Морфогенез листа листостебельного мха Physcomitrium pyriforme

Автор: Донсков Дмитрий Геннадиевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Материалы лекций и устных докладов
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
Представлен краткий и обобщенный обзор литературных данных по изучению морфогенеза листа у мхов. Впервые проведен анализ и частичная реконструкция развития листа Physcomitrium pyriforme (Hedw.) Hampe. Выявлены и охарактеризованы особенности морфогенеза листа данного вида.
Короткий адрес: https://sciup.org/146116212
IDR: 146116212 | УДК: 581.452
Текст научной статьи Морфогенез листа листостебельного мха Physcomitrium pyriforme
Морфогенез листа у мхов, как предмет исследования, до сих пор остается мало изученным. Сделанных работ по данной тематике было немного, и все они были проведены уже довольно давно.
Сразу следует упомянуть, что по сравнению с листьями сосудистых растений листья мхов состоят из меньшего числа клеток, и, как правило, проще устроены. Пластинка листа у большинства видов однослойная, реже дву- или многослойная. Жилка всегда многослойная. Часто большее число слоев имеется по краю листа; клетки края листа иногда значительно отличаются от клеток средней части пластинки и образуют кайму [1; 2; 3].
В истории исследования первые представления о развитии листьев у мхов высказал Hornschuch (1821). Он считал, что стебли листостебельных мхов и их листья должны возникать посредством «прикрепления друг к другу нитчатых волокон и их ветвления», под которыми понимается протонема. Morren (1841) же понял, что ни через какие «нитчатые волокна» листья образовываться не могут. Он считал, что клетки должны возникать как «сгусток», ему удалось получить данные по изменению формы клеток. Nageli (1845, 1846, 1855) обнаружил закономерности в расположении клеток зачаточной части растения. Он нашел, что рост верхушечной клетки у листьев идет вразрез с точкой зрения Morren относительно роста клеток и выступил вместе с тем против воззрения Schleiden (1843, 1850), согласно которому лист у листостебельных мхов обладает базальным ростом, т.е. лист вырастает из стебля.
Обширные сведения о процессе деления клетки при росте листа и стебля были получены: Lorentz (1864), Leitgeb (1868, 1869, 1874), Berggren (1867 – 1868, 1870 – 1871), Kühn (1874), Débat (1876 – 1877), Goebel (1892), Morin (1893) и Lorch (1894, 1909). Lorentz (1867) создал первую, до сих пор еще широко применяемую, терминологию [5].
Эти исследования завершены публикациями Pottier [7]. В его первой работе излагаются основные моменты по объяснению истории развития листа. Во второй работе Pottier выступил с доказательствами, прежде всего того, что лист нарастает через деятельность одной верхушечной (апикальной) клетки [7].
Следующий важный шаг в исследовании морфогенеза листьев мхов сделал Frey [5; 6], который обобщил и систематизировал ранее накопленный материал, а также подробно рассмотрел начальные этапы образования листа у некоторых видов зеленых мхов. А главное, он ввел довольно удобные обозначения, которые мы и будем использовать в работе [5; 6].
Он установил, что верхушечная клетка стебля производит через периклинальное деление клетку, которая затем делится поперечно-антиклинальной стенкой, располагающейся примерно перпендикулярно к оси стебля. Получаются две клетки – I и II. Клетка I еще раз делится и отчленяет вниз клетку I1, и в результате образуется клетка I2, оказывающаяся в верхушечном положении. Из совокупности клеток II, посредством антиклинальных и периклинальных делений, возникают внутренние ткани. Клетки I1 участвуют в образовании поверхности, а также покоящихся почек, антеридиальных волосков и антеридиев (рис. 1).
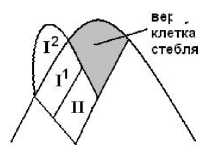
верхуш. клетка стебля
Рис. 1. Схема расположения клеток в верхушке стебля
Клетка I2 это и есть собственно апикальная клетка листа, от которой идет все дальнейшее построение листовой пластинки. Поэтому деятельность ее самой и ее производных является главным объектом в наших исследованиях. Деятельность остальных обозначенных выше клеток безусловно интересна, но лежит за гранью данной работы.
