Морфофункциональные признаки мужского гаметофита томата в условиях вирусного патогенеза

Автор: Салтанович Татьяна Исаевна, Андроник Лариса Исааковна, Анточ Людмила Петровна, Дончилэ Анна Николаевна
Журнал: Овощи России @vegetables
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (58), 2021 года.
Бесплатный доступ
Актуальность. Действие вирусных фитопатогенов измененяет качество мужского гаметофита и в результате снижает продуктивность растений. В этой связи информация об особенностях вариабельности морфофункциональных признаков микрогаметофита приобретает особую актуальность. Учитывая экспрессию части генома растений в пыльце, мужской гаметофит может быть использован как надежная система оценки реакции генотипов на действие вирусных патогенов. Исходя из того, что вопросы вариабельности признаков мужского гаметофита в условиях вирусного патогенеза изучены недостаточно, не выявлены основные факторы, детерминирующие изменчивость, цель исследований состояла в изучении влияния вирусных фитопатогенов на морфофункциональные признаки мужского гаметофита томата. Материал и методы: В экспериментах использовали гибриды Fi и сорта томата. Растения выращивали в теплице, в фазе 4-5 листьев проводили их механическую инокуляцию вирусом табачной мозаики или вирусом аспермии томата. Для определения качества пыльцы собирали цветки контрольных и инфицированных растений, выделяли пыльцу и высевали на питательную среду. Анализировали препараты под микроскопом, определяли жизнеспособность пыльцы и длину пыльцевых трубок, вычисляли соотношение этих показателей. Результаты. Инфицирование растений томата вирусами приводило к изменениям функциональных признаков мужского гаметофита, которые, проявлялись как реакции стимуляции, ингибирования или нейтральные эффекты. Главными источниками вариабельности признаков пыльцы являлись генотип и вирусные агенты. В условиях вирусного патогенеза обнаружена неравноценность пыльцевых зерен по скорости прорастания и роста пыльцевых трубок (ПТ). Анализ значений коэффициентов наследуемости соотношения признаков жизнеспособности пыльцы и длины ПТ (опыт/контроль) обнаружил положительное доминирование и сверхдоминирование в 70% случаев. Выявлены генотипы сочетающие высокий уровень жизнеспособности пыльцы при инфицировании разными вирусами, что предполагает возможность их использования в дальнейших исследованиях.
Томат, вирус, пыльца, жизнеспособность, изменчивость, устойчивость, селекция
Короткий адрес: https://sciup.org/140257575
IDR: 140257575 | УДК: 635.64:581.16:632.38 | DOI: 10.18619/2072-9146-2021-2-16-21
Morphofunctional characteristics of the tomato male gametophyte under conditions of viral pathogenesis
Relevance. The action of viral phytopathogens changes the quality of the male gametophyte, which ultimately leads to a decrease in plant productivity. So, information on the features of the variability of morphofunctional characteristics of the microgametophyte becomes topical. Taking into account the expression of a part of the plant genome in pollen, the male gametophyte can be used as a reliable system for assessing the response of genotypes to the action of viral pathogens. Presuming that the problems of variability of characteristics of the male gametophyte in conditions of viral pathogenesis have not been enough studied, as well as the main factors determining the variability have not been identified, the aim of the research was to study the effect of viral phytopathogens on the male gametophyte characters in tomato. Material and methods. Hybrids Fi and varieties of tomato were used in the experiments. Plants were grown in a greenhouse. They were inoculated with tobacco mosaic virus (TMV) or tomato aspermia virus (TAV) in the 4-5 leaf phase. To determine the quality of pollen, flowers of control and infected plants were collected; pollen was isolated and sown on a nutrient medium. The preparations were analyzed under microscope. The viability of the pollen and the length of the pollen tubes were assessed, and the ratio of these indicators was calculated. Results. Infection of plants with viruses changes some functional characteristics of the male gametophyte. The responses manifested as stimulation, inhibition, or neutral effect. The main sources of variability in characters of pollen were the genotype and viral agents. In viral pathogenesis, unequal indicators of pollen grains were found in terms of the rate of germination and growth of pollen tubes (PT). Analysis of the heritability coefficients of the ratio of pollen viability and PT length (experiment / control) revealed positive dominance and overdominance in 70% of cases. Genotypes have been identified that combine a high level of viability when infected with different viruses. The prospect of their use in further research is proposed.
