифицировались две разные популяции миофибробластов, происходящие из ПФ и ЖК [9-10]. Вероятно, эти противоречивые данные являются результатом различий используемых экспериментальных моделей, методов исследований и как следствие отличий в интерпретации полученных результатов. Изучение фиброгенной популяции клеток применительно к этиологии и стадии фиброза печени может имеет существенное значение в разработке антифибротических препаратов [11] .
Цель исследования: морфофункциональная оценка активированных портальных фибробластов (FAP + ) и жиронакапливающих клеток (a-SMA + ) печени в различные сроки токсического фиброза печени крыс.
Материалы и методы исследования. В настоящей публикации использованы международные термины по цитологии и гистологии [12]. В экспериментах использовались крысы-самцы породы Ви-стар массой 190-210 г. Протокол исследования одобрен комиссией по биоэтике и гуманному обращению с лабораторными животными Витебского государственного медицинского университета (протокол № 6 от 03.01.2019 г.). Животных содержали в пластико-металлических клетках по 6 особей при естественном освещении и со свободным доступом к корму и воде. Температура в помещении вивария поддерживалась на уровне 21-23°С, влажность воздуха составляла около 50%. Фиброз и цирроз печени моделировали путем хронической интоксикации тиоацетамидом (далее - ТАА), (Acros Organics). Свежеприготовленный раствор ТАА вводили интрага-стрально через зонд в дозе 200 мг/кг массы тела 2 раза в неделю в течение 17 недель. Крысы контрольной группы (n=12) получали воду без ТАА в аналогичном объеме. Животных рандомизировали на 8 групп по 12 животных в каждой в зависимости от длительности воздействия ТАА: 3 недели (1-я группа), 5 недель (2-я группа), 7 недель (3-я группа), 9 недель (4-я группа), 11 недель (5-я группа), 13 недель (6-я группа), 15 недель (7-я группа), 17 недель (8-я группа).
После декапитации под кратковременным эфирным наркозом из большой левой доли печени крыс забирали образцы печени диаметром 5-10 мм, которые помещали в 10% раствор нейтрального формалина на фосфатном буфере и фиксировали в течение 24 ч. Затем проводили обработку фиксированного материала с последующей заливкой в парафин с использованием автомата для гистологической обработки ткани STP-120 (Thermo Fisher Scientific) и станции для заливки ткани парафином EC350 (Thermo Fisher Scientific). От каждого животного получали по одному блоку для каждого метода окрашивания и с помощью ротационного микротома НМ340Е (MICROM, Laborgerate GmbH) готовили по 3-4 среза толщиной 4 мкм и помещали их на предметные стекла. Для получения обзорных гистологических препаратов срезы печени окрашивали гематоксилином и эозином, а для выявления соединительной ткани – по Маллори с помощью автомата для окраски HMS70 (Thermo Fisher Scientific). Степень фиброза определяли с помощью полуколичествен-ной шкалы K.G. Ishak [13].
Иммуногистохимическое исследование проводили на парафиновых срезах [14]. В качестве маркера активированных портальных фибробластов применяли по- ликлональные кроличьи антитела FAP (FAP-alpha, пролилэндопептидаза FAP, в разведении 1:100), активированных ЖК – моноклональные мышиные антитела к α-SMA (alpha-SMA, АСТА2, в разведении 1:1000). Использовались антитела производства Wuman Elabscience Biotechnology Incorporated Company, набор 2-step plus Poly-HRP Anti Rabbit/Mouse IgG Detection System/with DAB Solution; Retrieve-All Antigen (Unmasking System Basic), буфер для разведения антител (BioLegend), Твин-20 (Glentham Life Sciences), PBS (Melford). Для лучшей ориентации в препарате и правильной идентификации клеток, содержащих искомый антиген, срезы докрашивали гематоксилином Майера в течение 1 мин. Для объективной интерпретации результатов для каждой исследуемой серии (группы животных) включали положительный и отрицательный контроли. Иммуногистохимическую окраску оценивали, как положительную, только при отсутствии окрашивания в отрицательном контроле и, наоборот, как отрицательную при окрашивании в положительном контроле.