В отличие от других высших растений, образование клеток у мхов обычно является результатом ряда правильных делений. Такая регулярность почти всегда обнаруживается, по крайней мере, для некоторого числа делений. Поэтому, чтобы понять порядок образования клеток и развитие у Bryophytes, нужно рассмотреть последовательность этих делений.
Апикальная клетка I2 производит дочерние клетки в две стороны. Эти клетки и являются отправными точками для образования плоской структуры, которая будет закончена дальнейшими делениями.
Апикальная клетка отделяет через косые антиклинальные деления первое поколение потомков, от которых берет начало основание листа. Frey обозначил их как I31 и I32. Затем апикальная клетка последовательно производит клетки I41 и I42, I51 и I52, I61 и I62, I71 и I72, которые дают начало собственно пластинке. На этом деятельность апикальной клетки может приостановится, но иногда она еще откладывает одну или две пары клеток (рис. 2, a, b, c).
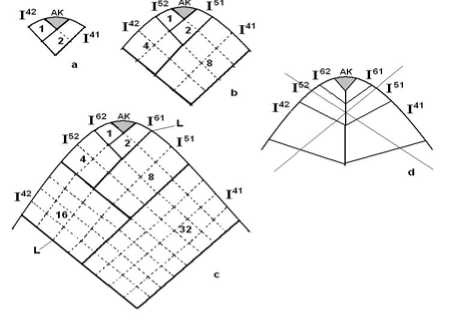
Рис. 2. Схема последовательности образования клеточной сети
-
a , b, c – идеальная последовательность; d - неосуществимый случай
Сформировавшийся вид клеточной сети листа – это следствие деления и удлинения клеток, происходящих гармонично. Деления не ограничены апикальной клеткой, что является общим правилом. Результат последовательных, правильных делений – равное распределение дочерних клеток по сторонам апикальной клетки и формирование двух симметричных половин. Таким образом, клетки получают начальное положение относительно главной оси листа, что предопределяет их последующие шаги деления.
После первого деления потомков происходит второе деление, перпендикулярное к первому, формируются квадранты и октанты однородных клеток с последующим делением и удлинением, в котором самое большое удлинение имеет место в недавно сформированной клеточной стенке. Следующее разделение тогда перпендикулярно длинной стороне, и т. д. Это означает, что между двумя делениями направление всегда изменяется приблизительно на 90 ° (рис. 2).
Окончательная форма листа есть последствие не только делений, но также и определенного удлинения клеточных стенок между делениями и, в большей степени, последующего роста растяжением, который происходит после того, как деления прекратились и конечное число клеток было достигнуто [4].
Основные удлинения клеточных стенок предопределены последним делением клетки. Новые клеточные стенки соседних клеток не выравниваются непосредственно друг с другом (чтобы формировать крестообразное пересечение и решетчатую структуру), но, как правило, немного перемещаются, формируя вид двойного Y. Каждая клетка поэтому окружена шестью соседними клетками, четыре из них с короткой общей стенкой и две с длинной. Этот тип образования может быть понят, если ось смещения всегда немного наклонена от длинной оси делящейся клетки [3; 4].
Материалы и методы. Объект исследования – верхоплодный мох Physcomitrium pyriforme (Hedw.) Hampe.
Данный объект выбран в связи с тем, что:
-
- подобные исследования на нем не проводились,
-
- он относительно легко доступен в живом виде, более подходящем для данного исследования,
-
- его листья построены из небольшого числа крупных клеток.
Изучение строения и развития листа проводилось путем изготовления микропрепаратов. Препараты готовились с живых или засушенных образцов, на предметных стеклах, и заливались в среду мовиол. Живые образцы собирались на территории Главного ботанического сада РАН им. В.Н.Цицина (ГБС). Засушенные образцы взяты из гербария ГБС.
Для изготовления препаратов использовались верхушечные, молодые листья побегов, на различных стадиях развития. Затем препараты фотографировались с помощью цифрового зеркального фотоаппарата Nikon D70s, закрепленного на микроскопе. Полученные фотографии просматривались на компьютере в программе Nikon Capture Editor. Всего изготовлено 120 препаратов, с них сделано около 350 фотоснимков.
Далее полученные фотографии упорядоченно выстраивались по степени развития листьев и давалась реконструкция развития, в значительной степени основывающаяся на работах Frey [5].