Текст научной статьи Морфофункциональные признаки мужского гаметофита томата в условиях вирусного патогенеза
Vegetable crops of Russia. 2021;(2):16-21. (In Russ.)
Несмотря на успехи классической селекции в создании генотипов, устойчивых к абиотическим и биотическим факторам, проблема комплексной устойчивости, в том числе и к наиболее распространенным заболеваниям, окончательно не решена. Это связано с генетической сложностью контроля признака устойчи- вости, эволюционными процессами в системе «патоген-хозяин», а также появлением новых биотипов патогенов [1]. Действие фитопатогенных организмов, вызывающих болезни растений, приводит к значительным экономическим потерям. Известно немало случаев, когда в отдельных экосистемах из-за сильного поражения зарегистрирован значительный процент гибели ценных культур, приводящий к эрозии самого вида. В качестве возбудителей болезней растений вирусы стоят на втором месте после грибов [2]. Вирусные заболевания относятся к наиболее вредоносной группе инфекций, которые приводят к ежегодным потерям урожая превышающим 25% уровень, что связано с отсутствием сравнительно эффективных мер борьбы с ними в полевых условиях, а также недостаточным уровнем устойчивости культур [3]. Проникновение и размножение вирусов в клетках восприимчивых хозяев приводит к изменениям метаболизма растений, нарушению нормального развития и функционирования отдельных тканей и органов в целом. Несмотря на то, что развитие болезни и защитные реакции протекают одновременно, вклад каждого из этих процессов в возникающие изменения значительно зависит от восприимчивости растения к вирусу [4]. Для преодоления негативных последствий таких инфекций приобретают актуальность исследования по изучению реакции генотипов на действие вирусных патогенов на основе использования как классических так и нетрадиционных (экспресс) методов, в том числе и методов гаметной селекции.
Учитывая то, что у многих видов растений более половины генома экспрессируется в пыльце, мужской гаметофит может быть использован как надежная система для оценки степени взаимодействия генотипа со стрессовыми факторами среды [5]. В процессе формирования пыльцы влияние биотических факторов, также как и абиотических, может может вызывать существенные различия в ее качестве [6]. По мнению некоторых исследователей качество пыльцевых зерен, является одним из определяющих факторов репродуктивной биологии, так как оно тесно связано со способностью растений формировать полноценные семена [7, 8]. В этой связи следует отметить, что в условиях действия биотических факторов информация о пыльцеобразующей способности растений и качестве мужского гаметофита приобретает особо важное значение. Так, показано, что гибриды томата, которые проявили генетическую устойчивость к вирусу табачной мозаики, обнаружили более низкую пыль-цеобразующую способность [9]. Инфицирование растений киноа (Chenopodium quinoa) вирусом Sowhane mosaic sobemovirus провоцировало появление ряда морфологических изменений пыльцевых зерен.Аналогичные изменения пыльцы были обнаружены и у других видов растений, инфицированных вирусами [10]. Следовательно, условия, в которых формируются репродуктивные органы, оказывают влияние на количество и качество генеративных клеток и впоследствии потомства. Вместе с тем, только результаты нескольких исследований показали возможность повышения жизнеспособности пыльцы в расщепляющихся поколениях на основе отбора высокофертильной пыльцы [11]. Считают, что для улучшения качества мужского гаметофита и повышения адаптивности репродуктивной системы к действию абиотических и биотических факторов важное значение имеют как тип, так и диапазон генетической изменчивости, которые связаны с жизнеспособностью пыльцы [12].