Гистологические препараты микро-скопировали с применением компьютерных программ анализа изображений ImageScope Color и cellSens Standard. Количество FAP-позитивных клеток (FAP+-клетки) и α-SMA-позитивных клеток (α-SMA+-клетки) подсчитывали в трех полях зрения каждого гистологического среза на базе микроскопа Olympus BX51 при увеличении объектива 40×.
Полученные результаты обрабатывались статистически. Для каждой выборки определяли нормальность частотного распределения каждого признака. Выборки не являлись малыми (n=60>50), поэтому проверку осуществляли по критерию Лиллие-форса. Получали описательные статистики и описывали количественные экспериментальные данные в виде средних и их соответствующих доверительных интервалов (М (95% ДИ: j-q)), медианы и значения 15-го – 85-го процентилей (Ме (15%;85%)). Об уровне статистической значимости различий судили по t-критерию Стьюдента; в случае отличия выборок от нормального частотного распределения использовали U-критерий Манна-Уитни.
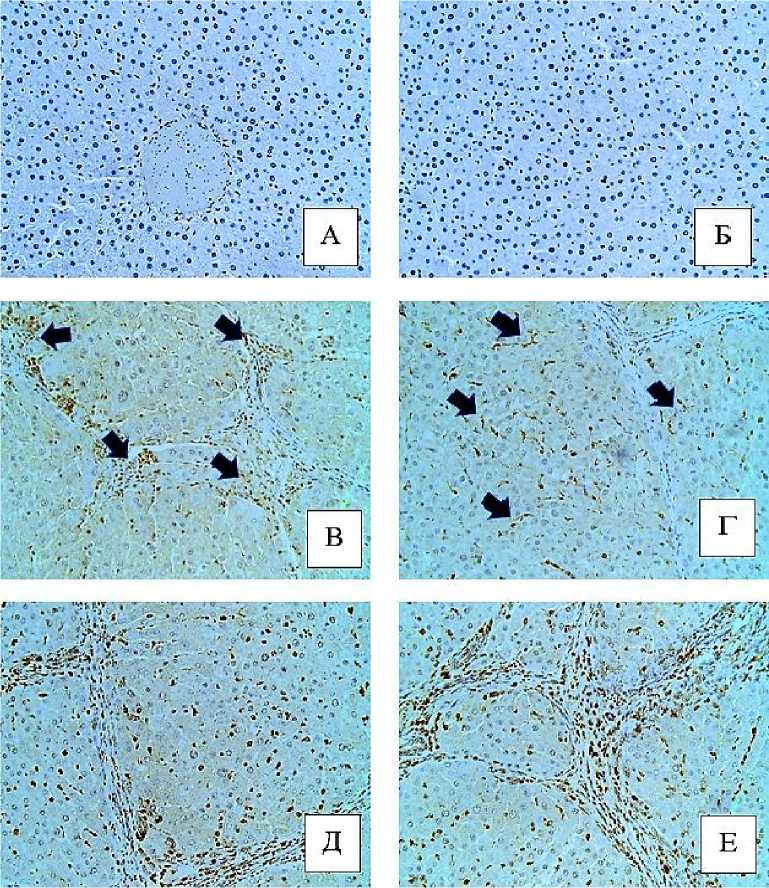
Рис. 1. Микрофото гистологических препаратов печени крыс. Окр.: иммуногистохимическое окрашивание на FAP + (А, В, Д), на a-SMA+ (Б, Г, Е), докраска гематоксилином Майера. Ув.: х400. Обозначения: А - печень крысы контрольной группы; Б - печень крысы контрольной группы; В - печень крысы с индуцированным циррозом через 9 недель после начала эксперимента; Г - печень крысы с индуцированным циррозом через 9 недель после начала эксперимента; Д - печень крысы с индуцированным циррозом через 15 недель после начала эксперимента; Е - печень крысы с индуцированным циррозом через 15
недель после начала эксперимента
Результаты исследования и обсуждение. Углубленное изучение источника миофибробластов и понимание молекулярных механизмом, регулирующих изменение фенотипа клеток печени в фиброгенную линию, имеют фундаментальное значения для разработки анти-фибротических препаратов [1-4, 11, 15]. При фиброзе органов миофибробласты преимущественно идентифицируют с помощью маркеров a-SMA (активированные ЖК) и FAP (активированные ПФ) [16-18]. В большинстве органов в условиях нормы уровни мРНК и белка FAP были низкими, за исключением молочной железы, эндометрия матки, поджелудочной железы, плаценты и кожи. В здоровой печени ЖК практически не экспрессируют маркер a-SMA [2, 4-5, 15]. Полученные в рамках настоящего исследования результаты согласуются с литературными данными. В печени контрольных крыс FAP+-клетки не определяли (рис. 1-А). В синусоидах отсутствовали a-SMA+-клетки (рис. 1-Б), однако в отдельных случаях их выявляли в стенках междольковых артерий, междольковых и поддольковых вен.
