Морфологическая изменчивость корневищ душицы обыкновенной (Orig'anum vulg'are L.) и разнообразие способов освоения пространства как компонент устойчивого состояния вида в сообществе
 и разнообразие способов освоения пространства как компонент устойчивого состояния вида в сообществе")
Автор: Дубровная Светлана Алексеевна, Савельева Екатерина Николаевна, Мавлюдова Ляйля Усмановна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 5-1 т.17, 2015 года.
Бесплатный доступ
В условиях лесных полян у душицы обыкновенной отмечали формирование корневищ с различной длиной междоузлий. Это способствовало формированию компактных участков клона (центров разрастания) и диффузных участков клона (коммуникационных корневищ). Максимальное образование побегов отмечается в центрах разрастания, которые соединяются друг с другом посредством коммуникационных корневищ. Онтогенетическое развитие надземных побегов, возникших на различных функциональных участках корневищ, не различалось. Парциальные побеги, определяющие рост главных и боковых осей, имели средневозрастное онтогенетическое состояние. Побеги, образованные на старых участках корневищ, соответствовали особям прегенеративного периода. Изъятие генеративных побегов в компактных участках клона не ведет к существенному ухудшению состояния ценопопуляции. Сбор генеративных побегов на коммуникационных участках корневищ ведет к снижению побегообразования, что в итоге затрудняет ветвление, поэтому сбор надземных побегов необходимо производить в местах компактного расположения цветущих побегов, с интервалом раз в два года, оставляя часть побегов для образования семян.
Лекарственные растения, ресурсосберегающие технологии
Короткий адрес: https://sciup.org/148203997
IDR: 148203997 | УДК: 581.4;
Morphological variability of oregano (Orig'anum vulg'are L.) rhizomes and variety of ways of space development as a component of steady state type of community
In the conditions of forest glades at marjoram ordinary it was noted formation of rhizomes with various length of interstices. It promoted formation of compact sites of clone (the centers of growth) and diffusion sites of clone (communication rhizomes). The maximum formation of sprouts is noted in the centers of growth which connect with each other by means of communication rhizomes. Ontogenetic development of the elevated sprouts which arose on various functional sites of rhizomes didn't differ. The partial sprouts determining growth of the main and side axes had a middle-aged ontogenetic state. The escapes formed on old sites of rhizomes corresponded to individuals of the pregenerative period. Withdrawal of generative sprouts in compact sites of a clone doesn't conduct to essential deterioration of coenopopulation state. Collecting generative sprouts on communication sites of rhizomes conducts to decrease in a sprout formation that as a result complicates branching therefore collecting elevated sprouts needs to be made in places of compact arrangement of the blossoming sprouts, with an interval of times in two years, leaving part of sprouts for formation of seeds.
Текст научной статьи Морфологическая изменчивость корневищ душицы обыкновенной (Orig'anum vulg'are L.) и разнообразие способов освоения пространства как компонент устойчивого состояния вида в сообществе
Лекарственные растения естественных сообществ остаются важными источниками сырья для фармакологического производства, а также широко востребованы населением, что определяет значительное давление на природные популяции. В то же время по ресурсному использованию лекарственных видов существуют рекомендации, которые основаны на определении допустимых объемов ежегодных заготовок [1-3]. Для этого используют ресурсный запас объекта на изучаемой территории, принимая во внимание средние сроки восстановления, а именно: при сборе соцветий у многолетних растений и надземных органов однолетних растений необходимый временной интервал для восстановления ценопопуляции 2 года, надземных органов многолетних растений – 4-6 лет, подземных органов – 15-20 лет [4, 5].
Для разработки адекватных рекомендаций [6] по сбору лекарственного сырья душицы обыкновенной, важным направлением исследования является изучение гетерогенности подземных органов полицентрической биоморфы, определяющих рост, развитие и положение в пространстве надземных побегов различных онтогенетических
состояний. Побеги сходного онтогенетического состояния в пределах полицентрической биоморфы могут иметь различную функциональную нагрузку, а потому изъятие их в пределах особи может неоднозначно сказаться на устойчивости це-нопопуляции (ЦП).
Цель исследования: изучить особенность формирования морфологической структуры подземных вегетативных органов душицы обыкновенной (Origánum vulgáre L.), для разработки рекомендаций по сбору лекарственного сырья в условиях естественных сообществ.
Материал и методика. Объектом исследования являлись ценопопуляции растения длиннокорневищной жизненной формы душицы обыкновенной ( Origánum vulgáre L.). Растение луговоопушечной эколого-ценотической группы, которое широко представлено во флоре европейской части России. В Республике Татарстан растет на полянах, опушках, среди кустарников, на сухих открытых травянистых местах, по склонам холмов, на суходольных и пойменных лугах, образуя скопления [7]. При этом ЦП на лесных полянах и опушках испытывают значительный антропогенный прессинг от неконтролируемого сбора надземной биомассы в качестве лекарственного сырья.