Необходимо отметить, что проведение анализа пыльцы для каждой сельскохозяйственной культуры имеет свои задачи, это может быть связано с вопросами адаптации генотипов, продуктивности, устойчивости к факторам среды или гибридной природой исследуемого объекта [13]. Для томата знания о характеристиках мужского гаметофита могут представлять интерес в связи с вопросами адаптации генотипа к факторам среды, а также устойчивости к абиотическим и биотическим стрессам.
Учитывая то, что вопросы вариабельности признаков мужского гаметофита в условиях вирусного патогенеза изучены недостаточно, а также не определены основные факторы, детерминирующие изменчивость, цель проведенных исследований состояла в изучении влияния некоторых вирусных фитопатогенов на изменчивость функциональных параметров мужского гаметофита гибридов F 1 и сортов томата.
Материал и методы исследований
В качестве объекта исследований использовали 8 гибридных комбинаций F 1 томата: Mary Gratefullу x Venet; Mary Gratefully x Flacară; Venet x Jacota; Venet x Anatolie; Flacara x Prestij; Flacara x Tomiș; Jacota x Prestij; Jacota x Mihaela, а также набор сортов: Mary Gratefully, Venet, Flacara, Jacota, Anatolie, Prestij, Tomiș, Mihaela и Rufina. Эксперименты проводили в условиях теплицы и в лаборатории. Растения выращивали рендомизированно по общепринятой для томатов методике, в фазе 4-5 листьев проводили их механическую инокуляцию вирусом табачной мозаики (ВТМ) или вирусом аспермии томата (ВАТ).
Для определения качества пыльцы собирали цветки с контрольных и инфицированных растений, отделяли и подсушивали пыльники, выделяли пыльцу и высевали ее на искусственную питательную среду, содержащую 15% водный раствор сахарозы и борную кислоту – 0,006%. Культивирование пыльцы осуществляли в термостате при оптимальном температурном режиме 26...28 ° C в течение 3-х часов. Анализировали препараты под микроскопом, определяли жизнеспособность пыльцы (ЖП) и длину пыльцевых трубок (ПТ) в контрольном и опытных вариантах, а также вычисляли соотношение этих показателей.
Статистическую обработку полученных результатов проводили с использованием программ Statgraphics Plus 5.0 и Exel 2013. Для выявления основных источников изменчивости признаков мужского гаметофита применяли метод многофакторного дисперсионного анализа.
Результаты и обсуждение
В результате проведенных исследований установлено, что инфицирование растений ВТМ и ВАТ вызывало изменение значений функциональных признаков мужского гаметофита как у гибридов F 1 , так и у сортов томата. На фоне вирусных инфекций было отмечено снижение значений жизнеспособности пыльцы в среднем на 9,5-10,4%, а также формирование пыльцевых трубок, длина которых была меньше по сравнению с контрольным вариантом на 20,4-27,2% (рис.1).
Jacota, Flacara x Tomis) и 3-х сортов (Flacara, Tomis, M ihaela) отмечено увеличение жизнеспособности пыльцы на 5,0-36,0% в зависимости от генотипа ( рис. 3А).
При анализе вариабельности размеров ПТ в опыте выявлено, более чем у 50% генотипов длина трубок уменьшилась на 5-20 делений окуляр-микрометра по сравнению с контролем, и только у 3-х гибридов и одного сорта значения длины трубок превышали показатели контроля (рис. 3Б).