При наступлении портального фиброза печени (F1, 3 недели интоксикации) в портальных зонах наблюдали FAP+-клетки, а в синусоидах – α-SMA+-клетки. Обе популяции клеток имели округлую форму. Спустя 5 недель с появлением портального, мостовидного, местами центролобулярного и диффузного перицел- люлярного фиброза (F2/F3) количество FAP+-клеток достоверно увеличилось (р=0,0213), а α-SMA+-клеток – не изменилось (р=0,3075) по сравнению с 3 неделей. На всех последующих стадиях эксперимента установлены достоверные различия как для FAP+-клеток (р=0,00001), так и для α-SMA+-клеток (р=0,00001) по сравнению с 3 неделей эксперимента. В таблице 1 представлена динамика числа FAP+- и α-SMA+-клеток на всех этапах эксперимента.
Таблица 1
Относительная величина площади соединительной ткани и количества FAP+- и a-SMA+- клеток в различные сроки фиброза и цирроза печени крыс при интоксикации тиоацетамидом
|
Недели эксперимента/ стадии фиброза |
Площадь соединительной ткани в %, Ме (15%,'85%) |
Количество клеток, М (95% ДИ: j-q) |
|
|
FAP + |
a-SMA+ |
||
|
контроль |
1,50 (1,000;3,000) |
- |
- |
|
3 недели/F1 |
3,00 (2,000;5,000) |
5,416 (4,814;6,018) |
7,722 (6,426;9,01) |
|
5 недель/F2/F3 |
4,00 (2,000;6,500) |
9,194 (8,080;10,308) |
11,777 (10,480;13,074) |
|
7 недель/F3/F4 |
5,00 (2,000;8,000) |
11,972 (10,361;13,582) |
18,888 (17,283;20,493) |
|
9 недель/F4/F5 |
7,00 (4,000;10,000) |
14,694 (13,083;16,305) |
20,861 (18,958;22,763) |
|
11 недель/F5 |
9,00 (6,000;11,000) |
17,222 (15,979;18,465) |
25,250 (23,064;27,435) |
|
13 недель/F6 |
11,00 (7,500;14,500) |
21,944 (20,251;23,637) |
31,500 (28,381;34,618) |
|
15 недель/F6 |
16,00 (9,000;21,500) |
24,888 (22,483;27,294) |
52,722 (48,076;57,377) |
|
17 недель/F6 |
21,00 (13,000;29,000) |
29,222 (27,143;31,301) |
58,944 (54,318;63,570) |
Следует отметить, что по мере прогрессирования фиброза печени скорость разрастания соединительной ткани была не одинаковой (табл. 1). К 3-й неделе эксперимента отмечали умеренное формирование соединительной ткани в портальных зонах (рис. 2-А, рис. 2-Б). Однако, на 5-й неделе образование фиброзной соединительной ткани замедлялось, при этом ее одновременно определяли, как в портальных зонах, так и в паренхиме. К 7й неделе эксперимента интенсивность синтеза соединительной ткани сохранилась практически на прежнем уровне по сравнению с 5-й неделей (рис. 2-В). На этапе перестройки паренхимы печени и перехода от фиброза к циррозу вновь наблюдалось усиленное формирование соединительной ткани аналогичное началу эксперимента (3 неделя). На 9-й неделе происходило образование ложных печеночных долек у портальных зон (рис. 2-Г), что являлось морфологическим критерием начальной трансформации фиброза в цирроз. В период с 11-й по 17-ю неделю разрастание соединительной ткани достигало максимально значения (рис. 2-Д, рис. 2-Е). При попарном сравнении на разных стадиях эксперимента применительно к площади соединительной ткани, количеству FAP+ и α-SMA+-клеток получены отличительные результаты (табл. 2).
