Морфологическое и анатомическое строение генеративных органов разных половых форм будры плющевидной {Glechoma hederacea L.)
")
Автор: Анисимова А.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 6, 2005 года.
Бесплатный доступ
Излагаются результаты сравнительного морфолого-анатомического изучения обоеполых и женских цветков Glechoma hederacea. Найдены отличия в размерах всех органов цветка, за исключением завязи. Сравнительно-анатомическое исследование строения венчика и чашечки позволило также выявить некоторые отличительные признаки: женские цветки имеют меньшие размеры клеток, характеризуются большим опушением простыми и железистыми волосками, большим количеством устьиц и эфирномасличных железок.
Короткий адрес: https://sciup.org/147204375
IDR: 147204375 | УДК: 581.8:
The morphological and anatomical structure of generative organs of different sexual forms of Glechoma hederacea L
The results of comparative morphology-anatomical study bisexual and female flowers of Glechoma hederacea are expounded. The differences in dimensions of all the organs of the flower are found, except for the overy. The comparatively-anatomical research of the structure of corona and calyx allowed also to discover some distinctive features: the female flowers have smaller size of cells, they are characterized by larger indumentum by simple and ferriferous hairs and they carry larger amount of stomas and essence glands.
Текст научной статьи Морфологическое и анатомическое строение генеративных органов разных половых форм будры плющевидной {Glechoma hederacea L.)
Пермский государственный университет, 614990, Пермь, ул. Букирева, 15
Излагаются результаты сравнительного морфолого-анатомического изучения обоеполых и женских цветков Glechoma hederacea. Найдены отличия в размерах всех органов цветка, за исключением завязи. Сравнительно-анатомическое исследование строения венчика и чашечки позволило также выявить некоторые отличительные признаки: женские цветки имеют меньшие размеры клеток, характеризуются большим опушением простыми и железистыми волосками, большим количеством устьиц и эфирномасличных железок.
Будра плющевидная (сем. Lamiaceae) - много- Целью нашей работы явилось сравнительное летнее травянистое короткокорневищное растение с ползучими и часто укореняющимися в узлах боковыми побегами. Стебель 20-50 см длиной, четырехгранный. Листья по краю городчатые, почковидные. Встречается по тенистым берегам рек, в кустарниках, по опушкам лесов, на лугах и около жилищ, в редколесьях и лиственных лесах, на сорных местах у дорог (Куприянова, 1948; Флора СССР, 1954; Буданцев, 1992).
G. hederacea является лекарственным и медоносным растением. С лечебной целью используется надземная часть в период цветения. В траве содержится эфирное масло от 0.03-0.06 % (Лавре-нов, Лавренова, 1999) до 0.29 %, в цветках - витамин С. Эфирное масло используется как антисептическое и ранозаживляющее средство. Траву употребляют для ароматизации и приготовления тонизирующих напитков (Растительные ресурсы СССР, 1991; Дудченко, Козьяков, Кривенко, 1998).
Будра плющевидная является гинодиэцичным растением, т.е. в её популяциях встречаются обоеполые особи, имеющие только гермафродитные цветки, и женские с однополыми андростерильны-ми цветками (Knuth, 1899; Демьянова, 1997). Судя по литературным данным, анатомическое и морфологическое строение генеративных органов обоеполых и женских цветков у G. hederacea совершенно не изучено. В различных определителях и “Флорах” даже не указано наличие в популяциях женских особей, что в свою очередь может создавать затруднения при определении вида. Доля женских особей варьирует от 9.12 (Демьянова, Мухлынина, Козина, 1984) до 53.55% (по нашим данным для Пермского района Пермской области).
изучение морфологического и анатомического строения генеративных органов G. hederacea, выявление диагностических признаков разных половых форм.