Результаты. Апикальная клетка листа имеет более или менее ромбовидную форму. По серии фотографий видно, что она совершает косые деления, последовательно откладывая клетки в обе стороны.
Эти клетки, потомки апикальной, практически сразу начинают делится. Отчетливо просматриваются сформировавшиеся при этом квадранты, октанты и т. д.
новых клеток в последовательности 2, 4, 8 … (далее эти группы клеток будем именовать сектора , присваивая им те же обозначения I41 и I42, I51 и I52 и т. д.) В дальнейшем этот численный ряд нарушается, и число клеток в секторе не строго соответствует количеству 16, 32, 64.
В молодом листе на самой верхушке побега клетки мелкие и однородные (рис. 3). В листьях среднего возраста, располагающихся чуть ниже и активно растущих, клетки вверху листа сильно увеличиваются в размерах (рис. 4). В нижней половине они по-прежнему остаются мелкими. Во взрослом, полностью сформированном листе, все клетки довольно крупные и их число колеблется от 200 до 500 (рис. 5). Ширина половины плоскости взрослого листа – 7 – 16 клеток.
У Physcomitrium pyriforme листья, расположенные внизу стебля, мелкие, но при этом образованы крупными клетками (рис. 6). Число последних может достигать всего лишь 25 – 30.
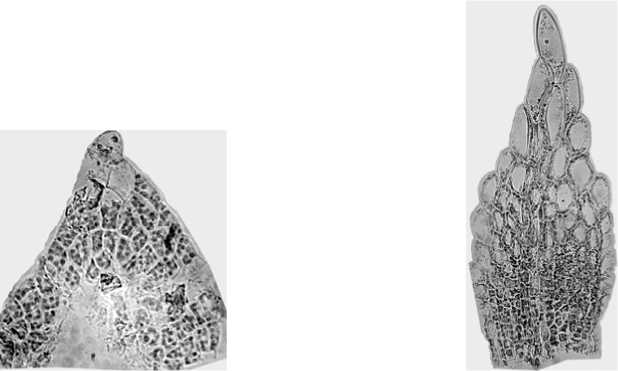
Рис. 3. Молодой лист
Рис. 4. Лист среднего возраста
Границы секторов отчетливо просматриваются у молодых листьев и у взрослых мелких листьев с нижней части стебля. На средней стадии развития листа картина бывает различной. Во взрослом листе выделить границы с ходу довольно трудно, но последовательно перебирая различные варианты все же их можно установить с большой долей вероятности.
Зато во взрослой стадии листа отчетливо становятся видны продольные ряды клеток, тянущиеся от верхушки к основанию листа. Причем, что характерно, ряды расположены по принципу «кирпичной кладки», при которой они смещены относительно друг друга.
Особое внимание заслуживает хорошо заметная клетка в верхушке листа Physcomitrium pyriforme , расположенная под апикальной клеткой, по главной оси листа. Назовем ее субапикальной . Она имеет более-менее ромбовидную форму, как правило, немного больше остальных клеток, почти всегда выпуклая и располагается вне плоскости листа (рис. 5).
Так же у этого вида в верхушке листа отчетливо видно, как клетки края заворачиваются вовнутрь, немного налегая на соседние клетки. При этом они часто сильно изогнуты, причем с одной из сторон намного сильнее. Это приводит к тому, что верхушка листа выглядит более острой (рис. 5).
У мелких взрослых листьев, расположенных в нижней части стебля, жилка не образуется, и лист полностью однослойный. У листьев, которые впоследствии станут крупными, наоборот, жилка образуется довольно рано. Ее хорошо видно уже на очень ранних стадиях. Жилка строится из узких, продольных клеток, резко отличных от клеток плоскости листа. Ее появление вносит дополнительную неупорядоченность в границы секторов, и становится трудно детально прорисовать, где по главной оси сектора соприкасаются. По длине жилка сильно варьирует – она может достигать только до середины листа, или оканчиваться в верхушке. При этом вверху она подходит к субапикальной клетке.

Рис. 5. Взрослый лист
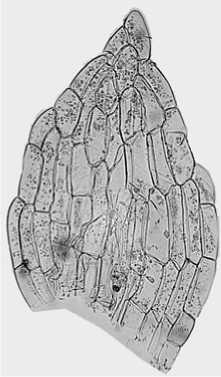
Рис. 6. Взрослый листа, с нижней части стебля
Обсуждение. На фотоснимках видно, что апикальная клетка листа имеет форму близкую к ромбовидной, и это вполне закономерно, так как именно при такой форме возможно равномерное отделение клеток в обе стороны, с созданием первоначальной плоскости листа.
