Морфология чешуи и рост сига Coregonus lavaretus (Coregonidae) Йокостровской Имандры
 Йокостровской Имандры")
Автор: Зубова Елена Михайловна, Кашулин Николай Александрович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 1 т.17, 2014 года.
Бесплатный доступ
Рассмотрены размерно-возрастные особенности морфологического строения чешуи сига Сoregonus lavaretus L. Йокостровской Имандры, заполярного интенсивно загрязняемого водоема. Показана неравномерность относительной скорости роста различных секторов данной минерализованной структуры в течение жизни при постоянстве ее формы. Для обратных расчислений длины рекомендуется использовать передний диагональный радиус чешуи с применением регрессионного метода. Анализ динамики темпов роста сига показал, что самый высокий темп характерен для первого года жизни, затем идет снижение значений как средних годовых приростов, так и удельной скорости роста рыб. Описано влияние цикла полового созревания у сига на темп его роста.
Сиг, оз. имандра, морфология чешуи, обратные расчисления, темп роста
Короткий адрес: https://sciup.org/14294662
IDR: 14294662
Текст научной статьи Морфология чешуи и рост сига Coregonus lavaretus (Coregonidae) Йокостровской Имандры
Исследования изменения рыбной части сообщества оз. Имандра, испытывающего многофакторное антропогенное влияние, проводятся уже более 70 лет ( Крогиус , 1931; Галкин и др ., 1966; Беляева и др ., 1972; Решетников , 1980; Моисеенко , 1983; 1997; 2000; 2002; Моисеенко, Яковлев , 1990; Моисеенко, Лукин , 1999; Антропогенные... , 2002; Лукин и др ., 2006; Решетников и др ., 2011;
Решетников, Попова , 2012; Терентьев, Кашулин , 2012 и др.). Основное внимание в исследованиях уделяется ее доминирующему комплексу – сиговым рыбам, в частности сигу Coregonus lavaretus , который рекомендован как тест-объект ихтиологического мониторинга ( Моисеенко , 1991).
При описании популяционных характеристик сига озера большое внимание всегда уделялось изменению ростовых характеристик рыб, которые касались не только наблюдаемых размерно-весовых параметров, но и расчисленной длины, что позволяло реконструировать особенности ростовых характеристик изучаемых рыб за период их жизни (Крогиус, 1931; Владимирская, 1956; Решетников, 1966; Беляева и др., 1972; Моисеенко, 2002; Зубова, 2013; Зубова, Кашулин, 2013). Однако в представленных работах методические особенности обратных расчислений длины сигов оз. Имандра приводились только Ю.С. Решетниковым (1966; 1980). Вместе с тем данная процедура имеет ряд особенностей, которые, во-первых, заключаются в выборе регистрирующей структуры, объективно отражающей начало и конец периода роста (Дгебуадзе, 1979), во-вторых, в выборе оптимального участка или сектора данной структуры, что достигается подробным изучением ее морфологии и размерновозрастной изменчивости (Вовк, 1956; Брюзгин, 1969; Сasselman, 1990; Хуршут, 2003). Также исследование темпа роста рыб требует более точного выбора метода обратных расчислений и дальнейшей его характеристики (Шмальгаузен, 1935; Чугунова, 1959; Брюзгин, 1969; Мина, Клевезаль, 1976; Дгебуадзе, 1979; 2001; Хуршут, 2000; 2003).
Для сиговых рыб умеренной и субарктической зоны северного полушария наиболее часто используемой структурой для обратных расчислений является чешуя ( Van Oosten , 1923; Решетников , 1966; 1980; Черешнев, Скопец , 1992; Смирнов, Смирнова-Залуми , 1993). Особенности закладки годового кольца на чешуе у сигов бассейна оз. Имандра были подробно описаны Ю.С. Решетниковым (1966;
1980), но морфологические особенности чешуи не приводились. Есть неаргументированные указания на то, что темпы роста расчислялись по переднему радиусу чешуи ( Решетников , 1966; 1980).
Вместе с тем чешуя как регистрирующая структура может отражать не только особенности роста рыб, но и особенности условий их обитания. Интенсивное загрязнение водоема в последние десятилетия сказывается на нарушениях развития и строения внутренних органов и структур костного и кожного скелета, в том числе и чешуи, что затрудняет ее использование для определения возраста и исследования темпов роста. Это обусловливает необходимость уточнения морфологии чешуи и методик ее использования в размерно-возрастных расчислениях.
В работах по темпу роста сига оз. Имандра реконструкция длины проводилась по методу Э. Леа. При этом в полученных результатах всегда проявлялся "феномен" Розы Ли, что затрудняет сравнение разновозрастных группировок сига ( Мина , 1981). Также нельзя оценить статистическую значимость различий между данными из различных источников, так как авторы не приводили оценок статистической точности расчетов.