Для исследования были взяты растения разных половых форм, произрастающие совместно в сходных экологических условиях на территории Пермской области. Для промеров использовали одновозрастные цветки как в свежем, так и в фиксированном в 70" спирте состоянии. Выборка составила от 10-30 переходных и до 50-100 обоеполых и женских цветков. Измерения околоцветника проведены с помощью миллиметровой бумаги с точностью до 0.5 мм, а измерения репродуктивных органов, в связи с их незначительными размерами, с помощью окуляр-микрометра с последующим пересчетом результатов в микрометры и миллиметры. Строение эпидермы генеративных органов изучено на временных препаратах, приготовленных по общепринятой методике (Прозина, 1960) с применением микроскопа МБИ-1. Рисунки репродуктивных органов и эпидермы околоцветника выполнены с помощью рисовального аппарата РА-6. Полученные данные обработаны методом вариационной статистики (Лакин, 1990).
Гинодиэция хорошо выражена в сем. Губоцветные (Lamiaceae). У них разница в размерах околоцветника обоеполых и женских особей проявляется вполне отчетливо (Демьянова, Титова, 1981), причем у G. hederacea размеры эти постоянны (Пономарев, Демьянова, 1975). Цветки G. hederacea собраны по 3-4 в пазухах средних и верхних стеблевых листьев. Прицветники короче цветоножек, нитевидные, 1-1.5 мм длиной (Куприянова,
Таблица 1
Сравнительные размеры частей цветка обоеполых и женских форм Glechoma hederacea L.
|
Признак |
Половая форма |
Пределы колебаний |
М±ш |
Cv, % |
td |
|
Длина цветка от основания до вершины верхней губы, см |
Обоеполая Женская |
1.50-2.10 0.72-1.25 |
1.77±0.02 0.98±0.01 |
9.04 10.20 |
35.33 |
|
Длина цветоножки, мм |
Обоеполая Женская |
1.2-2.0 0.8-1.8 |
1.70±0.03 1.27±0.05 |
14.12 25.98 |
7.37 |
|
Длина прицветника, мм |
Обоеполая Женская |
0.86-1.84 0.86-3.04 |
1.16±0.04 1.49±0.07 |
22.41 32.21 |
4.09 |
|
Длина трубки чашечки, мм |
Обоеполая Женская |
3.6-6.0 3.8-5.5 |
4.99±0.06 4.30±0.07 |
8.82 11.63 |
7.51 |
|
Длина зубцов чашечки, мм |
Обоеполая Женская |
1.2-1.8 1.0-1.8 |
1.35±0.02 1.27±0.03 |
11.11 14.96 |
2.22 |
|
Высота зева, мм |
Обоеполая Женская |
6.5-11.0 4.0-8.2 |
7.98±0.16 6.28±0.12 |
14.41 13.85 |
8.50 |
|
Длина тычиночной нити, см |
Обоеполая Женская |
1.15-1.8 0.52-0.78 |
1.55±0.02 0.63±0.01 |
9.03 14.29 |
41.14 |
|
Длина пыльника, мм |
Обоеполая Женская |
0.64-1.0 0.14-0.22 |
0.85±0.01 0.19±0.004 |
10.59 15.79 |
61.27 |
|
Длина завязи, мм |
Обоеполая Женская |
0.28-0.46 0.3-0.46 |
0.36±0.01 0.37±0.01 |
13.89 10.81 |
0.71 |
|
Длина столбика и рыльца, см |
Обоеполая Женская |
1.6-1.95 0.75-1.2 |
1.74±0.02 0.96±0.01 |
5.75 9.38 |
34.88 |
|
Длина нектарника, мм |
Обоеполая Женская |
0.5-0.76 0.4-0.56 |
0.59±0.01 0.47±0.01 |
8.47 10.64 |
8.49 |
|
Длина лопастей рыльца, мм |
Обоеполая Женская |
0.78-1.30 0.80-1.50 |
1.04±0.03 1.15±0.04 |
12.50 15.65 |
2.20 |
Примечание: Cv - коэффициент вариации, свидетельствующий о небольшом (0-10 %), среднем (11-20%), большом варьировании признака; td - критерий достоверности разности, если он равен или больше 3, разница считается достоверной.
1948; Флора СССР, 1954). Гермафродитные цветки оказались практически в 2 раза крупнее женских, для них же характерны большая высота зева (расстояние от средней лопасти нижней губы до верхней) (табл. 1; рис. 1).
