Морфологическая гетерогенность сахалинского подкаменщика Cottus amblystomopsis Schmidt, 1904 островов Сахалин, Кунашир, Итуруп и Хоккайдо

Автор: Жидков З.В., Павлов И.А.
Журнал: Природные системы и ресурсы @ns-jvolsu
Рубрика: Экология и охрана окружающей среды
Статья в выпуске: 1 т.15, 2025 года.
Бесплатный доступ
Статья посвящена изучению внутривидовой изменчивости сахалинского подкаменщика, Cottus amblystomopsis (Cottidae). Этот вид описан из рек Южного Сахалина и обнаруживается на островах южной части Курильской гряды, а также на Хоккайдо. Морфологическое исследование сахалинского подкаменщика по всему ареалу до сих пор не проводилось. Для определения морфологических различий между разными островными популяциями этого вида проанализирован комплекс морфометрических признаков (пластических и меристических) с помощью многомерных статистических методов. По результатам исследования идентифицированы три формы Cottus amblystomopsis: 1) типичная форма, населяющая речные системы Южного Сахалина, Кунашира и Хоккайдо; 2) итурупская форма; 3) северосахалинская форма (из реки Малая Александровка).
Cottidae, Дальний Восток, многомерный анализ, морфометрия, Япония
Короткий адрес: https://sciup.org/149148538
IDR: 149148538 | УДК: 504.5 | DOI: 10.15688/nsr.jvolsu.2025.1.5
Morphological Heterogeneity of Sakhalin Sculpin Cottus amblystomopsis Schmidt, 1904 from Sakhalin, Kunashir, Iturup and Hokkaido
The article is devoted to the study of intraspecific variability of the Sakhalin sculpin, Cottus amblystomopsis (Cottidae). This species has been described from the rivers of Southern Sakhalin and is found on the islands of the southern part of the Kuril Archipelago, as well as on Hokkaido. A morphological study of Sakhalin sculpin across its entire range has not yet been undertaken. To determine morphological differences between various island populations of this species, a complex of morphometric characters (plastic and meristic) was analyzed using multidimensional statistical methods. Based on the results of the study, three forms of Cottus amblystomopsis were identified: 1) the typical form inhabiting the river systems of South Sakhalin, Kunashir and Hokkaido; 2) the Iturup form; and 3) the North Sakhalin form (from the Malaya Aleksandrovka River).
Текст научной статьи Морфологическая гетерогенность сахалинского подкаменщика Cottus amblystomopsis Schmidt, 1904 островов Сахалин, Кунашир, Итуруп и Хоккайдо
DOI:
Вид Cottus amblystomopsis (Actinopterygii: Cottidae) описан П.Ю. Шмидтом (1904) из реки Лютога, южная часть Сахалина [5]. Сахалинский подкаменщик населяет реки и их эстуарии с пониженной соленостью на островах южной части Курильской гряды, а также на Сахалине и Хоккайдо [1; 2; 4; 7]. Северной границей распространения этого вида рыб является река Тумь, Северный Сахалин, где С.В. Шедько (2003) были обнаружены взрослые и ювенильные особи сахалинского подкаменщика [4].
Таким образом, ареал C. amblystomopsis характерезуется мозаичностью. Популяции, населяющие разные острова, географически хорошо изолированы в виду того, что соленая океаническая вода является непреодолимой преградой для распространения вида. При этом, биотопические и пищевые предпочтения сахалинского подкаменщика в разных частях ареала очень сходны: взрослые особи этого вида обитают в нижнем и среднем течении горных рек на каменисто-галечных и крупнозернистых песчаных грунтах, питаются насекомыми, бентосными организмами и икрой [1; 3].
К настоящему времени изучена генетическая гетерогенность сахалинского подкаменщика на остове Хоккайдо [6]. Также изучены морфологичекие и морфометрические характеристики C. amblystomopsis с острова Сахалин и Кунашир [3; 7]. Исследований, посвященных морфологической изменчивости сахалинского подкаменщика на всем ареале до сих пор не проводилось. В настоящей работе мы проанализировали материалы по C. amblystomopsis из фондовых коллекций Зоологического института РАН (ЗИН РАН), собранные на островах Сахалин, Кунашир, Итуруп и Хоккайдо.