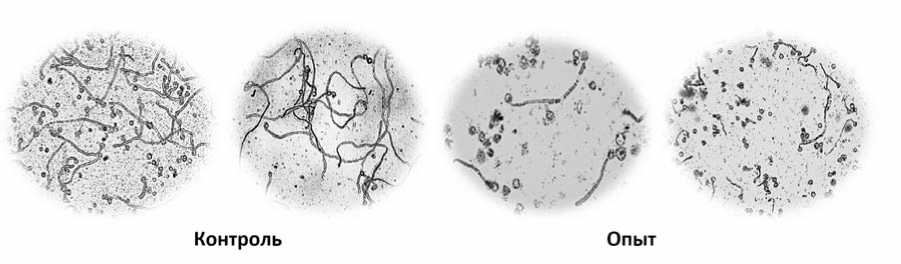
Рис.1.Влияние вирусной инфекции на изменение признаков мужского гаметофита томата
Fig.1.Influence ofthe viralinfection on changes in the characteristics ofthe tomato male gametophyte
Между опытными и контрольным вариантами обнаружено также наличие различий по частоте встречаемости пыльцевых зерен, способных к образовыванию пыльцевых трубок больших размеров. Так, если в контрольном варианте большинство пыльцевых зерен формировало трубки длиной 30-60 условных единиц, то в опытных вариантах (ВАТ и ВТМ), их длина была гораздо меньше – 10-30 условных единиц. Из общего числа изученных пыльцевых зерен в опыте только 2,518,0% имели длинные трубки. Следовательно, в результате действия вирусной инфекции лишь отдельные пыльцевые зерна способны формировать нормально развитые ПТ, что может определяться их неравноценностью по реакции на вирус (рис.2).
В результате анализа полученных данных установлено, что мужской гаметофит изученных сортов и гибридных комбинаций реагировал дифференцированно на действие вирусов. У большинства генотипов действие вируса ингибировало жизнеспособность пыльцы и только у 3-х генотипов не было обнаружено различий между опытным и контрольным вариантами. В то же время, на фоне ВМТ у 2-х гибридов (F 1 Venet x
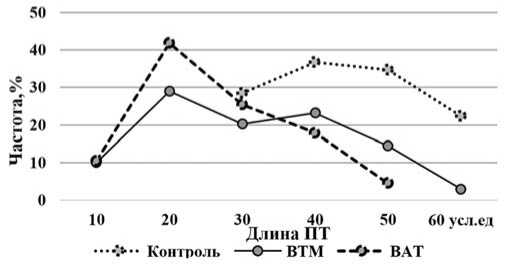
Рис. 2. Влияние ВТМ и ВАТ на распределение пыльцевых зерен по признаку длина пыльцевых трубок
Fig.2.Influence ofTMV andTAV on the distribution ofpollen grains based on pollen tube length
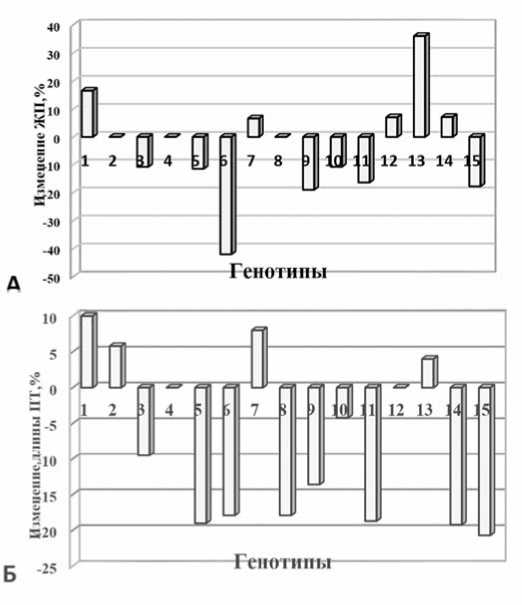
Рис.3. Влияние ВТМ на изменение жизнеспособности пыльцы (А)и длины ПТ(Б) томата:
1-8 – гибриды F1 VenetxJacota, Mary Gratefully xFlacara, Mary Gratefully x Anatolie, Mary Gratefully x Venet, Flacara x Prestij, Jacota xPrestij, Flacara x Tomis, Jacota x Mihaela; 9-15 – сорта Venet, Jacota, Mary Gratefully, Flacara, Tomis, Mihaela, Rufina Fig. 3. Influence of TMV on changes in tomato pollen viability (А)and PT (Б) length
У большинства растений, инфицированных ВАТ, жизнеспособность пыльцы была ниже контрольных значений на 7,0-40,0%, при этом только у 2-х гибридов и 2 сортов этот показатель возрастал. Среди изученных генотипов наиболее интенсивная реакция в сторону уменьшения жизнеспособности отмечена у гибридов