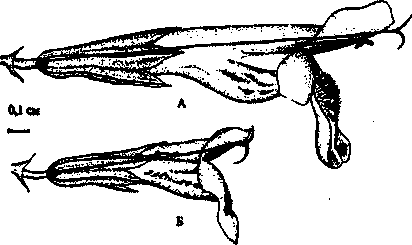
Рис. 1. Гинодиэция у Glechoma hederacea: А - обоеполый цветок; Б - женский цветок
Чашечка зеленая, узкая, трубчатая или колокольчатая, неясно двугубая, трубка прямая, в зеве скошенная (Буданцев, 1992), снаружи волосистая (Шишкин, 1963). Зубцы треугольно-ланцетные или линейно-ланцетные, внезапно тонко заостренные, в 3-4 раза короче трубки (Куприянова, 1948; Фло ра СССР, 1954; Буданцев, 1992). Каждый из 5 чашелистиков снабжен тремя основными проводящими пучками.
При морфологических измерениях нами учитывались длина трубки и зубцов чашечки отдельно. Данные табл. 1 свидетельствуют о большей длине трубки и зубцов чашечки у гермафродитных цветков.
Из литературных источников следует, что чашечка у G. hederacea изнутри голая (Буданцев, 1992). По нашим данным, напротив, внутренняя эпидерма опушена простыми одноклеточными остроконечными волосками, направленными к зубцам чашечки (рис. 3, 1Г). Среди них встречаются в небольшом количестве и железистые волоски (рис. 2, /). Устьица диацитного типа, равномерно распределены по внутренней эпидерме трубки чашечки, при продвижении же к зубцам их количество увеличивается.
Ребра чашечки с наружной стороны опушены 3-7-клеточными простыми волосками и железистыми волосками с 1- и 2-клеточной головкой (рис. 2; 2, 3, 4). В межреберье находятся головчатые волоски с 2-клеточной головкой и эфирномас-
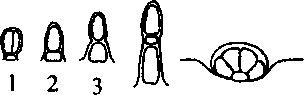
100 мкм 4 5
»-----------и
Рис. 2. Железистое опушение: 1-4 - железистые волоски; 5 - эфирномасличная железка личные железки (рис. 2; I, 5). Клетки эпидермы вытянуты в длину, со слегка извилистыми стенками, сверху покрыты складчатой кутикулой. Устьица диацитного типа (рис. 3; А, К).
Размеры клеток внешней и внутренней эпидермы чашечки гермафродитных цветков превышают таковые у женских в 1.3-1.5 раза. Количество простых волосков и устьиц на 1 мм2 больше у женских цветков. Аналогичные данные были получены нами ранее при изучении анатомического строения вегетативных органов G. hederacea, где было установлено, что большее количество устьиц находится на нижней эпидерме листьев женских растений (Анисимова, 2004). Большее количество устьиц может свидетельствовать о предрасположенности женских форм к более высокой интенсивности транспирации, чем обоеполых.
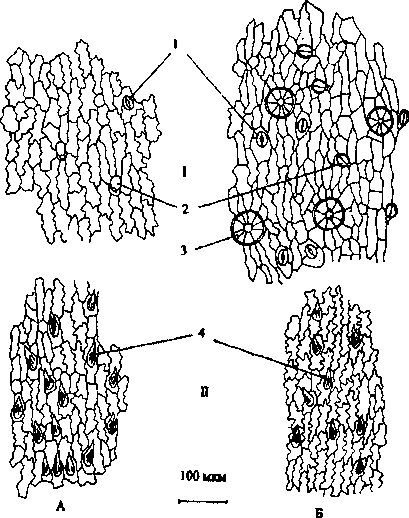
Рис. 3. Эпидерма чашечки обоеполых (Л) и женских (5) цветков: I- внешняя эпидерма; //- внутренняя эпидерма; 1 - устьице; 2 - железистый волосок; 3 - эфирномасличная железка; 4 - простой одноклеточный волосок
Особое внимание следует обратить на характер железистого опушения. Количество головчатых волосков на 1 мм2 эпидермы примерно одинаковое у обеих половых форм как на внешней, так и на внутренней эпидерме, с небольшим преобладани ем их у женских цветков. Несмотря на меньшие размеры чашечки женских цветков, количество эфирномасличных железок у них оказалось большим как в расчете на одно межреберье -18.94±1.39 (для сравнения: у обоеполых -13.00±0.82), так и на всей чашечке в целом -80.84±4.48 (у обоеполых - 60.80±4.89). Как известно, установлена прямо пропорциональная зависимость количественного содержания эфирного масла в сырье от количества железок на чашечке (Мухортова, 1977), а также на 1 мм2 на нижней эпидерме листа (Бубенчиков, Гримальская и др., 1998). Опираясь на эти сведения, можно предположить больший выход эфирного масла из растений женской формы.
