Морфофункциональная реакция Iba-I и MHC-II позитивных клеток селезенки на введение мелатонина

Автор: Лузикова Елена Михайловна, Шатских О.А., Ефремова Ольга Александровна, Сергеева Валентина Ефремовна
Журнал: Морфологические ведомости @morpholetter
Рубрика: Краткие сообщения
Статья в выпуске: 4 т.24, 2016 года.
Бесплатный доступ
Изучена реакция Iba-I и MHC-II позитивных клеток селезенки на введение мелатонина. Установлено, что введение препарата мелатонин «Мелаксен») с водой перорально (раствор 4 мг препарата на 1л Н О) в течение 4 недель приводит к морфологическим изменениям: уменьшению среднего количества клеток макрофагального ряда (Iba-1+) и активированных макрофагов (MHCII+) в селезенке животных, содержавшихся при естественном освещении; незначительные изменения среднего количества клеток макрофагального ряда (Iba -1+) и активированных макрофагов (MHCII+) в селезенке животных, содержавшихся при затемнении. Наиболее чувствительными к световым условиям содержания животных, которым вводился мелатонин, являются ПАЛМ (периартериолярная лимфоидная муфта), маргинальный синус и субкапсулярная зона красной пульпы селезенки.
Селезенка, мелатонин, антигенпрезентирующие клетки, макрофаги
Короткий адрес: https://sciup.org/143177154
IDR: 143177154
Morphofunctional reaction of Iba-I and MHC-II positive cells of the spleen on the injection of melatonin
Reaction of Iba-I and MHC-II positive eells in the spleen was studied on the introduction of melatonin. It was detected that oral administration of melatonin (drug «Melaxen») with water (solution 4 mg of the drug per 1 l H20) for 4 weeks leads to morphological changes in the spleen: increase in quantity of lymphoid nodules, more pronounced in the spleen of animals kept by artificial darkening; decrease in the quantity of eells of the macrophage series (Iba-1+) and activated macrophages (MHCII+) in the spleen of animals kept by daylight; low changes in the quantity of eells of the macrophage series (Iba-1+) and activated macrophages (MHCII+) in the spleen of animals kept by artificial darkening. The most sensitive to lighting conditions animals injected with melatonin, are PALM, marginal sinus and subeapsular area of the red pulp of the spleen.
Текст краткого сообщения Морфофункциональная реакция Iba-I и MHC-II позитивных клеток селезенки на введение мелатонина
Введение. Мелатонин — нейропептид и нейротрансмиттер участвующий в синхронизации циркадных ритмов, оказывающий онкопротектор-ное [1], геропротекторное [3, 6] и иммуностимули рующее действие [3, 6, 9, 11]. Экстрапинеальный мелатонин играет ключевую роль в координации клеточных функций и межклеточных связей при иммунном ответе [11]. Одни исследователи считают, что мелатонин не оказывает ни прямого, ни опосредованного Т-лимфоцитами влияния на активированные макрофаги [13]. Результаты других исследований показывают, что количество натуральных киллеров и макрофагов в селезенке под влиянием мелатонина возрастает [8]. Существует ряд исследований, свидетельствующих о чувствительности иммунных клеток селезенки к условиям освещения, в которых находится организм [14] и, вероятно, реакция этих клеток на введение мелатонина будет так же зависеть от светового режима.
Белки главного комплекса гистосовместимости второго класса (MHC-II) необходимы для развития Т-клеточного иммунного ответа, а также для обеспечения взаимодействия между Т-лимфоцитами и макрофагами в процессе иммунного ответа [4], и следовательно, являются специфичными маркерами активированных макрофагов. Динамичность экспрессии молекул MHC-II зависит от функционального состояния клетки и изменения под влиянием различных стимулов. Это один из механизмов, контролирующих индукцию и реализацию иммунного ответа [5]. Ионизирующая кальций связывающая адапторная молекула (Iba-1) идентифицирована как кальций-связывающий белок, экспрессирующийся клетками моноцитарно-макрофагального происхождения [2]. Iba-1 принимает участие в реорганизации актинового скелета и образовании мембранных складок макрофагов при индуцированном фагоцитозе [10]. Связь между Iba-1 и MHC-II молекулами проявляется в процессах антигенпрезентации. Антигены захватываются макрофагальными цитоплазматическими отростками, в образовании которых участвует молекулы Iba-1, и после фрагментации связываются с молекулами MHC-II. Снижение экспрессии Iba-1 или MHC-II приводит к ослаблению иммунного ответа [2]. Следовательно, изменение количества Iba-1- и MHC-II-позитивных клеток является показателем силы иммунного ответа.