Целью настоящего исследования является выяснение степени морфологической гетерогенности вида C. amblystomopsis из географически изолированных островных попу- ляций. Для этого будет применен набор многомерных статистических методов для комплекса морфометрических признаков (пластических и меристических).
Материал и методы
Изученные материалы по сахалинскому подкаменщику . Всего было изучено 87 экземпляров C. amblystomopsis (включая голотип), хранящихся в ихтиологической фондовой коллекции Зоологического института РАН, Санкт-Петербург (ЗИН РАН).
Сахалин : р. Лютога, 1 экз. (голотип ZIN 12763); р. Малая Александровка, 21 экз.; р. Кирпичная, 18 экз.; р. Бахура, 12 экз.
Хоккайдо : р. Hekiriji, 12 экз.
Кунашир : оз. Песчаное, 2 экз.; с. Кинокай, 1 экз.; руч. Петрова, 4 экз.
Итуруп : р. Курилка, 16 экз.
Географическое положение изученных выборок с островов Сахалин, Хоккайдо, Итуруп, Кунашир представлено на карте (см. рис. 1). Для измерения использовались взрослые неповрежденные особи (52 экз.). Рентгеновские снимки выполнялись для всех экземпляров.
Изучение осевого скелета . Для описания осевого скелета и скелета плавников сахалинского подкаменщика использовались снимки, полученные с помощью многоцелевой рентгенодиагностической установки (ПРДУ), находящейся в составе ЦКП «Таксон» ЗИН РАН. На рентгеновских снимках определялись числа позвонков в туловищном и хвостовом отделах позвоночника, количество плевральных ребер. Также определялось число лучей в плавниках: первом и втором спинном (DI, DII), анальном (A), хвостовом (C). Для всех меристических признаков осевого скелета определялись частоты их встречаемости в каждой выборке, а также модальные значения.
Морфометрический анализ . Измерение пластических признаков C. amblystomopsis выполнялось при помощи штангенциркуля согласно схеме, разработанной специально для
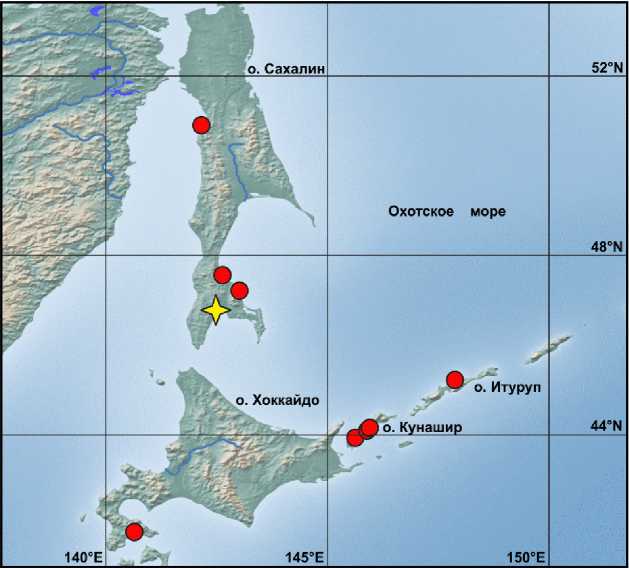
Рис. 1. Карта с точками местонахождений сахалинского подкаменщика.
Звездой обозначено место поимки типового экземпляра С. amblystomopsys (ZIN 12763)
коттоидных рыб [8]. Для каждой измеренной особи получено 24 измерения: тотальная длина ( TL ); стандартная длина ( SL ); длина головы ( HL ); антедорсальное расстояние; антеа-нальное расстояние; максимальная высота тела; высота хвостового стебля; длина хвостового стебля; длина основания первого спинного плавника; длина основания второго спинного плавника; длина основания анального плавника; высота первого спинного плавника; высота второго спинного плавника; высота анального плавника; длина грудных плавников; длина брюшных плавников; горизонтальный диаметр глаза; высота головы у затылка; длина рыла; заглазничное расстояние; длина верхней челюсти; длина жаберной щели; межглазничное расстояние; ширина головы.