Венчик фиолетово-синий, лиловый или розовый, редко - белый, от 10 до 18 мм длиной, обычно в 2-2.5 раза длиннее чашечки (Флора СССР, 1954; Шишкин, 1963; Буданцев, 1992). Основания лепестков образуют трубку, разделенную на дистальную (нижнюю) часть, состоящую из трех лепестков, и проксимальную (верхнюю) часть, включающую два других лепестка (Фегри, Пэйл ван дер, 1982). Трубка венчика длиннее чашечки, к отгибу постепенно расширенная. Проксимальная часть (верхняя губа) прямая, нижняя часть повернута наружу и вниз, образуя посадочную площадку для опылителя (Фегри, Пэйл ван дер, 1982). Центральная лопасть нижней губы выпуклая или плоская, выемчатая (Буданцев, 1992).
Внешняя эпидерма верхней и нижней губы состоит из изодиаметрических клеток с сильно извилистыми стенками. У обоеполых цветков размеры клеток больше, чем у женских, причем разница эта достоверна (рис. 4; I, II). Эпидерма опушена простыми 2-5-клеточными (у обоеполых) и 2-8клеточными (у женских) волосками, направленными вверх. Количество их на 1 мм2 больше у женских цветков - 30.72±1.28, у обоеполых - 23.24±1.48. Железистое опушение также более обильное у анд-ростерильных цветков - 18.72± 1.68 на 1мм2, у обоеполых- 10.08±1.11 на 1 мм2 эпидермы.
Устьица венчика, в отличие от устьиц чашечки и листьев (здесь они диацитные), имеют 4 (редко 5-8) побочных клеток, располагающихся крест накрест и имеющих размеры и очертания такие же, как и другие эпидермальные клетки (рис. 4, 7). Таким образом, можно предположить, что тип устьичного аппарата - аномоцитный (Баранова, 1985). Количество устьиц на 1 мм2 эпидермы больше у женских цветков.
При продвижении от отгиба к средней части трубки венчика и затем к ее основанию можно наблюдать увеличение размеров клеток в 3-3.5 раза. Если в средней части они имеют еще извилистые стенки, то в основании трубки венчика клетки прямоугольные, сильно вытянутые в длину. В средней части трубки венчика эпидерма опушена простыми 1, 2, 3-клеточными и редкими желези-
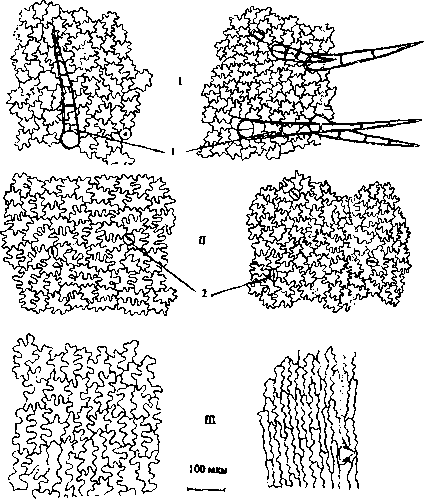
Рис. 4. Внешняя эпидерма венчика обоеполых (Л) и женских (5) цветков: /- верхняя губа; II- нижняя губа; 111- средняя часть трубки венчика; 1 - простой волосок; 2 - устьице
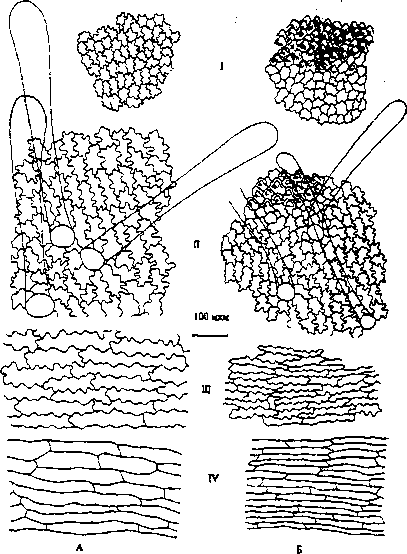
Рис. 5. Внутренняя эпидерма венчика обоеполых (Л) и женских (5) цветков: / - верхняя губа; 11 - нижняя губа; III - средняя часть трубки венчика
стыми волосками (рис.2; 2, 3, 4), причем количество и тех и других на 1 мм2 поверхности преобладает у женских цветков. У основания трубки венчика опушение отсутствует.
