Морфологические типы организации пиреноидов у зеленых монад

Автор: Болдина Ольга Николаевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Материалы лекций и устных докладов
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
В статье обсуждается роль пиреноида у зелёных монадных водорослей. У видов Chlamydomonas к настоящему времени выявлено 6 типов пиреноидов, различающихся между собой строением крахмальной обкладки, числом и характером расположения тилакоидов в строме пиреноида. Дан анализ распределения типов пиреноидов в группах Chlamydomonas и на кладограммах, предлагаемых разными авторами. Высказывается предложение рассматривать пиреноид как пример модульной организации. Ставится вопрос о гомологии пиреноидов у зелёных водорослей.
Гомологии пиреноидов у зелёных водорослей, тилакоидов, пиреноида
Короткий адрес: https://sciup.org/146116182
IDR: 146116182 | УДК: 582.26
Текст научной статьи Морфологические типы организации пиреноидов у зеленых монад
Ботанический институт им. В.Л. Комарова РАН, Санкт-Петербург
В статье обсуждается роль пиреноида у зелёных монадных водорослей. У видов Chlamydomonas к настоящему времени выявлено 6 типов пиреноидов, различающихся между собой строением крахмальной обкладки, числом и характером расположения тилакоидов в строме пиреноида. Дан анализ распределения типов пиреноидов в группах Chlamydomonas и на кладограммах, предлагаемых разными авторами. Высказывается предложение рассматривать пиреноид как пример модульной организации. Ставится вопрос о гомологии пиреноидов у зелёных водорослей.
Пиреноид – крупная структура в составе хлоропластов водорослей, имеющая высокую степень преломления световых лучей в оптическом микроскопе, как правило, окруженная обкладкой из полисахаридных метаболитов. Помимо водорослей пиреноиды обнаружены и у некоторых высших растений – у антоцеротовых мхов и цикадовых растений.
Физиологическая роль пиреноида дискутируется с 70-х гг. ХХ в. и до сегодняшнего дня не совсем ясна. Значительную его часть составляет фермент РДФ-карбоксилаза, являющийся ключевым ферментом в фотосинтетических реакциях растений [1, 21]. Однако многие виды водорослей, не имеющие пиреноида, прекрасно вегетируют и осуществляют процессы фотосинтеза с той же эффективностью, что и содержащие пиреноид [16]. Обнаруженные в пиреноиде следы нуклеиновых кислот позволяют осуществлять в некоторых случаях самостоятельное деление, совмещенное по времени с делением хлоропластов водорослей. В других случаях пиреноиды не делятся, а просто исчезают в процессе деления и заново появляются в хлоропластах у вновь образованных клеток [5].
Наблюдения за пиреноидами позволили выявить различия в их строении еще в начале прошедшего века. Были предложены их классификации [3, 9, 18]. Впоследствии помимо классификаций [5] указывались их отличительные особенности [6].
Существенный прогресс в изучении морфологии пиреноидов был связан с появлением методов просвечивающей электронной микроскопии. С того момента у разных таксонов водорослей были описаны пиреноиды, различающиеся тонким строением его компонетов [4, 8, 11, 12, 13, 14, 15 и др.].
Наибольшее разнообразие в ультраструктуре пиреноида было выявлено у зеленых монадных водорослей, относимых к роду Chlamydomonas [2].
Хламидомонады – это большая группа организмов, насчитывающая, согласно классическим представлениям [7], более 500 видов. Для удобства классификации род был разбит на подроды и секции [10, 18], а впоследствии – только на группы [5, 7]. В основу таксономии хламидомонад положены взаимосвязанные признаки «форма хлоропласта» и «расположение и число пиреноидов в хлоропласте» (рис. 1).
К настоящему моменту у Chlamydomonas выделено 6 типов пиреноидов. Типы различаются особенностями строения крахмальной обкладки (сплошная или фрагментированная – из крупных пластинок или из мелких зерен крахмала), числом тилакоидов в пачке (1, 2, 3 или более) и характером их распределения в строме пиреноида.
Распределение типов по группам хламидомонад представлено на рис. 1. Таким образом, таксономические группы Chlamydomonas неоднородны. Но во многих из них выявляются доминирующие типы.
Так, у наиболее характерной и многочисленной группы рода – группы Euchlamydomonas – превалирует IV тип пиреноида ( C. angulosa, C. augusta, C. bullosa, C. debaryana, C. gelatinosa, C. nivalis, C. parallelistriata, C. reinhardtii, C. regina, C. spaeroides ). При этом установлено наличие единого типа пиреноида у штаммов, относимых к одному виду ( C.reinhardtii – 6 штаммов и C.debaryana – 7 штаммов).
III тип пиреноида наиболее характерен для видов группы Chlamydella ( C. elliptica var. brittanica, C. moewusii, C. monadina ) и Amphichloris ( C.pseudopertusa, C. desmidii ). Только у двух видов группы Chlamydella ( C. noctigama и C. planoconvexa ) обнаружен V тип.
В группе Chlorogoniella отмечено наибольшее разнообразие типов пиреноидов. Так, большинство изученных видов ( C. asymmetrica, C. baca, C.badensis, C. minutissima, C. proteus, C. oblonga ) содержит пиреноид IV типа; 2 вида ( C. applanata, C. gloephila var irregularis ) – I тип, а C. сhlorococcoides ( 2 штамма) – VI тип.