Статистический анализ. Анализ изменчивости по комплексу пластических признаков проводился по их абсолютным и относительным (% SL) величинам. Для каждого из относительных измерений рассчитывалось среднее значение и стандартное отклонение. Оценка межвыборочной изменчивости по признакам проводилось методом главных компонент (PCA). PCA выполнялся на основе корреляционных матриц в программе PAST version 4.03. Выделенные главные компоненты в дальнейшем рассматривали как новые некоррелированные признаки. Для изучения общей вариабельности формы тела были использованы компоненты, вносящие наибольший вклад в общую дисперсию. Меру и направление изменчивости исходных признаков определяли по знаку и величине факторных нагрузок (коэффициентов корреляции). Большинство исходных пластических признаков имели высокие и положительные факторные нагрузки на первую главную компоненту (PC1). Это значит, что PC1 определяет в первую очередь размерные различия между особями, а не разницу в форме тела. Для описания различий, не связанных с размерной изменчивостью, была построена диаграмма рассеяния в пространстве второй и третей компонент. Каждая выборка формирует на графике свой кластер (морфопространство). Степень перекрывания кластеров свидетельствует о степени их морфологической дифференциации.
Для оценки надежности различий между выборками C. amblystomopsis из разных островов по комплексу пластических признаков был проведен дискриминантный анализ (DFA), реализованный в STATISTICA 10
(StatSoft). Дискриминантный анализ выполнен на основе относительных значений (% SL), характеризующих форму тела рыбы. Оценка качества дискриминации проведена при помощи показателей Wilks’ Lambda и F-критерия. Близкие к 0 значения Wilks’ Lambda свидетельствуют о хорошо выраженной дискриминации. Масштаб различий между популяциями сахалинского подкаменщика определялся на основе значений квадрата расстояния Ма-халанобиса. Вклад каждого признака в диск-римининирующую силу модели оценивался по значению Partial Lambda. Чем меньше этот показатель, тем выше вклад переменной. Результаты этого анализа визуализированы с помощью диаграммы рассеяния канонических переменных в пространстве первой и второй дискриминантных осей.
Результаты и обсуждение
Анализ изменчивости пластических признаков сахалинского подкаменщика из разных островных популяций . Использование метода главных компонент (рис. 2 a ) позволило наглядно показать вариабельность всего комплекса пластических признаков внутри и между изученными выборками C. amblystomopsis из разных островных популяций. Оценка вклада признаков в дисперсию по каждой из осей (PC2 и PC3) выполнялась на основе значений факторных нагрузок.
Дифференция выборок по второй главной компоненте определяется, главным образом четырьмя признаками, имеющими наиболь- шие факторные нагрузки на эту ось: длина V (–0,421), высота DI (0,275), высота DII (0,238), длина верхней челюсти (–0,220). Таким образом, различия по второй главной компоненте определяются, в основном. размерами плавников. Наибольшие нагрузки на третью главную компоненту имели три признака: длина межжаберного промежутка (0.409), межглазничное расстояние (0,252), длина P (–0,233). Дифференциация выборок по этой оси, прежде всего, связана с признаками формы головы.
Значимость попарных различий между выборками была определена с помощью многомерного дисперсионного анализа (MANOVA), в качестве данных для которого использовались значения, полученные для 2 и 3 главных компонент. Выявлены статистически значимые различи по признакам формы тела между выборками сахалинского подкаменщика из южного и северного Сахалина ( p = 0,001). Подтверждаются различия C. amblystomopsis островов Курильской гряды (Кунашир, Итуруп) от особей с Хоккайдо ( p = 0,001–0,005) и южного Сахалина ( p = 0,006–0,007).
Анализ изменчивости меристических признаков сахалинского подкаменщика из разных островных популяций . С помощью метода главных компонент также была изучена вариабельность комплекса меристических признаков внутри и между изученными выборками C. amblystomopsis . Поскольку значения меристических признаков не зависят от размера особей, мы использовали первую и вторую компоненту для визуализации отличий на диаграмме (рис. 2 b ).