Исследования проводились в северо-восточной части Приволжской возвышенности, на правобережье Волги, в Верхнеуслонском районе РТ. Район характеризуется умеренно-континентальным климатом с теплым летом и умеренно- холодной зимой и переходом почвенно-растительных зон от леса к степи. Леса района представлены широколиственными видами. Для изучения структуры особи и ЦП душицы обыкновенной в Верхнеуслонском районе были выбраны несколько лесных опушек в дубраве лещино-пролесниковой. Травянистый ярус представлен растениями неморальной и лугово-опушечной эколого-ценотичес-кой групп. Отмечалось внедрение видов остепнен-ных лугов. Почвы серые лесные, травяно-кустарничковый ярус цельный, хорошо выражен процесс задернованности. Опушка леса характеризовалась значительным варьированием освещенности. Растения, произрастающие в центральной части опушки, существовали в условиях фоновой освещенности. Освещенность на участках, примыкающих к лесу, была существенно ниже (от 10 до 60% от фоновой), что связано с разрастанием подлеска.
Счетная единица у длиннокорневищных растений может быть представлена частью растения – побегом или парциальным кустом [8, 9]. При исследовании растений длиннокорневищной жизненной формы мы учитывали рекомендации по выбору счетной единицы Е.Л. Любарского [10]. В качестве счетной единицы были использованы парциальные побеги или парциальные кусты, с хорошо выраженной корневой системой. При морфологической целостности подземных частей, отельные парциальные побеги оказываются физиологически автономными и разобщенными [10, 11]. Побеги, развивающиеся на корневище, но не формирующие собственную корневую систему, в качестве счетной единицы не использовались. Всего было проанализировано более 20 особей вида. У растений учитывали количество рамет, длину междоузлия корневищ, интенсивность ветвления на разных участках корневища, онтогенетическое состояние рамет, их положение на корневище [12]. Для изучения онтогенетической структуры были заложены трансекты 20х1 м. Определялось онтогенетическое состояние каждого парциального побега. Растения выкапывались, положение побегов на корневище и относительно друг друга переносили на карту-схему. Статистическую обработку данных проводили путем использования программы Statistica 5.1. При отсутствии нормального распределения выборки оценивали медиану (Ме), для проверки гипотезы о равенстве генеральных средних двух независимых выборок был использован непараметрический критерий Уил-консона-Манна-Уитни, с оценкой р-уровня значимости (p-level).
Результаты исследования. Анализ строения подземных корневищ душицы обыкновенной показал, что в пределах сформированной полицентрической биоморфы длина междоузлий корневищ неравнозначна. Хорошо выделялись участки с короткими междоузлиями, на которых отмечалось интенсивное образование надземных побегов. Такие участки корневища были нами определены как центры разрастания (компактные участки клона). Центры разрастания, образованные скоплениями большого количества надземных побегов, соединялись друг другом посредством корневищ с длинными междоузлиями (коммуникационные корневища или диффузные участки клона), на которых отмечалось минимальное количество побегов (табл. 1).
Таблица 1. Изменчивость длины междоузлий в пределах полицентрической биоморфы
|
Тип участка корневища |
Сре днее |
Стандартная ошибка среднего |
Медиа на |
Диапазон изменчивости признака |
Нижняя и верхняя границы 95% доверительного интервала медианы |
|
коммуникационные корневища |
5,5 |
0,3 |
5 |
1 - 14 |
3 - 7 |
|
центры разраста ния |
2,8 |
0,2 |
2 |
1 - 12 |
2 - 3 |
Примечание: разница статистически значима р=0,0000001
Таблица 2. Интенсивность побегообразования на разных участках корневища полицентрической биоморфы
|
Тип участка корневища |
Число междоузлий |
Число рамет |
Интенсивность побегообразования |
|
коммуникационные корневища |
153 |
55 |
0,36 |
|
центры разрастания |
48 |
74 |
1,5 |
Примечание: разница статистически значима р =0,000021
Интенсивность побегообразования оценивалась нами дифференцировано для разных участков корневищ, в центрах разрастания и на коммуникационных корневищах. Проведенные исследования показали, что на корневищах с длинными междоузлиями, которые были определены как коммуникационные участки, на каждое междоузлие приходилось 0,3 побега (интенсивность побегообразования). Значительно больше побегов в расчет на одно междоузлие образовывалось на участках, которые мы определили как центры разрастания (табл. 2).