При анализе таблицы 2 следует, что до наступления стадии фиброза печени F3/F4 (3-й по 7-ю неделю) количество FAP+ и α-SMA+-клеток увеличивалось попеременно. На этапах трансформации фиброза в цирроз (с 7-й по 11-ю неделю) повышение их количества происходило незначительно. На стадии неполного (F5) и до наступления достоверного цирроза (F6, с 11-й по 15-ю неделю) количество FAP+ и α-SMA+-клеток было непостоянным: происходило попеременное увеличе-
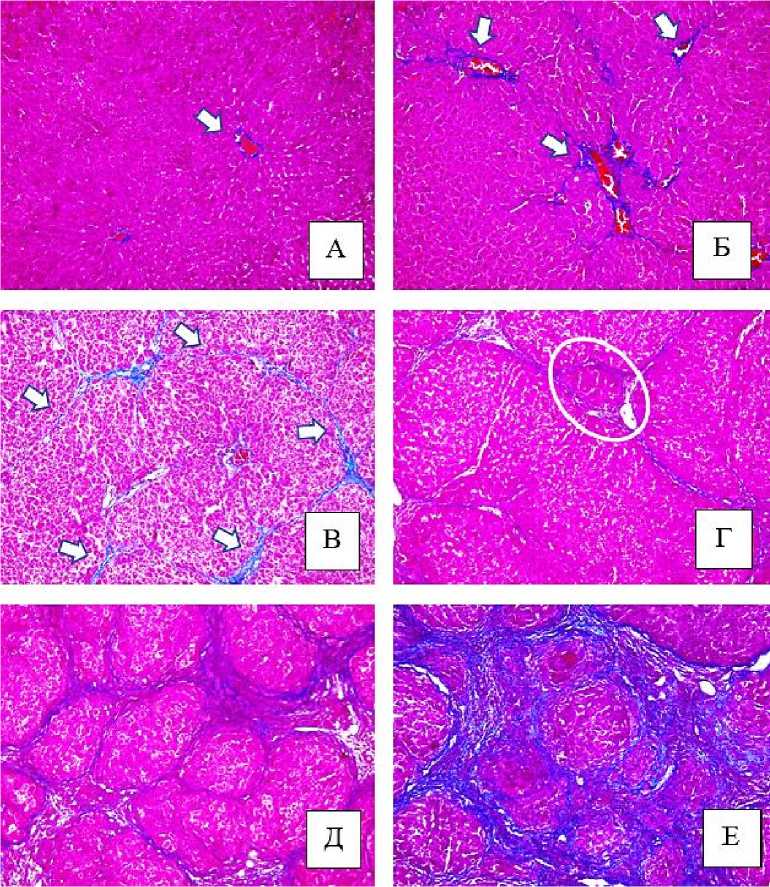
Рис. 2. Микрофото гистологических препаратов печени крыс. Окр.: по Маллори. Ув.: х200. Обозначения: А – печень крысы контрольной группы, стрелками отмечена соединительная ткань; Б – печень крысы с индуцированным циррозом через 3 недели после начала эксперимента, стрелками отмечена соединительная ткань; В – печень крысы с индуцированным циррозом через 7 недель после начала эксперимента, стрелками отмечена соединительная ткань; Г – печень крысы с индуцированным циррозом через 9 недель после начала эксперимента; Д – печень крысы с индуцированным циррозом через 13 недель; Е -печень крысы с индуцированным циррозом через 17 недель после начала эксперимента
ние и уменьшение их количества. В настоящее время объяснить такие количественные вариации не представляется возможным. Предположительно, это связано с компенсаторно-приспособительными механизмами, происходящими в печени, индуцированием/ингибировани-ем молекулярных сигналов, регулирующих пролиферацию этих клеток или другими процессами. С помощью иммуногистохимических методов было установлено, что две морфологически неоднород-
Таблица 2
Показатели значимости попарных различий площади соединительной ткани и числа FAP+ и α-SMA+-клеток
FAP+-клетки округой формы через 3 недели эксперимента располагались вокруг междольковых сосудов и вблизи междольковых желчных протоков портальных зон, а со стадии F2/F3 их выявляли в соединительнотканных септах и синусоидах (рис. 1-В, рис. 10-Д). На данном этапе рядом с желчными протоками концентрировались крупные СК19+-клетки. По морфологическим признакам они соответствуют печеночным стволовым/овальным клеткам. С 5-й недели эксперимента отмечали увеличение числа междольковых желчных протоков и к его концу вокруг одной портальной зоны можно было обнаружить более 30 их поперечных сечений (протоковая