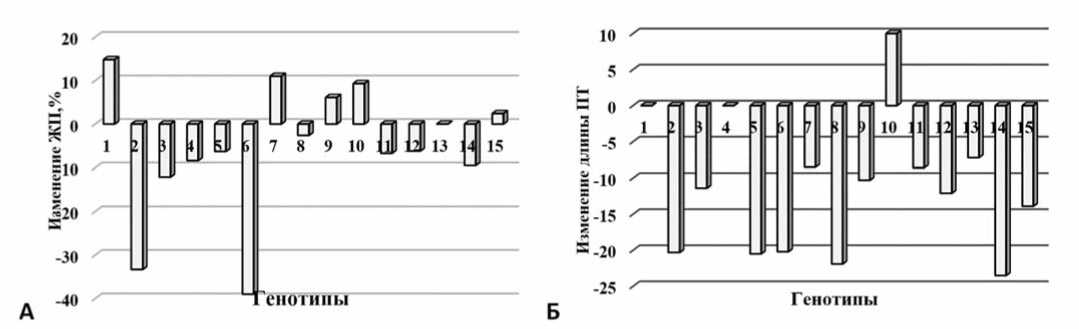
Рис.4.Влияние ВАТ на изменение жизнеспособности пыльцы (A) и длины ПТ(Б)томата:
1-8 – гибриды F 1 VenetxJacota, Mary Gratefully xFlacara, Mary Gratefully xAnatolie, Mary Gratefully xVenet, Flacara xPrestij, Jacota xPrestij, Flacara xTomis, Jacota xMihaela;9-15 – сорта Venet, Jacota, Mary Gratefully, Flacara, Tomis, Mihaela, Rufina.
Fig.4.Influence ofTAV on changes in tomato pollen viability (А)and PT (Б)length
F 1 Mary Gratefully x Flacara и Flacara x Prestij (рис. 4А). Действие ВАТ вызывало ингибирование процесса роста пыльцевых трубок, приводя к значительному уменьшению их размеров у 80,0% генотипов (рис. 4Б).
Таким образом, на основе обобщения полученных результатов установлено, что в зависимости от генотипа и варианта опыта (ВТМ или ВАТ) у пыльцевых зерен изученных генотипов наблюдаются разные реакции на действие вирусов: нейтральная, эффекты стимуляции или ингибирования. Так, в изученных вариантах частота проявления стимуляциционного эффекта составляла 6,7-27,0%, нейтральные реакции были выявлены в 13,0-20,0% случаев, тогда как в большинстве случаев наблюдалось ингибирование значений признаков – 53,3-80,0%. Следует отметить, что в литературе [14] известны случаи стимуляции жизнеспособности пыльцы под действием грибных патогенов. Так, при инфицировании растений томата грибом Verticillium dahlia также было обнаружено повышение жизнеспособности пыльцы у опытных растений. Исследователи полагают, что проявление стимуляционного эффекта, может быть связано с процессом селективной элиминации на этапах формирования пыльцевых зерен. В таком случае растение-хозяин является фоном для отбора
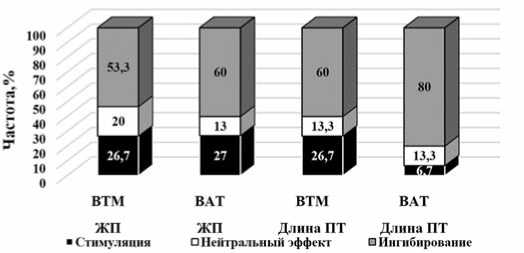
Рис.5.Реакция мужского гаметофита томата на действие ВТМ и ВАТ
Таким образом, мужской гаметофит некоторых генотипов томата весьма чувствителен к действию ВТМ и ВАТ, действие которых оказывает негативное влияние на активность пыльцевых зерен, что может снижать вероятность их участия в оплодотворении.