На внутренней эпидерме венчика устьица и железистые волоски отсутствуют. Размеры клеток сохраняют тенденцию к увеличению у обоеполых цветков. В выпуклой части центральной лопасти нижней губы (в месте перехода нижней губы в трубку венчика) располагаются многочисленные простые волоски, в верхней части - булавовидно утолщенные (рис. 5, 77). Длина волосков достигает 0.5-0.7 мм, они различимы невооруженным глазом. Количество данных волосков на 1 мм2 больше у женских цветков. При рассмотрении цветков под микроскопом обнаружено, что на волосках в основании нижней губы находится большое количество пыльцы. .
Таким образом, чашечка и венчик женских цветков имеют меньшие размеры, в связи с этим и меньшие размеры клеток внешней и внутренней эпидермы. Эти цветки более опушенные простыми, железистыми волосками и эфирномасличными железками, они имеют большее количество устьиц и клеток в расчете на 1 мм2 поверхности.
Андроцей G. hederacea состоит из 4 тычинок с длинными параллельными тычиночными нитями, срастающимися с трубкой венчика. Тычинки короче венчика, пыльники гладкие, расходящиеся под прямым углом (Буданцев, 1992). Они располагаются под верхней губой и защищены от дождя. У женских цветков тычинки рудиментарны, они поч- ти неразличимы невооруженным глазом (табл.1). Пыльники мелкие, лишены морфологически оформленной пыльцы (рис. 6).
Гинецей G. hederacea, как и у всех представителей сем. Губоцветные, формируется двумя плодолистиками. Завязь верхняя, ложночетырехгнездная. Размеры ее у обоеполых и женских цветков почти одинаковые. Длина столбика больше у гермафродитных, а длина рыльца, напротив, у женских цветков (табл.1).
Нектарник у всех губоцветных является производным основания плодолистиков, имеет вид диска, расположенного на нижней части завязи. Диск имеет 4 зубца, которые чередуются с лопастями завязи. Наиболее мощного развития и наиболее сильной дифференцировки достигает передний зубец; задний зубец, обращенный к верхней губе, развит значительно слабее как в отношении размера, так и по ряду анатомических признаков (Карташова, 1960; 1965). Боковые зубцы не отличаются по величине от заднего зубца. У G. hederacea нектарники крупных гермафродитных цветков несколько больше по размерам, нежели у женских (табл. 1; рис. 7, Л).
У данного вида гинодиэции сопутствует гино-моноэция. В этом случае в популяции встречаются гермафродитные, женские особи и отдельные растения, имеющие обоеполые и женские цветки. Доля женских цветков у таких особей составляет не более 5%. Гиномоноэцичные растения являются в сущности обоеполыми, у них тенденция к мужской
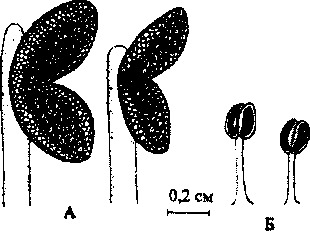
Рис. 6. Строение тычинок обоеполого (Л) и женского (Б) цветка
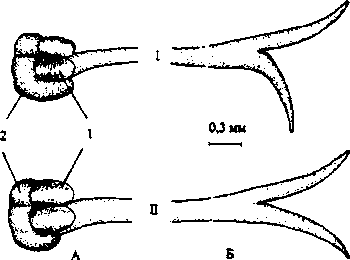
Рис. 7. Завязь (Л) и рыльце (Б) обоеполого (7) и женского (77) цветка: 7 - завязь; 2 - нектарник
стерильности «захватила» не все, а лишь некоторую часть цветков (Пономарев, Демьянова, 1975).