реакция) [20]. Установлено, что эпителиальные клетки протоковой реакции экспрессируют профиброгенные факторы (TGF-в, PDGF) и могут активировать ПФ [21]. На гистологических препаратах отмечалось направленное разрастание волокон соединительной ткани с FAP+-клетками от двух портальных зон через паренхиму печени навстречу друг другу, таким образом, предопределяя путь для фиброзной соединительной ткани и формирования соединительнотканных мостов - мостовидный фиброз. Важно отметить, что образование ложных печеночных долек происходило у портальных зон. Предположительно, FAP+-клетки первыми синтезируют межклеточное вещество в пор- тальных зонах и, возможно, регулируют его секрецию, участвуют в формировании соединительнотканных септ (мостов). а-БМА+-клетки выступают в роли миофибробластов позже. Вероятно, этап активации и трансдифференцировки ЖК более продолжительный или молекулярные сигналы, регулирующие смену фенотипа клеток, индуцируются с опозданием, или до определенного момента ингибируются. Не следует исключать тот факт, что на ранних стадиях фиброгенеза ЖК выполняют другие функции. На 5-й неделе количество а-БМА+-клеток не изменилось (р=0,3075) по сравнению с 3-й неделей эксперимента. При этом отмечалось повышение числа FAP+-клеток (табл. 1 и 2). Для объективной интерпретации того, какая из клеточных популяций доминирует в синтезе межклеточного вещества на ранних этапах фиброза требуются более детальные исследования.
FAP - это многофункциональный белок. Он принимает участие в регуляции энергетического и липидного обменов, стимулирует воспаление и выполняет профиброгенную функцию. Роль белка FAP в фиброзе печени остается до конца не изученной и его фармакологическое ингибирование в качестве варианта потенциальной терапии фиброза печени до сих пор не проводилось [2, 11, 22-24]. В печени крыс при тиоацетамид-индуцированном фиброзе и циррозе жировая дистрофия не диагностировалась. Лимфоидно-гистиоцитарный инфильтрат до 9-й недели был выражен незначительно, а к концу эксперимента определялись очаги с лимфоидно-гистиоцитарным инфильтратом в соединительной ткани портальных зон и соединительнотканных септ. На основании этого можно заключить, что функции белка FAP обусловлены состоянием плотности соединительной ткани, клеточным составом, активностью генов и эпигенетическими механизмами.