Табл.1. Источники изменчивости признаков мужского гаметофита томата
Table 1. Sources of variability of the characteristics of the tomato male gametophyte
Жизнеспособность пыльцы Длина пыльцевых трубок
|
Источники изменчивости (факторы) |
Степени свободы |
Сумма квадратов эффектов |
Доля влияния факторов, % |
Степени свободы |
Cумма квадратов эффектов |
Доля влияния факторов, % |
|
ВТМ |
||||||
|
Генотип |
14 |
21108,7* |
57,2 |
14 |
6709,2* |
22,2 |
|
ВТМ |
1 |
629,4* |
23,9 |
1 |
1477,9* |
68,4 |
|
Взаимодействие факторов |
14 |
6922,6* |
18,8 |
14 |
2387,4* |
7,9 |
|
Случайные эффекты |
60 |
151,7* |
5,1 |
60 |
1895,4* |
1,5 |
|
ВАТ |
||||||
|
Генотип |
14 |
21666,8* |
58,8 |
14 |
8613,9* |
16,9 |
|
ВТМ |
1 |
755,7* |
28,7 |
1 |
2783,3* |
76,5 |
|
Взаимодействие факторов |
14 |
4545,5* |
12,3 |
14 |
1915,1* |
3,8 |
* - Р < 0,05
Taбл. 2. Степень доминантности (h p ) признаков мужского гаметофита томата Table 2. Degree of dominance (h p ) of some characteristics of the tomato male gametophyte
|
Гибриды F 1 |
Степень доминантности, (h p ) |
|||
|
Жизнеспособность пыльцы (опыт / контроль) |
Длиа ПТ (опыт / контроль) |
|||
|
ВТМ |
ВАТ |
ВТМ |
ВАТ |
|
|
Venet x Jacota |
13,5 |
12,9 |
12,9 |
0,03 |
|
Mary Gratefully x Flacara |
0,53 |
0,50 |
0,50 |
-21,60 |
|
Mary Gratefully x Venet |
4,70 |
2,06 |
2,06 |
4,98 |
|
Flacara x Tomiș |
0,65 |
0,78 |
0,78 |
1,48 |
|
Jacota x Mihaela |
2,0 |
-1,38 |
-1,38 |
-1,21 |
Для изучение вклада генетических и средовых факторов в изменчивость функциональных признаков мужского гаметофита в условиях вирусного патогенеза использовали метод двухфакторного дисперсионного анализа. Согласно результатам статистической обработки генотип,вирусные патогены и их взаимодействие достоверно влияли на изменчивость признаков мужского гаметофита. Главным источником вариабельности жизнеспособности пыльцы как на фоне ВТМ, так и на фоне ВАТ был генотип, доля его влияния в общей изменчивости составляла 57,258,8%, сила влияния вирусов была более слабой и варьировала в пределах 23,9-28,7%, а взаимодействие этих факторов изменялось от 12,3 до 18,8%.
Несколько иная картина наблюдалась в случае выявления источников изменчивости длины пыльцевых трубок. Действие вирусов вносило существенный вклад – 68,4-76,6% в вариабельность их размеров, доля влияния генотипа была значительно ниже и составляла 16,9-22,2%, а взаимодействие этих факторов было равно 3,8-7,9% (табл.1). Выявленный довольно высокий вклад генотипа в изменчивости жизнеспособности пыльцы предполагает возмож- ность эффективного отбора генотипов по этому признаку.
Для выявления особенностей наследуемости признаков мужского гаметофита была вычислена степень доминантности. Значения этого показателя варьировали от отрицательного сверхдоминирования до положительного сверхдоминироваия. Отрицательное свехдо-минирование наблюдали в 20% случаев, промежуточное наследование отмечено в 10% случаев, положительное доминирование – 15% и положительное сверхдоминирование – 55% случаев (табл.2). Таким образом, положительное доминирование и сверхдоминирование было отмечено в 70% случаев, что может указывать на преобладание особенностей родителя с более высокими значениями признака.