Таблица 2
Сравнительные размеры частей обоеполого и переходных цветков с разной степенью редукции андроцея
|
Признак |
Обоеполый цветок |
Переходные цветки с редуцированными тычинками |
|
|
одной |
двумя |
||
|
Длина цветка от основания до вершины верхней губы, см |
1.77±0.02 |
1.65±0,08 |
1.59±0.09 |
|
Длина цветоножки, мм |
1.70±0.03 |
1.67±0.07 |
1.65±0.05 |
|
Длина трубки чашечки, мм |
5.10±0.01 |
4.99±0.06 |
4.81±0.01 |
|
Длина зубцов чашечки, мм |
1.41±0.02 |
1.40±0.02 |
1.33±0.01 |
|
Высота зева, мм |
7.98±0.16 |
7.75±0.25 |
6.75±0.25 |
|
Длина тычиночной нити, см |
1.55±0.02 |
1.53±0.01 |
1.33±0.18 |
|
Длина пыльника, мм |
0.85±0.01 |
0.79±0.01 |
0.78±0.02 |
|
Длина завязи, мм |
0.36±0.01 |
0.33±0.05 |
0.32±0.04 |
|
Длина столбика и рыльца, см |
1.74±0.02 |
1.70±0.00 |
1.55±0.10 |
|
Длина нектарника, мм |
0.59±0.01 |
0.60±0.00 |
0.56±0.00 |
Кроме обоеполых и женских цветков на отдельных особях G. hederacea встречаются переходные цветки. При этом у одних цветков рудиментарна одна, у других - две тычинки с полностью редуцированными пыльниками, не отличающиеся от тычинок женских цветков. Фертильные тычинки имеют меньшую длину тычиночной нити и пыльников, то же касается и размеров цветка в целом. Переходные цветки по размерам занимают промежуточное положение между обоеполыми и женскими. Причем размеры зависят от количества редуцированных тычинок - чем больше их количество в цветке, тем меньше параметры: длина цветка, чашечки, тычиночных нитей, пыльников, завязи и столбика (табл. 2).
Ч. Дарвин утверждал, что “уменьшение размеров венчиков женских цветков обусловлено тенденцией к редукции, перешедшей с тычинок на лепестки” (Дарвин, 1948). Эта мысль высказана и в гормональной теории Бейкера (Baker, 1957) и Плака (Plack, 1957). Плак (Plack, 1957) предположил, что рост венчика находится под влиянием гормона, продуцируемого в пыльниках. Он экспериментально доказал это, произведя кастрацию в обоеполых бутонах Glechoma и удаляя от 1 до 4 пыльников. Размеры венчиков кастрированных цветков уменьшались соответственно количеству удаленных пыльников (Пономарев, Демьянова, 1975). Таким образом, гормональная теория не вскрывает причин гинодиэции, но удовлетворительно объясняет уменьшение размеров венчиков женских цветков и переходных цветков с частичной редукцией андроцея (Демьянова, Титова, 1981).
Выводы
В результате статистической обработки материала были обнаружены достоверные различия в размерах всех исследованных органов цветка, за исключением длины завязи. Меньшие размеры женских цветков удовлетворительно объясняет гормональная теория, предполагающая зависимость роста венчика от гормона, продуцируемого в пыльниках. Встречающиеся на отдельных особях переходные цветки имеют промежуточные размеры между обоеполыми и женскими цветками, связанные с числом редуцированных тычинок, что также подтверждает гормональную теорию.
В связи с меньшими размерами венчика и чашечки женские цветки имеют меньшие размеры клеток, чем гермафродитные цветки. Женские цветки более густо опушены простыми и железистыми волосками, у них же преобладает количество устьиц на 1 мм2, что может свидетельствовать о большей интенсивности транспирации андросте-рильных растений. Чашечка у этих цветков содержит почти в 1.5 раза больше эфирномасличных железок, чем у обоеполых. Опираясь на эти данные, можно предположить больший выход эфирного масла из растений женской формы.