Цель исследования - выявить реакции Iba-1- и MHC-II-позитивных клеток селезенки на экзогенный мелатонин при разных световых режимах.
Материал и методы исследования. Объектом исследования служила селезенка 120 двухмесячных мышей-самцов одной массы (18 - 22 г), содержавшихся при сбалансированном рационе питания и в различных условиях освещения.
Животные были разделены на 4 группы:
I - (n = 30) – животные, которые содержались в обычных условиях вивария в течение 4 недель эксперимента (естественное освещение; продолжительность светового дня 8 – 9 часов; освещенность на уровне клеток в утренние часы 50 – 150 люкс, днем в пасмурный день – до 500 люкс, в ясный день – до 1000 люкс, вечером 100 – 200 люкс; влажность воздуха 40 – 60%, свободный доступ к питьевой воде и корму);
-
II - (n = 30) – животные получали препарат мелаксен ad libium в концентрации 4 мг/литр с питьевой водой в течение 4 недель и находились в условиях обычного освещения (естественное освещение; продолжительность светового дня 8 – 9 часов; освещенность на уровне клеток в утренние часы 50 – 150 люкс, днем в пасмурный день – до 500 люкс, в ясный день – до 1000 люкс, вечером 100 – 200 люкс; влажность воздуха 50 – 70%);
-
III - (n = 30) – животные находились в условиях постоянного затемнения (освещенность в клетках в течение дня составляла 0 – 0,5 люкс) в течение 4 недель и получали питьевую воду и корм без ограничений;
-
IV - (n = 30) – животные получали препарат мелаксен ad libium в концентрации 4 мг/литр с питьевой водой в течение 4 недель и находились в условиях постоянного затемнения (освещенность в клетках в течение дня составляла 0 – 0,5 люкс).
Селезенка у животных забиралась после декапитации на 28-е сутки эксперимента во второй половине дня (1500 – 1700). Все действия, предусматривавшие контакт с лабораторными животными, осуществлялись с учетом требований «Правил проведения работ с использованием экспериментальных животных» («Приказ МЗ РФ от 19.06.2003 г. №267) и в соответствии с «Европейской конвенцией о защите позвоночных животных, используемых для экспериментов или в иных научных целях».
Криостатные срезы селезенки толщиной 5 мкм обрабатывались непрямым иммунофлуорес-центным методом по стандартному протоколу: эндогенная пероксидазная активность подавля- лась путем инкубации срезов в 3% растворе H2O2 в течении 30 минут с последующей трехкратной промывкой 0,1 М фосфатным буфером. Блок неспецифического связывания проводился преинкубацией срезов с 10% козьей сывороткой и 0,05% тритоном Х-100. В качестве первичных антител были использованы анти-MHC класс II (1:4; rat anti rat RT1Bu, Class II polymorphic; Serotec, Germany) и анти-Iba-1 (1:400; rabbit polyclonal anti-Iba-1,Wako Chemicals GmbH, Neuss, Germany). Для визуализации MHC-II и Iba-1 использованы меченные флуорохромом вторичные антитела Alexa Fluor 488 (1:250; goat anti rat IgG; Invitrogen) [12]. Препараты анализировались при помощи светового микроскопа Микмед 5, подсчет структур проводился при увеличении 1500.