На основе обобщения полученных данных по оценке признаков мужского гаметофита у контрольных и инфицированных растений, а также результатов дисперсионного анализа, мы проанализировали значения жизнеспособности пыльцы и длины пыльцевых трубок на всех фонах, что может дать представление об адаптивности генотипов в этих условиях. Среди изученных генотипов на фоне ВТМ наиболее высокий уровень
Табл. 3. Характеристика генотипов томата по признакам мужского гаметофита Table 3. Characteristics of tomato genotypes according to the characteristics of male gametophyte
|
Генотипы |
ВТМ |
ВАТ |
||
|
Жизнеспособность пыльцы, % |
Длина ПТ, (усл.ед.) |
Жизнеспособность пыльцы,% |
Длина ПТ, (усл. ед.) |
|
|
Гибриды F 1 |
||||
|
Venet x Jacota |
33,7 |
25,4 |
32,8 |
21,3 |
|
Mary Gratefully x Flacara |
70,6 |
42,8 |
53,5 |
45,1 |
|
Mary Gratefully x Anatolie |
45,9 |
30,7 |
45,3 |
29,7 |
|
Mary Gratefully x Venet |
75,1 |
42,1 |
70,3 |
43,1 |
|
Flacara x Prestij |
58,9 |
38,3 |
61,5 |
37,8 |
|
Jacota x Prestij |
66,4 |
32,4 |
67,9 |
31,2 |
|
Flacara x Tomis |
72,2 |
43,8 |
74,3 |
37,9 |
|
Jacota x Mihaela |
25,3 |
23,8 |
24,9 |
21,7 |
|
Сорта |
||||
|
Venet |
64,7 |
56,4 |
77,3 |
58,6 |
|
Jacota |
39,4 |
33,5 |
53,9 |
40,4 |
|
Mary Gratefully |
32,7 |
23,7 |
37,5 |
28,6 |
|
Flacara |
54,6 |
36,2 |
48,1 |
30,9 |
|
Tomis |
59,3 |
33,4 |
40,8 |
28,2 |
|
Mihaela |
40,2 |
30,8 |
38,9 |
28,7 |
|
Rufina |
55,3 |
43,6 |
63,2 |
46,9 |
|
НСР0,5 |
2,07 |
7,32 |
2,12 |
2,26 |
жизнеспособности пыльцы обнаружили 5 гибридных комбинаций F 1 (Mary Gratefully x Flacara, Mary Gratefully x Venet, Jacota x Prestij, Flacara x Prestij, Flacara x Tomis), а также 4 сорта (Venet, Jacota, Tomis, Rufina). Следует отметить, что большая часть вышеназванных генотипов сочетала высокий уровень жизнеспособности пыльцы со способностью формировать длинные пыльцевые трубки.
В то же время из числа генотипов инфицированных ВАТ выделены 5 гибридов F 1 (Mary Gratefully x Venet, Mary Gratefully x Flacara, Flacara x Prestij, Jacota x Prestij, Flacara x Tomis), а также 3 сорта (Venet, Jacota, Rufina) с высокими значениями изученных признаков. Таким образом, сопоставляя полученные результаты, можно заключить, что на фоне инфицирования ВТМ и ВАТ хорошие показатели качества мужского гаметофита проявили 5 гибридных комбинаций и 2 сорта, которые могут быть включены в дальнейшие исследования.
Выводы
-
• Инфицирование растений томата ВТМ и ВАТ приводит к изменениям признаков мужского гаметофита, которые, в зависимости от генотипа и вируса, проявляются как нейтральные реакции, эффекты стимуля-
- ции или ингибирования жизнеспособности пыльцы и длины пыльцевых трубок. Выявленная изменчивость в большей степени детерминируется генотипом и вирусными агентами.
-
• Анализ гистограмм распределения пыльцевых зерен по длине пыльцевых трубок выявил в инфицированных вариантах уменьшение частоты образования пыльцевых трубок больших размеров, что может свидетельствовать о гетерогенности популяции гамет по скорости их роста в условиях вирусного патогенеза.
-
• Значения коэффициентов наследуемости соотношения признаков жизнеспособности пыльцы и длины ПТ (опыт/контроль) варьируют от отрицательного сверхдоминирования до положительного сверхдоминирования, при этом положительное доминирование и сверхдоминирование отмечено в 70% случаев, что указывает на преобладание особенностей родителя с более высокими значениями признака.