Изучение вегетативного разрастания показало, что число образовавшихся побегов на корневищах, характеризующихся укороченными междоузлиями, существенно выше. Среднее число побегов здесь составило 5.4, несколько меньше был показатель медианы – 4.5. Число образовавшихся побегов на коммуникационных корневищах составило – 1.5 (табл. 3). Формирующийся компактный участок клона обеспечивает длительное удержание территории и максимальное вегетативное разрастание. На таких участках разрастание клона идет как за счет роста новых корневищ в разные направления, на которых образуются надземные побеги, так и на старых участках. Корневища с длинными междоузлиями, разрастаясь диффузно, обеспечивают освоение новых территорий. Интересно отметить, что в условиях луговых сообществ формируются, как правило, компактные клоны (скопления) в то время как в зарослях кустарника – диффузные. Гетерогенные условия лесных опушек определяет разнообразные способы освоения пространства.
Таблица 3. Характеристика вегетативного разрастания на разных участках корневища (число образовавшихся надземных побегов)
|
Типы побегов корневища |
Сре днее |
Стандартная ошибка среднего |
Медиана |
Диапазон изменчивости признака |
Нижняя и верхняя границы 95% доверительного интервала медианы |
|
коммуникационный участок |
1,5 |
0,2 |
1,0 |
0 - 4 |
1 - 2,5 |
|
центр разрастания |
5,4 |
0,8 |
4,5 |
1- 10 |
3 - 9 |
Примечание: разница статистически значима р=0,0000001
Несмотря на то, что участки корневища в пределах полицентрической биоморфы существенно различались по качеству, развитие надземных парциальных побегов в пределах клона не отличалось. Частота растения различных онтогенетических групп на коммуникационных участках и центрах разрастания была сходной (рис. 1). На всех участках клона была высокой доля потенциальных побегов вегетативного размножения (ППВР), которые на момент наблюдения не реализовывались в полноценные ассимиляционные побеги полицентрической биоморфы.

Рис. 1. Спектр онтогенетических состояний рамет, возникших на разных участках корневища душицы обыкновенной (коммуникационные корневища, в центрах разрастания)
Онтогенетический спектр ценопопуляции нами определялся двумя способами: 1 – на основе подсчета надземных побегов выкопанных полицентрической биоморфы, 2 – с использованием традиционной методики подсчета растений и определения их онтогенетического стояния на временных трансектах. Сравнение онтогенетических спектров проводилось с использованием критерия χ2. Под «наблюдаемым» вариантом подразумевалось соотношение парциальных побегов на основе выкопанных полицентрических биоморф.
Преимущество определения онтогенетической структуры данным способом заключается в том, что здесь точно учитываются парциальные образования с хорошо выраженной корневой системой и не учитываются побеги, развивающиеся из спящих почек корневища, но не формирующие корневую систему, т.е. побеги, которые нельзя учитывать как счетные единицы. Под «предполагаемым» подразумевалось соотношение растений различных онтогенетических групп, подсчитанных непосредственно в природе на трансекте. Проведенные исследования показали, что соотношение растений онтогенетических групп, выявленных разными способами, не различалось (χ2=8,69; ν = 5 P<0,05).
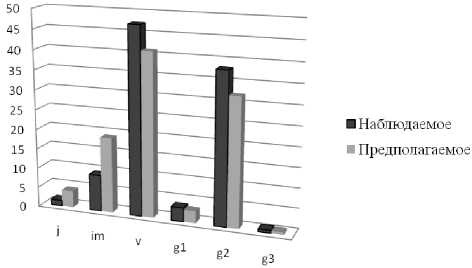
Рис. 2. Онтогенетические спектры ЦП душицы обыкновенной, построенные на основе непосредственного пересчета парциальных побегов особи и на трансекте
Можно видеть некоторое уменьшение доли растений имматурного онтогенетического состояния в онтогенетическом спектре, построенном на основе непосредственной идентификации счетных единиц. Однако разница между «наблюдаемой» и «предполагаемой» онтогенетической структурой была незначимой. Не различались также демографические показатели ценопопляции, подсчитанные различными способами. Уменьшение плотности ценопопуляции связано с тем, что побеги, возникающие на старых участках корневища, но не формирующие собственную корневую систему, мы не рассматривали в качестве счетных единиц, в то время как при определении плотности и онтогенетической структуры на трансекте данные побеги учитывались. Таким образом, анализ онтогенетической структуры ЦП на временных трансектах является достаточно корректным способом, хотя при этом учитываются побеги, возникшие на корневищах из спящих почек. Такие побеги также могут перейти к ветвлению, однако на момент наблюдения, не формируя собственную корневую систему, побеги не рассматривались в качестве самостоятельных счетных единиц.