Результаты исследования и их обсуждение. Введение мелатонина в течение 4-х недель приводит к изменению морфометрических параметров селезенки: увеличивается средний вес селезенки от 0,19 ± 0,02 г в контрольной группе до 0,21 ± 0,05 г. (p = 0,01) во II-й экспериментальной группе и до 0,24 ± 0,03 (p = 0,004) в III-й экспериментальной группе. Увеличивается количество лимфоидных узелков от 4 ± 0,2 в контрольной группе до 15 ± 0,8 во II экспериментальной группе (p = 0,004) и до 17 ± 1,4 (p = 0,001) в III-й экспериментальной группе. Эти результаты определяются способностью мелатонина стимулировать пролиферацию лимфоцитов, которая усиливается у животных, находящихся в условиях затемнения.
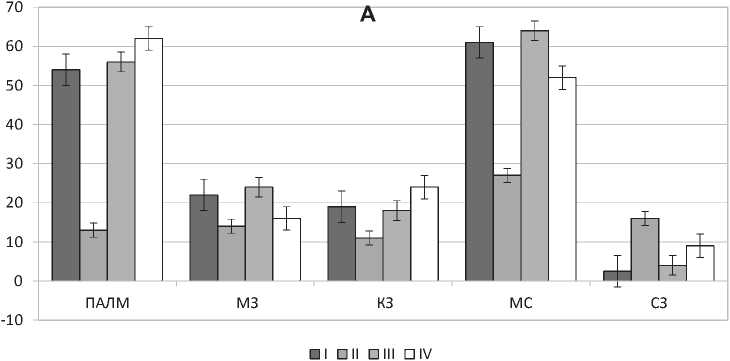
Исследование клеток моноцитарно-макрофагального происхождения показывает, что ежедневный введение мелатонина в течение 4 недель животным, находящимся в условиях естественного освещения приводит к уменьшению среднего количества Iba-1-позитивных клеток в 2 раза (p = 0,0006). Наблюдается уменьшение количества Iba-1-позитивных клеток в ПАЛМ и во всех зонах лимфоидных узелков. Исследование красной пульпы селезенки животных, получавших мелатонин, показало уменьшение количества Iba-1-позитивных клеток в маргинальных синусах и увеличение в субкапсулярных синусах красной пульпы (рис. 1).
Введение мелатонина в течение 4 недель животным, находящимся в условиях затемнения приводит к незначительному увеличению среднего количества Iba-1-позитивных клеток, отмечается увеличение количества исследуемых клеток в ПАЛМ, маргинальных зонах лимфоидных узелков и субкапсулярной зоне красной пульпы, что может свидетельствовать об усилении миграции макрофагов из кровотока в пульпу.
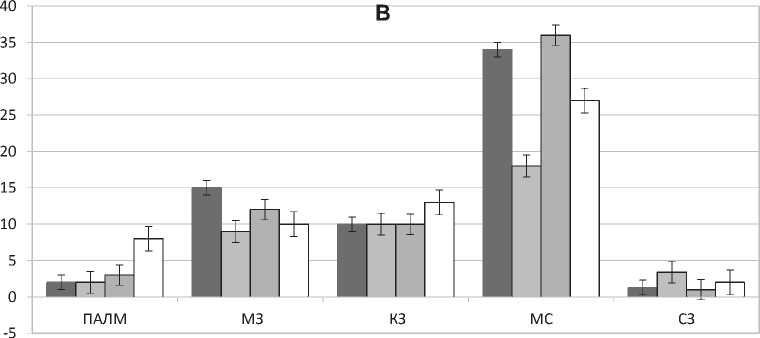
Изучение антигенпредставляющих клеток селезенки (MHC II-позитивных) показало, что на введение мелатонина в условиях естественного
■ I□ II □ III □ IV
Рис. 1. Количественная реакция Iba-1- (А) и MHC II-позитивных клеток (B) селезенки мышей на ежедневное введение мелатонина в течение 4-х недель. ПАЛМ — периартериальная лимфоидная муфта, МЗ — мантийная зона лимфоидного узелка, КЗ — маргинальная зона лимфоидного узелка, МС — маргинальный синус, СЗ — зона красной пульпы, расположенная под капсулой органа. I, II, III, IV — экспериментальные группы животных. Достоверность р <0,001 отмечается подчеркиванием числового значения, р <0,01 — двойным подчеркиванием.