В группах с малым числом видов: Agloe ( C. mutabilis ), Pseudagloe ( C. agloeformis ) и Pleiochloris ( C. carrizoensis ) – выявлен пиреноид II типа.
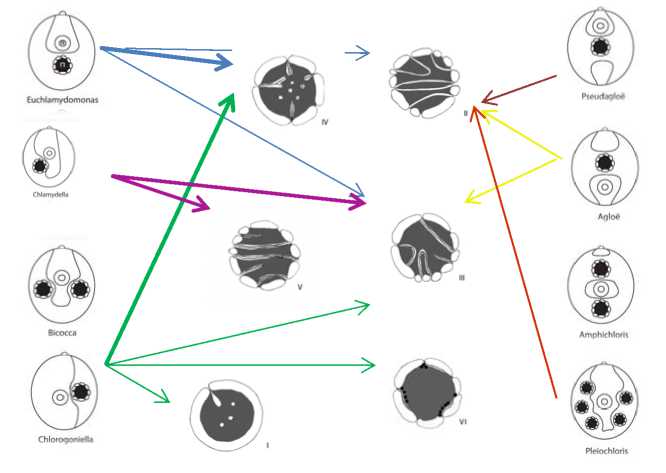
Рис. 1. Распределение типов пиреноидов ( I – VI ) в группах Chlamydomonas . Жирными стрелками указаны наиболее характерные для группы типы пиреноидов
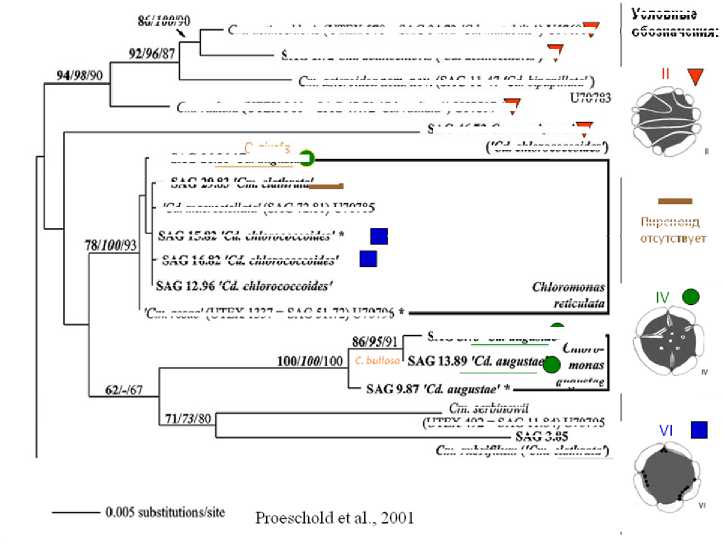
С развитием методом молекулярной биологии и появлением многочисленных деревьев, содержащих различные виды зеленых водорослей, нам представилась возможность оценить расположение типов пиреноидов на этих деревьях [17; 19 ; 20].
Сопоставление таких данных для одних и тех же штаммов зеленых монад выявило корреляцию типов пиреноидов с кладами, полученными разными исследователями. При этом в одной и той же кладе нередко объединялись водоросли с различным таксономическим положением, но имеющие один и тот же тип пиреноида.
86 Ш'УП
Cm. asteroidea nom. nov. (SAG 11-47 Cd. bipapillata*)
U7O783
- 'Cd. macrostellata (SAG 72.81) U70785
SAG 15,82 'Cd, chlorococcoides' *
- SAG 16.82 Cd. chlorocwcoides*
SA<; 12.96 'Cd. chiorococcoides'
Cm. rosae (UTEX 1337 = SAG 51.72) U70796
SAG 5.73 *Cd. august^---- ChltirO-
Условные обозначения:
i шреноид
UKVKiey^T
L hhiromimax reticulata
SAG 46.72 Cm. carrizoensi^^
(Cd. chlorococcmdesi
Cm. acrinochloris (UTEX 578 = SAG 34.72 'Cd. mutabdis*) U57693V
— SAG 26.86 *C(t augustd® i— SAG 29.83 ’Cm. clatitrato^^— augusrae
Cm. s^rhitiowii
(UTEX 492 = SAG 11.84) U70795
SAG 3.85
Cm. rubrifilum ( Cm. clathrcim^
Рис. 2. Пример сопоставления типов пиреноидов и данных молекулярнофилогенетического анализа у представителей рода Chlamydomonas (Cm )
SAG 1.72 Cm. acrinochloris CCtL acrinochloris*) ™
Cm. radiate (UTEX 966 = SAG 47.72 ’Cd. radiata' ) U57597
По мере накопления собственного и литературного материала гипотеза о прогностическом характере анализируемых деревьев (касательно отдельных их ветвей) все больше подтверждается (рис. 2).
В то же время разные клады могут содержать водоросли с одинаковым типом пиреноида. Следовательно, тип пиреноида у зеленых водорослей может быть рассмотрен и как пример их модульной организации.
Дальнейший анализ распределения типов пиреноидов поможет установить или отвергнуть их гомологию у разных групп Chlorophyta.