-
• Мужской гаметофит гибридов F 1 Mary Gratefully x Venet, Mary Gratefully x Flacara, Flacara x Prestij, Jacota x Prestij, Flacara x Tomis и сортов Venet, Rufina сочетал высокую жизнеспособность при инфицировании ВМТ и ВАТ, что предполагает перспективу их использования в дальнейших исследованиях.
Об авторах:
Людмила Петровна Анточ – научный сотрудник
Анна Николаевна Дончилэ – младший научный сотрудник
Aboutthe authors:
Larisa I. Andronic – Doc. Sci. (Biology), director of the institute
Liudmila P. Antosh – Researcher
Anna N. Doncila – Trainee Scientific Researcher
(1997)078[1632:HEFAPP];2.
Vegetable crops of Russia. 2019;(4):32-36. (In Russ.)
Список литературы Морфофункциональные признаки мужского гаметофита томата в условиях вирусного патогенеза
- Халилуев М.Р. Шпаковский Г.В. Генно-инженерные стратегии повышения устойчивости томата к грибным и бактериальным патогенам. Физиология растений. 2013;60(6):721-732. https://doi.org/10.7868/S0015330313050084
- Котельникова И.М., Крылов A.B. Липиды при вирусном заражении растений// Вестник ДВО. 2001;(4):38-55.
- Чанг Нгуен Ха Тхи Куинь. Распространение и патогенез вирусных заболеваний томата в условиях Вьетнама и России. Москва, 2013.
- Реунов А. В. Вирусный патогенез и защитные механизмы растений. Владивосток: Дальнаука. 1999.
- Johnson M., Harper J., Palanivelu R. A. A Fruitful journey: Pollen tube navigation from germination to fertilization. Annual Review of Plant biology. 2019;(70):809-837. https://doi.org/10.1146/annurev-arplant-050718-100133
- Delph L.F., M.H. Johannsson, A.G. Stephenson. 1997. How environmental factors affect pollen performance ecological and evolutionary perspectives. Ecol. 1997;78(6):1632-1639. https://doi.org/10.1890/0012-9658(1997)078[1632:HEFAPP]2.0.CO;2.
- Круглова Н.Н. Оценка качества пыльцевых зерен цветковых растений. Бюллетень ГНБС. 2020;(135):50-56. https://doi.org/10.36305/0513-1634-2020-135-50-56.
- Беспалова Л.А., Агаев Р.А., Агаева Е.В. Морфологические особенности пыльцы у сортов пшеницы и тритикале селекции НЦЗ. Научный журнал КубГАУ. 2020;3(157). https://doi.org/10.21515/1990-4665-157-024
- Король В.Г. Опыление и плодообразование у культуры томата в защищенном грунте. Овощи России. 2019;(3):32-36. https://doi.org/10.18619/2072-9146-2019-4-32-36.
- Haight F., Gibbs A. Effect of viruses on pollen morphology. Plant Pathology. 2007;32(4):369-372. https://doi.org/org/10.1111/j.1365-3059.1983.tb02849.x.
- Razzaq M., Rauf F., Khurshid M. et al. Pollen viability an index of abiotic stresses tolerance and methods for the improved pollen viability. Pakistan jurnalof agricultural reserch. 2019;32(4):609-624. https://doi.org/10.17582/journal.pjar/2019/32.4.609.624
- Kalyar T., Rauf S., Teixeira S.J.A. Handling sunflower (Helianthus annuus L.) populations under heat stress. Arch. Agron. Soil Sci. 2014;(60):655- 672. https://doi.org/10.1080/03650340.2013.799276
- Цаценко Л.В., Назаров А.Л., Леленева А.Р. Пыльцевой анализ в селекции пшеницы. Научный журнал КубГАУ. 2021;165(01). http://ej.kubagro.ru/2021/01/pdf/09.pdf.
- Балашова Н.Н. Действие инфекционных фонов на репродуктивную систему и потомство зараженных растений (на примере культуры томатов). 1989. С.16-27.