Заключение. Таким образом, при экспериментальном токсическом фиброзе и циррозе в печени крыс иммуногистохимическим методом одновременно были выявлены две морфологически неоднородные популяции миофибробластов, которые экспрессировали различные типы маркеров. а-БМЛ+-клетки на ранних стадиях фиброза не располагались совместно с FAP+-клетками. До начала процесса трансформации фиброза печени в цирроз (F4/F5) а-БМЛ+-клетки отмечали в синусоидах печени и очагах некроза, а затем они локализовались как в синусоидах, так и в соединительнотканных септах, вокруг ложных печеночных долек. FAP+-клетки на стадии портального фиброза располагались около междольковых сосудов и междольковых желчных протоков портальных зон, а со стадии F2/F3 их выявляли в соединительнотканных септах и синусоидах. В количественном отношении на всех стадиях фиброза преобладали а-БМЛ+-клетки.
На основании полученных результатов можно предположить, что FAP+-клетки играют главную роль в развитии портального и начального этапа мостовидного фиброза. Их следует рассматривать как одну из разновидностей популяции миофибробластов при тиоацетамид-индуцированном фиброгенезе печени. РЛГ+-клетки могут быть использованы в качестве мишени для антифибротической терапии.
Список литературы Морфофункциональная оценка FAP+ и -SMA+-клеток в различные сроки токсического фиброза печени у крыс
- Baglieri J, Brenner DA, Kisseleva T. The Role of Fibrosis and Liver-Associated Fibroblasts in the Pathogenesis of Hepatocellular Carcinoma. Int J Mol Sci. 2019;20(7). 1723.DOI. 10.3390/ijms20071723
- Lay AJ, Zhang HE, McCaughan GW et al. Fibroblast activation protein in liver fibrosis. Front Biosci (Landmark Ed). 2019;24(1).1-17. DOI.10.2741/4706
- Dhar D, Baglieri J, Kisseleva T et al. Mechanisms of liver fibrosis and its role in liver cancer. Exp Biol Med (Maywood). 2020;245(2).96-108. D0I.10.1177/1535370219898141
- Luo N, Li J, Wei Y et al. Hepatic Stellate Cell. A Double-Edged Sword in the Liver. Physiol Res. 2021;70(6).821-829. DOI. 10.33549/physiolres. 934755
- Sufletel RT, Melincovici CS, Gheban BA et al. Hepatic stellate cells - from past till present. morphology, human markers, human cell lines, behavior in normal and liver pathology. Rom J Morphol Embryol. 2020;61(3).615-642. D0I.10.47162/RJME.61.3.01
- S0rensen MD, Thiele M, Krag A et al. Stage-dependent expression of fibrogenic markers in alcohol-related liver disease. Pathol Res Pract. 2022;231.153798. DOI. 10.1016/j.prp.2022.153798
- Higashi T, Friedman SL, Hoshida Y. Hepatic stellate cells as key target in liver fibrosis. Adv Drug Deliv Rev. 2017;121.27-42. DOI. 10.1016/j .addr.2017.05.007
- Sun Y, Liu B, Xie J et al. Aspirin attenuates liver fibrosis by suppressing TGF ß1/Smad signaling. Mol Med Rep. 2022;25(5):181. DOI.10.3892/mmr.2022.12697
- Wells RG. Portal Fibroblasts in Biliary Fibrosis. Curr Pathobiol Rep. 2014;2(4).185-190. DOI. 10.1007/s40139-014-0054-y