Таким образом, практически все исследованные нами признаки у обоеполых и женских цветков имеют достоверные различия и могут быть использованы при диагностике содержания эфирных масел.
Список литературы Морфологическое и анатомическое строение генеративных органов разных половых форм будры плющевидной {Glechoma hederacea L.)
- Анисимова А.Г. Анатомическое строение вегетативных органов разных половых форм будры плющевидной {Glechoma hederacea L.) II Вести. Перм. ун-та. 2004. Вып. 2. Биология. С. 44-52.
- Баранова М.А. Классификация морфологических типов устьиц//Бот жури. 1985. Т.70, № 12. С. 1585-1595.
- Бубенчиков А.А., Гргшальская СИ., Бубенчиков Р.А. Влияние морфолого-анатомических показателей на выход эфирного масла из растений рода чабрец//Человек и его здоровье/Курск, гос. мед. ун-т 1998. № 1. С. 299-301.
- Буданцев А.Л. Конспект трибы Nepeteae (Lamiaceae). Роды Meehania, Glechoma, Drepanocaryum, Marmoritis и Hymenocrater II Бот журн. 1992. Т. 77, №12. С. 118-128.
- Дарвин Ч. Различные формы цветков у растений одного и того же вида//Соч. Т. 7. М., 1948. 650 с.
- Демьянова Е.И. Половая структура природных популяций сексуально-полиморфных растений Предуралья.//Вести. Перм. ун-та. 1997. Вып. 3. Биология. С.9-20.
- Демьянова Е.И., Мухлынина Э.Н., Козина Т.А. Половая структура природных популяций гиноди-эцичных и двудомных растений Хомутовской степи//Экология опыления растений. Пермь, 1984. С. 15-25.
- Демьянова Е.И., Титова А.В. Морфология и размеры цветков разных половых типов у гиноди-эцичных растений//Экология опыления. Пермь, 1981. С. 3-20.
- Дудченко Л.Г., Козьяков А.С., Кривенко ВВ. Пряно-ароматические и пряно-вкусовые растения: Справочник. Киев: Наук, думка, 1989. 304 с.
- Карташова Н.Н. Некоторые данные по морфологии цветка губоцветных (Labiatae)//Бот. журн. 1960. Т. 45, № I.e. 109-114.
- Карташова Н.Н. Строение и функция нектарников цветка двудольных растений. Томск, 1965. 195 с.
- Куприянова Л.А. О роде Glechoma L. и его видах//Бот. журн. 1948. Т. 33, № 2. С. 230-238.
- Лавренов В.К., Лавренова Г.В. Полная энциклопедия лекарственных растений. СПб.: Нева; М.: ОЛМА-ПРЕСС, 1999. T.I.
- Лакин В.Ф. Биометрия. М.: Высш. шк., 1990.
- Мухортова Т.Г. Анатомическое строение надземных органов гибрида лаванды -лавандина//Раст ресурсы. 1977. Т. 15, вып. 1.С. 83-90.
- Пономарев А.Н., Демьянова Е.И. К изучению гинодиэции у растений//Бот. журн. 1975. Т.60, № I.e. 3-15.
- Прозина М.Н. Ботаническая микротехника. М, 1960. Растительные ресурсы СССР: Цветковые растения, их химический состав, использование: Семейства Hippuridaceae-Lobeliaceae. СПб.: Наука, 1991.200 с.
- Фегри К. Л., Пэйл ван дер. Основы экологии опыления. М., 1982.
- Шишкин Б.К Labiatae//Флора СССР. М.; Л.: Изд-во АН СССР, 1954.Т.20,21.
- Шишкин Б.К. Ботанический атлас. М.; Л., 1963.
- Baker Н. J. Expression of sex in flowering plants. Mature, 1957, V. 180, N 4586. P. 614-615.
- Knuth P. Handbuch der Blutenbiologie. Leipzig: Verlag von Wilhelm Engelmann. 1899. Bd II, T. II. 705 S.
- Plack A. Sexual dimoфhism in Labiatae II Nature, 1957. V.180,N4596.P. 1218-1219.