освещения они реагируют подобно Iba-1-пози-тивным клеткам. Наблюдается снижение среднего количества клеток в поле зрения в 1,5 раза (p = 0,001), за исключением MHC II-позитивных клеток периартериолярных лимфоидных муфт и краевых зон лимфоидных узелков (рис. 1Б). Чувствительность макрофагов, дендритных и других антигенпрезентирующих клеток, вероятно, объясняется наличием мелатониновых рецепторов МТ3 [7], посредством которых осуществляется иммуномодулирующее действие. Введение мелатонина животным, находящимся в условиях затемнения не приводит к значимым изменениям среднего количества активированных макрофагов, но, можно отметить увеличение количества клеток в ПАЛМ, в маргинальных зонах лимфоидных узелков (рис. 2).
Мелатонин, вводимый перорально в течение четырех недель вызывает морфологические изменения в селезенке:
увеличение количества лимфоидных узелков, более выраженное в селезенке животных, содержавшихся в условиях затемнения;
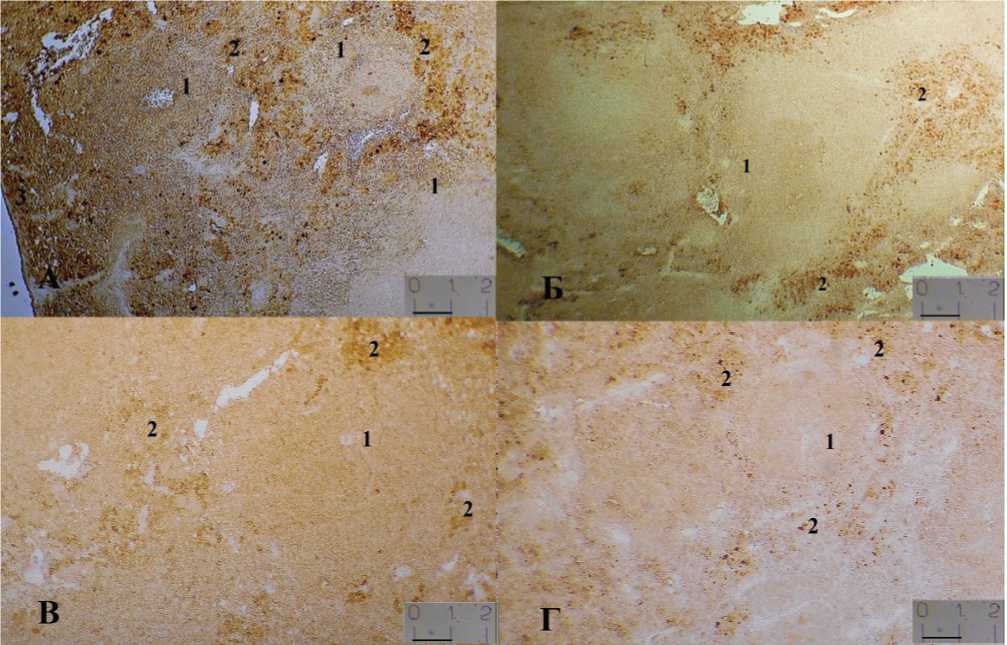
Рис. 2. Локализация MHC II позитивных клеток в селезенке мышей разных экспериментальных групп: А – III, Б – IV, В — I, Г — II; 1 - лимфоидный узелок, 2 - маргинальный синус. Иммуногистохимическая реакция с анти-MHC. Одно деление шкалы соответствует 22 мкм.
уменьшение среднего количества клеток макрофагального ряда (Iba-1+) и активированных макрофагов (MHCII+) в селезенке животных, содержавшихся при естественном освещении;
незначительные изменения среднего количества клеток макрофагального ряда (Iba -1+) и активированных макрофагов (MHCII+) в селезенке животных, содержавшихся при затемнении.
Наиболее чувствительными к световым условиям содержания животных, которым вводился мелатонин, являются периартериальные лимфоидные муфты, маргинальные и субкапсулярные синусы красной пульпы селезенки.