Таблица 2. Демографические показатели ЦП O. vulgare
|
Варианты наблюдения |
∆ |
ω |
Тип популяции |
I 1 |
Плотность |
|
наблюдаемой структуре |
0,2656 |
0,6292 |
молодая |
0,5775 |
9,47 |
|
предполагаемой структуре |
0,2318 |
0,5594 |
молодая |
0,6429 |
11,2 |
В центрах разрастания побеги возобновления и побеги, возникшие из спящих почек, различных онтогенетических состояний расположены достаточно близко на корневище. На каждое междоузлие приходится 1,5 побегов. Это позволяет предположить о взаимоучастии всех побегов в формировании корневища – заложении и формировании почек возобновления, накоплении питательных веществ. В этом случае изъятие генеративных побегов существенно не повлияет на функционирование компактного участка клона.
Для коммуникационных корневищ рост главного и боковых осей определяется побегом возобновления. Такие парциальные побеги имели средневозрастное генеративное онтогенетическое состояние. Побеги, возникающие из спящих почек, соответствовали растениям прегенеративного периода. При удалении парциального побега средневозрастного онтогенетического состояния главной и боковой оси может существенно снизиться процесс побегообразования и ветвление корневищ. Кроме того, парциальный побег определяет направление роста главной и боковой оси, накопление и мобилизацию питательных веществ. А потому сбор сырья в условиях разреженного произрастания душицы нецелесообразен, поскольку это существенно ухудшит устойчивость вида, вплоть до его выпадения из состава сообщества.
Выводы: в процессе освоения пространства в пределах полицентрической биоморфы душицы обыкновенной могут формироваться корневища с различной длиной междоузлий, интенсивностью побегообразования, что определяет формирование центров разрастания (компактные участки клонов), соединенных между собой коммуникационными корневищами. В пределах центров разрастания отмечается максимальное побегообразование. Побегообразование идет во все направления, что способствует разрастанию компактного участка в пространстве. Коммуникационные корневища обеспечивают функцию и вегетативного размножения и удаления побегов возобновления на отдаленные территории. Развитие побегов различных участков клона не различалось, однако число парциальных побегов на м2 в условиях компактных участков было значительно выше. Хорошо развитые парциальные побеги средневозрастного генеративного состояния обеспечивают не только функцию пополнения ценопопуляции семенами, но и формируют максимально развитые почки возобновления и спящие почки.
Сбор сырья должен производиться с нанесением минимального урона ЦП, чтобы временной интервал, необходимый для ее восстановления также был минимальным. Сбор сырья в условиях лесных полян целесообразно производить на участках максимального скопления парциальных побегов средневозрастного онтогенетического состояния, что наблюдается в компактных участках клона.
Список литературы Морфологическая изменчивость корневищ душицы обыкновенной (Orig'anum vulg'are L.) и разнообразие способов освоения пространства как компонент устойчивого состояния вида в сообществе
- Буданцев, А.Л. Оценка современного состояния ресурсов важнейших лекарственных и пищевых растений флоры России//Фундаментальные основы управления биологическими ресурсами. -М., 2005. С. 87-92.
- Крылова, И.Л. Методика определения запасов лекарственных растений. -М., 1986. 52 с.
- Методика выявления дикорастущих сырьевых ресурсов при лесоустройстве. -М: Госкомплекс СССР, 1987. 54 с.
- Правила заготовки пищевых лесных ресурсов и сбора лекарственных растений.//Приказ МПР РФ от 10 апреля 2007 г. №83. 5 с. URL: http://www.forestforum.ru/info/laws/normativy/Pishev_lekarstv.doc
- Федоров, Н.И. О методических подходах к контролю динамики важнейших ресурсных видов лекарственной флоры Республики Башкортостан/Н.И. Федоров, С.Н. Жигунова, О.И. Михайленко//Известия Самарского научного центра РАН. 2011. Т. 13, № 5(3) С. 113-115
- Подгаевская, Е.Н. Онтогенез душицы обыкновенной (Origanum vulgare L.)//Онтогенетический атлас лекарственных растений. -Йошкар-Ола, 2002. Т.3. С. 214-217.
- Бакин, О.В. Сосудистые растения Татарстана/О.В. Бакин, Т.В. Рогова, А.П. Ситников. -Казань: КГУ, 2000. 496 с.
- Ценопопуляции растений: (Очерки популяционной биологии). -М., 1988. 182 с.
- Широков, А.И. Изучение ценопопуляций растений/А.И. Широков, В.П. Воротников, А.К. Ибрагимов. -М.: Наука, 1994. 38 с.
- Любарский Е.Л. Экология вегетативного размножения высших растений. Казань, 1967. 180 с.
- Басаргин, Е.А. Биоморфология некоторых длиннокорневищных видов растений и структура их ценопопуляций на юге Сибири. Автореф. дисс. на соиск. уч. степ. кандидата биологических наук. -Новосибирск, 2010. 136 с.
- Дубровная, С.А. Методы изучения растительных сообществ: учеб. -метод. пособ/С.А. Дубровная, О.И. Волков -Казань: Бриг, 2009. 118 с.