В своих работах Pottier, Frey, Bopp указывают на небольшое количество делений апикальной клетки. Это подтверждают и мои исследования на Physcomitrium pyriforme . У маленьких нижних листьев верхушечная клетка от 4-х до 6-ти делений, у больших – от 6 до 10. После этого ее деления полностью прекращаются и плоскость листа образуется посредством делений этих клеток, производных от апикальной. Таким образом, наглядно видно, что весь лист строится из потомков довольно малого числа клеток.
Потомки апикальной клетки быстро переходят к делениям. Причем на первых этапах их деления хорошо коррелируют с делением верхушечной клетки и близлежащих секторов. Это отчетливо иллюстрируют стадии секторов состоящие из 2, 4, 8 клеток. В дальнейшем, с ростом секторов, синхронизация делений в соседних секторах уменьшается, количество клеток получается разное и редко соответствует правильному – 16, 32, 64.
Последующие размытие границ секторов во взрослом листе объясняется продольным вытягиванием клеток. Структура «кирпичная кладка» скорее всего получается при удлинении новой продольной клеточной стенки, образовавшейся вследствии поперечного деления двух соседних клеток. Это легко объяснить, если принять, что поперечные стенки образуются со смещением относительно друг друга (рис. 7). Образование такой структуры оправданно с точки зрения прочности листа, как конструкции, состоящей из многих блоков. Надо помнить, что пластинка листа большинства мхов, и в частности Physcomitrium pyriforme, однослойная и не может быть укреплена сверху или снизу. Вариант со смещением – один из немногих, способных ее упрочить.
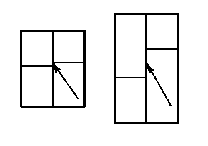
Рис. 7. Схема образования структуры «кирпичная кладка»
Особый вопрос, на котором хотелось бы заострить внимание – выделение секторов в листе. Идеальная картина построения клеточной сети представлена на рис. 2,с, когда все деления абсолютно правильны и скоррелированы друг с другом. В реальности такого, конечно, не бывает, однако эта схема помогает сформулировать некоторые критерии выделения секторов:
-
1) учет общих особенностей деления растительной клетки, не позволяющих смещаться им друг относительно друга;
-
2) верхняя, внутренняя граница нижележащего сектора должна соприкасаться с нижними границами двух верхних секторов. Т.е. случай представленный на рис. 2, d невозможен;
-
3) продолжение линии, обозначающей верхнюю, внутреннюю границу верхнего сектора, оказывается линией первого деления в нижележащем секторе; данное правило представляется наиболее логичным и согласующимся с полученными картинами клеточных сетей; вместе с тем возможность альтернативных правил не исключена.
Пример – линия верхней внутренней границы сектора I51 соприкосается с секторами I61 и I52, и продолжается в секторе I42 (линия L) (рис. 2,с).
Уже эти три критерия позволяют нам создать первоначальную реконструкцию развития листа. Сравним рис. 8 и 9. Реконструкция, представленная на первом, выполнена по введенным нами правилам. Линии 0 отмечают границы секторов, линии 1, 2, 3 и т. д. показывают последовательность делений клеток в секторах. Реконструкция на втором рисунке не может быть осуществлена до конца, т. к. верхняя, внутренняя граница сектора I52 не имеет продолжения в нижнем секторе I51 (линия L).
Конечно, развитие листа в реальности далеко от идеала. Как уже упоминалось количество клеток в больших секторах не соответствует ряду 16, 32, 64. Отдельные клетки могут совершать дополнительные деления, не коррелирующие с делением соседних клеток. Или наоборот, вместо логичного деления предпочитают более сильное растяжение.
Учитывая вышеприведенные критерии, построим реконструкции листьев разного возраста – рис. 8, 10, 11, 12; линии 0 отмечают границы секторов, линии 1, 2, 3 и т. д. – показывают последовательность делений клеток в секторах.