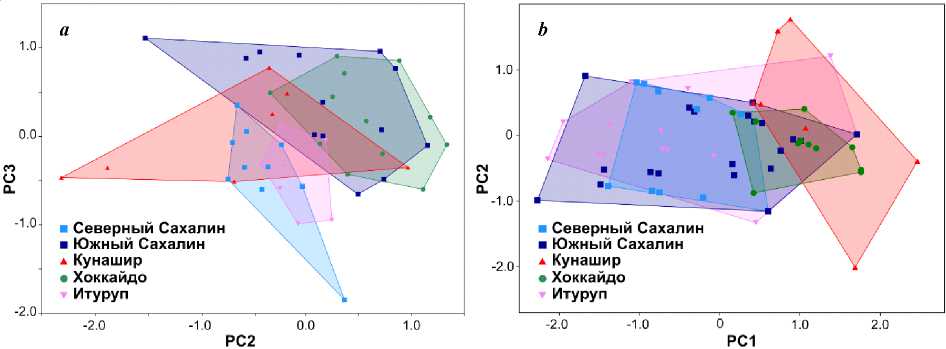
Рис. 2. Результаты анализов, выполненных методом главных компонент: а – комплекс пластических признаков; b – комплекс меристических признаков
Экземпляры сахалинского подкаменщика из рек и озер островов Сахалин (северная и южная части), Кунашир, Итуруп и Хоккайдо образуют единый, но неоднородный кластер на графике рассеивания (рис. 2 b ). Внутри этого кластера наблюдается заметная дифференциация особей из северного Сахалина и Кунашира (их морфопространства на графике не пересекаются). Также наблюдается заметное расхождение выборок из северного Сахалина и Хоккайдо. В отличие от результатов PCA, выполненных на основе пластических признаков, выборки из южного и северного Сахалина почти полностью перекрываются на диаграмме (рис. 2 b ), что говорит о том, что счетные признаки не могут дифференцировать эти популяции, хорошо отличающиеся по форме тела.
Различия между выборками по первой главной компоненте определяются двумя признаками, имеющими наибольшие факторные нагрузки на эту ось: число лучей в DII (0,836), число лучей в A (0,823). Следовательно, различия по первой главной компоненте определяются, в основном, числами лучей в непарных плавниках. Наибольшие нагрузки на вторую главную компоненту имели: общее число позвонков (0,792) и число хвостовых позвонков (0,773). Можно заключить, что дифференциация выборок по этой оси связана с количеством позвонков в хвостовом отделе позвоночника. Этот отдел, как правило, гораздо более вариабелен у коттоидных рыб по сравнению с туловищным.
Статистическая значимость различий между выборками по комплексу меристических признаков оценивалась также с помощью MANOVA. В отличие от анализа, проведенного по пластическим признакам, значимая дифференциация между южно- и севоросахалинс-кой популяциями не обнаружена ( p = 0,605). По комплексу меристических признаков выявлены статистически значимые ( p = 0,0001– 0,0023) отличия C. amblystomopsis из Кунашира и Хоккайдо от всех остальных выборок. При этом различия между ними самими не значимы ( p = 0,379).
Классификация популяций сахалинского подкаменщика на основе комплекса пластических признаков . Результаты PCA, полученные при анализе пластических и мерис- 50
тических признаков, показали морфологическую неоднородность C. amblystomopsis в изученной части ареала. Учитывая относительно низкий масштаб различий по меристическим признакам классификация выборок выполнена только по комплексу измерений, отражающих изменчивость формы тела рыбы. Для оценки качества дифференциации между выборками использован дискриминантный анализ (рис. 3), в котором в качестве данных использованы относительные значения измерений (% SL). Дополнительно добавлены измерения для голотипа C. amblystomopsys , собранного в р. Лютога (Южный Сахалин). По причине очень крупного размера и меньшей степени сохранности, чем другие использованные экземпляры этого вида, типовой экземпляр не был использован для анализа главных компонент.