Список литературы Морфофункциональная реакция Iba-I и MHC-II позитивных клеток селезенки на введение мелатонина
- Арушанян, Э. Б. Гормон мозговой железы эпиф иза мелатонин - универсальный естественный адаптоген / Э. Б. Арушанян, Э. В. Бейер // Успехи физиологических наук. - 2012. - Т. 43. - № 3. - С. 82 - 100
- Кирик О.В., Сухорукова Е.Г, Коржевский Д.Э. Кальций-связывающий белок Iba-1/AIF1 в клетках головного мозга крысы // Морфология. 2010. Т. 136, № 2. С. 5 -7
- Князькин, И. В. Экстрапинеальный мелатонин в процессах ускоренного и преждевременного старения у крыс / И. В. Князькин // Успехи геронтопогии. - 2008. - Т. 21. - № 1. - С. 80 - 82
- Маянский H.A., Маянский А.Н. Номенклатура и функции главного комплекса гистосовместимости человека // Иммунология. 2006. Т. 27, № 1. С. 43-46
- Сарилова И.Л., Сергеева В.Е., Смородченко A.T. Эффект тестэктомии на структуры випочковой железы, экспрессирующие главный комплекс гистосовместимости II класса // Бюпп. Эксп. Биоп. Мед. 2008. Т. 146, № 1. С. 103 - 106
- Baeza, I. Improvement of leucocyte functions in ovariectomised aged rats after treatment with growth hormone, melatonin, oestrogens or phyto-oestrogens / I. Baeza, C. Alvarado, P. Alvarez, V. Salazar, C. Castillo, C. Ariznavarreta, J. A. Fdez-Tresguerres // J Reprod Immunol. - 2009. - V. 80. - № 1-2. - P 70 - 79
- Cernysiov, V. Expression of melatonin receptors in the cells of immune system / V. Cernysiov, R. Bozaite, M. Mauricas, I. Girkontaite // Immunology. - 2012. - V. 137. - № 1. - P 296
- Currier, N. L. Exogenous melatonin: quantitative enhancement in vivo of cells mediating non-specific immunity /N. L. Currier, L. Z. Sun, S. C. Miller // J Neuroimmunol. - 2000. V. 104. - № 2. - P 101 - 108
- Gupta, S. Photoperiodio regulation of nuclear melatonin receptor RORo in lymphoid organs of a tropical rodent Funambulus pennanti: role in seasonal oxidative stress [Электронный ресурс] / S. Gupta, С. Haldar, R. J. Ahmad // Photochem Photobiol B. 2014. - Режим доступа: http://www.doi: IO. 1016/j. Jphotobiol.2014.11.013
- Ohsawa K., Imai Y., Sasaki Y, Kohsaka S. Microglia / macrophage-specific protein Ibal binds to fimbrin and enhances its actin-binding activity // J. Neurochem. 2OO4. Vol. 88. P. 844 - 856
- Ozkanlar, S. Melatonin Modulates the Immune System Response and Inflammation in Diabetic Rats Experimentally-Induced by Alloxan [Электронный ресурс] / S. Ozkanlar, A. Kara, E. Sengul, N. Simsek
- A. Karadeniz, N. Kurt // Horm Metab Res. - 2O15. -V. 47. - P. 1 - 8
- Smorodchenko A.,Wuerfel I., Pohl E., Vogt I. et. al. CNS - irrelevant T-cells enter the brain, cause blood-brain barrier disruption but no glial pathology // Eur J. Neurosci. - 2OO7. V. 26. - № 6. - P. 1387-1398
- Shafer, I.E. Assessment of melatonin's ability to regulate cytokine production by macrophage and microglia cell types / L. L. Shafer, J. A. McNulty, M. R. Young // J Neuroimmunol. - 2OO1. - V. 120. - № 1-2. - P. 84 - 93
- Valdés-Tovar, M. Constant light suppresses production of Met-enkephalin-containing peptides in cultured splenic macrophages and impairs primary immune response in rats / M. Valdés-Tovar, С. Escobar, H. Solfs-Chagoyân, M. Asai, G. Benftez-King // Chronobiol Int. - 2O15. - V. 32. - № 2. - P. 164 - 177