Еще один важный вопрос – образование жилки. Она образуется при усиленном делении внутренних клеток секторов, близких к оси листа, с формированием многослойной структуры. Данные клетки оказываются зажаты между краевыми клетками, и не могут делится в плоскости листа до бесконечности, т. к. основание листа имеет конечную ширину, жестко связанную с диаметром стебля.
Поэтому их единственный вариант – делится перпендикулярно плоскости с образованием многослойности.
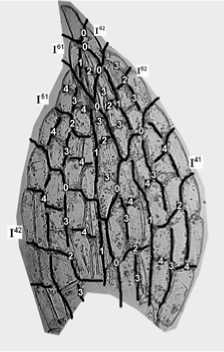
Рис. 8. Реконструкция последовательности делений клеток во взрослом листе, с нижней части стебля
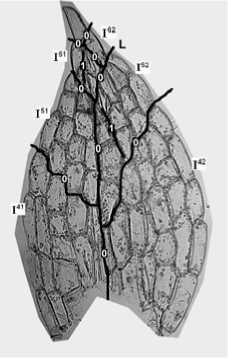
Рис. 9. Вариант реконструкции последовательности делений клеток в листе, выполненный не по введенным правилам
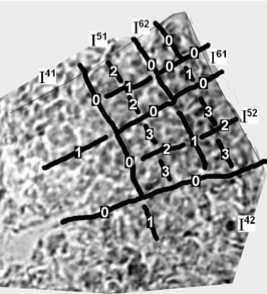
Рис. 10. Реконструкция последовательности делений клеток в молодом листе

Рис. 11. Реконструкция последовательности делений клеток в листе среднего возраста
Характерно, что взрослые мелкие листы, расположенные внизу стебля, не имеют жилки. Это можно объяснить тем, что клеток в основании мало, диаметр стебля достаточно велик, а следовательно, осевые клетки не испытывают напряжения с боков и могут дифференцироваться стандартно, как и соседние клетки.
В настоящий момент еще трудно сказать, какие именно клетки идут на образование жилки. Также сложно обозначить ее связь с секторами. Ее появление вносит дополнительную неупорядоченность в границы секторов, и не всегда возможно детально прорисовать, где по главной оси сектора соприкасаются.
Особенность верхушки листа Physcomitrium pyriforme , выраженную в том, что клетки края заворачиваются вовнутрь, налегая на соседние клетки, можно объяснить с точки зрения дополнительного укрепления конструкции. Такой изгиб клеток влечет за собой вогнутость листа и создает дополнительные ребра жесткости, сходящиеся к уже имеющемуся, а именно – жилки. Причем заворачивание клеток имеет место и у нижних листьев, у которых нет жилки. В результате этого весь лист становится достаточно прочным к внешним воздействиям.
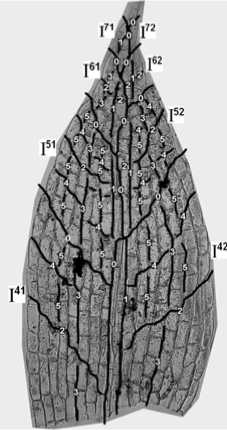
Рис. 12. Реконструкция последовательности делений клеток во взрослом листе
Клетка, названная мною субапикальной, у Physcomitrium pyriforme занимает уникальное положение и, вполне возможно, сильно влияет на все развитие листа. Однако этот вопрос требует дальнейшего изучения.
В заключение можно сделать выводы:
-
1. Развитие листа Physcomitrium pyriforme происходт в соотвествии с закономерностями, описанными Фреем;
-
2. Апикальная клетка делится от 6 до 8 раз, отделяя в две стороны по 3 или 4 клетки, которые и формируют лист;
-
3. Число делений клеток I41 и I42, строящих нижние сектора листа также обычно 8, в недоразвитых листьях 6, в верхних секторах число делений последовательно уменьшается;
-
4. В верхних секторах листа число и расположение клеток приближается к теоретически рассчитываемому согласно схеме на рис. 2; в нижних секторах картина нарушается в связи с образованием жилки;
-
5. Реконструкция листа Physcomitrium pyriforme на основе 3 приведенных критериев, как правило, дает непротиворечивую и однозначную последовательность клеточных делений.
An overview of literature data of leaf morphogenesis in mosses is given. Leaf development in Physcomitrium pyriforme (Hedw.) Hampe was reconstructed as a sequence of cell divisions and principles of this reconstruction are discussed.