- Fuji H, Miller G, Nishio T et al. The role of mesothelin signaling in portal fibroblasts in the pathogenesis of cholestatic liver fibrosis. Front Mol Biosci. 2021;8.790032. DOI. 10.3389/fmolb.2021.790032
- Yang AT, Kim YO, Yan XZ et al. Fibroblast Activation Protein Activates Macrophages and Promotes Parenchymal Liver Inflammation and Fibrosis. Cell Mol Gastroenterol Hepatol. 2022.S2352-345X(22)00253-3. DOI. 10.1016/j.jcmgh.2022.12.005
- Banin VV, Belousova TA, Bykov VL i dr. Terminologia Histologica. Mezhdunarodnye terminy po tsitologii i gistologii cheloveka s ofitsial'nym spiskom russkikh ekvivalentov. spravochnoe posobie. M.. GEOTAR-Media, 2009.- 272s. In Russian
- Everhart JE, Wright EC, Goodman ZD et al. HALT-C Trial Group. Prognostic value of Ishak fibrosis stage. findings from the hepatitis C antiviral long-term treatment against cirrhosis trial//Hepatology. 2010. Vol. 51, N 2. P. 585-594. DOI. 10.1002/hep.23315
- Kirik OV, Karpenklo MN, Korzhevskiy DE. Teoreticheskie osnovy i prakticheskoe primenenie metodov immunogistokhimii. Rukovodstvo. S-Pb.. SpetsLit, 2014.- 119s. In Russian
- Avery D, Govindaraju P, Jacob M et al. Extracellular matrix directs phenotypic heterogeneity of activated fibroblasts. Matrix Biol. 2018;67.90-106. DOI. 10.1016/j.matbio.2017.12.003
- Younis NS, Ghanim AMH, Elmorsy MA et al. Taurine ameliorates thioacetamide induced liver fibrosis in rats via modulation of toll like receptor 4/nuclear factor kappa B signaling pathway. Sci Rep. 2021 Jun 10;11(1).12296. DOI. 10.1038/s41598-021-91666-6
- Voutilainen SH, Kosola SK, Lohi J et al. Expression of 6 Biomarkers in Liver Grafts After Pediatric Liver Transplantation. Correlations with Histology, Biochemistry, and Outcome. Ann Transplant. 2020;25.e925980. DOI. 10.12659/AOT.925980
- Fitzgerald AA, Weiner LM. The role of fibroblast activation protein in health and malignancy. Cancer Metastasis Rev. 2020;39(3).783-803. DOI. 10.1007/s10555-020-09909-3
- Wu Y, Li Z, Xiu AY et al. Carvedilol attenuates carbon tetrachloride-induced liver fibrosis and hepatic sinusoidal capillarization in mice Drug Des Devel Ther. 2019;13.2667-2676. DOI. 10.2147/DDDT.S210797
- Lebedeva EI. The Role of CK19-Positive Portal Zone Cells in Thioacetamide Induced Rat Liver Cirrhosis. Cell and Tissue Biology. 2021;15(6).568-576. DOI. 10.1134/S1990519X21060067
- Lemoinne S, Thabut D, Housset C. Portal myofibroblasts connect angiogenesis and fibrosis in liver. Cell Tissue Res. 2016;365(3).583-589. DOI. 10.1007/s00441-016-2443-5
- Dendl K, Koerber SA, Kratochwil C et al. FAP and FAPI-PET/CT in Malignant and Non-Malignant Diseases. A Perfect Symbiosis? Cancers (Basel). 2021;13(19).4946. DOI. 10.3390/cancers13194946
- Fitzgerald AA, Weiner LM. The role of fibroblast activation protein in health and malignancy. Cancer Metastasis Rev. 2020;39(3).783-803. DOI. 10.1007/s10555-020-09909-3
- Hamson EJ, Keane FM, Tholen S et al. Understanding fibroblast activation protein (FAP). substrates, activities, expression and targeting for cancer therapy. Proteomics Clin Appl. 2014;8(5-6).454-463. DOI. 10.1002/prca.201300095