Проведенный анализ показал высокое качество дискриминации выборок (Wilks’ Lambda = 0,0007, approx. F(104,81) = 4,123, p < 0,0001). Наименьшие квадраты расстояния Махаланобиса (23.96–44.83) выявлены между выборками из Южного Сахалина, Хоккайдо и Кунашира. Наибольшее расхождения наблюдалось между выборками из Хоккайдо и Северного Сахалина (квадрат расстояния Махаланобиса = 83,10), а также между выборками Северного Сахалина и Кунашира (квадрат расстояния Махаланобиса = 80,44).
Признаки, вносящие наибольший вклад в дискриминирующую модель определены на основе показателя Partial Lambda. Наибольшее значение (в порядке уменьшения) имели следующие семь признаков: длина рыла (0,492), высота головы (0,511), максимальная высота тела (0,517), ширина головы (0,542), длина жаберной щели (0,553), длина основания первого спинного плавника (0,569), длина верхней челюсти (0,583). Большая часть (пять из семи) признаков, вносящих наибольший вклад в дискриминирующую модель, связаны с характеристиками головы. Признак «длина верхней челюсти» также находился среди показателей, имевших наибольшую нагрузку на ось PC 2 в анализе главных компонент. Признаки головы, как правило, имеют высокую точность измерения и могут быть полезны для идентификации форм.
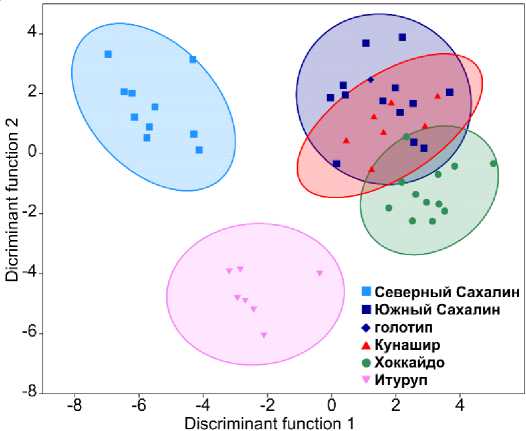
Рис. 3. Результаты дискриминантного анализа, проведенного на основе комплекса относительных пластических признаков (% SL)
Результаты дискриминантного анализа показали высокую степень отличий выборок из Северного Сахалина и Итурупа от остальных трех выборок, которые попадают в один кластер, который можно обозначить как особей, соответствующих типичной форме (рис. 3). Типовой экземпляр вида (точка в виде ромба) попадает почти в середину кластера «Южный Сахалин + Кунашир + Хоккайдо». Особи из водоемов острова Итуруп образуют хорошо обособленный кластер в пространстве первой и второй дискриминантных осей. Так же хорошо дифференцированы от всех остальных особи из Северного Сахалина. Обе эти выборки представляют собой формы, хорошо отличающиеся от типичного C. amblystomopsis , которые можно определить по набору морфометрических признаков.
Заключение
По результатам анализа морфометрических признаков можно дифференцировать три формы сахалинского подкаменщика:
-
1) типичная форма С. amblystomopsis ; имеет наибольшую ширину головы (31,91 ± 2,13 % SL против 28,34 ± 2.,27 % SL у северосахалинской и итурупской форм), более широкое межглазничное расстояние (5,78 ± 0,80 % SL против 4,38 ± 0,44 % SL) и наибольшую высоту первого спинного плавника (8,62 ± 0,99 % SL против 7,29 ± 0,75 % SL).
-
2) итурупская форма С. amblystomopsis ; отличается от остальных форм наименьшей высотой анального плавника (12,38 ± 1,54 % SL против 14,55 ± 1,34 % SL у типичной формы и 13,45 ± 0,79 % SL у северосахалинской).
-
3) северосахалинская форма C. ambly-stomopsis ; характеризуется наименьшей длиной основания первого спинного плавника (16,64 ± 2,53 % SL против 18,70 ± 1,79 % SL у типичной формы и 18,89 ± 1,23 % SL у иту-рупской).
Для дальнейшего установления таконо-мического статуса и взимосвязей описанных форм необходимо проведение молекулярногенетического исследования особей из разных островных популяций этого вида рыб.