В вышеуказанных работах по темпу роста сига, в основном, приводятся абсолютные количественные показатели роста рыб, в то время как более удачным при сравнительных исследованиях роста оказывается применение относительных показателей ( Шмальгаузен , 1935; Чугунова , 1959; Мина, Клевезаль , 1976; Дгебуадзе , 1979; 2001).
Для решения этих проблем в рамках комплексного обследования оз. Имандра в 2011-12 гг. нами были исследованы преднерестовые скопления сига одного из плесов данного водоема – Йокостровской Имандры. Цель данной работы – уточнение методов реконструкции и характеристики роста сигов оз. Имандра за период их жизни с учетом современных условий обитания. В ходе работы решались следующие задачи: исследование современных особенностей морфологического строения чешуи сигов с выбором наиболее оптимального сектора чешуи для обратных расчислений длины; выбор оптимальной методики обратного расчисления длины для исследуемых сигов.
2. Материал и методы
Материал в 2011-12 гг. собран в летне-осенний период в проливе между мысом Кукисньярк и островом Нурксуол (Большой Йокостровский), проливе Узкая Салма и районе губы Глубокая плеса Йокостровская Имандра (рис. 1).
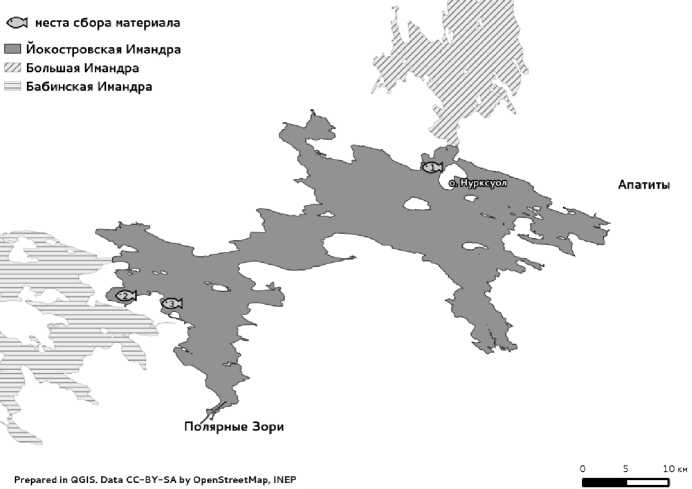
Рис. 1. Карта-схема сбора материала в плесе Йокостровская Имандра, 2011-12 гг.:
1 – пролив между мысом Кукисньярк и островом Нурксуол (Большой Йокостровский),
2 – пролив Узкая Салма, 3 – р-н губы Глубокая
Рыбу отлавливали ставными жаберными донными сетями из нейлонового монофиламента стандартной длиной 25 м, высотой 1.5 м и размером ячеи 10, 12.5, 16, 22, 25, 30, 35, 38 и 45 мм, что обеспечивало вылов рыбы длиной от 5 см и более. Обработка материала проводилась по стандартной методике ( Правдин , 1966). Для выделения внутривидовых форм у исследуемых сигов производился подсчет количества тычинок на первой жаберной дуге ( Правдин , 1966; Решетников , 1980; Siwertsson et al. , 2008).
В выборке было 243 экземпляра сига. Чешуя для исследования всегда бралась с одного и того же участка - с середины тела, под передней частью спинного плавника. Для определения возраста просматривалось под бинокуляром 4-6 чешуй от каждого сига (Галкин, 1958; Чугунова, 1959; Зиновьев, Мандрица, 2003; Чернова, Дгебуадзе, 2008). У 173 рыб выборки исследованы морфологические особенности чешуи с дальнейшим выбором оптимального сектора чешуи для обратных расчислений длины. Для этого с помощью окуляр-микрометра проводились дополнительные измерения ее продольного и поперечного диаметров, а также радиусов: переднего (базального) и переднего диагонального. Боковой и задний (каудальный) радиусы не измерялись; их находили путем деления значения поперечного диаметра на 2 и вычитания из значения продольного диаметра значения переднего радиуса соответственно (рис. 2). На основе этих измерений были определены:
J - относительный размер чешуи, равный продольному диаметру ( ab ) в процентах от длины тела ( АС);
В - отношение продольного диаметра ( ab ) к поперечному ( cd) - характеризует форму (ширину)
чешуи;
br - передний радиус ( bo ), выраженный в процентах от продольного диаметра чешуи ( ab ), -показывает положение ядра чешуи (рис. 2) ( Галкин , 195 8; Чернова , Дгебуадзе , 2008).
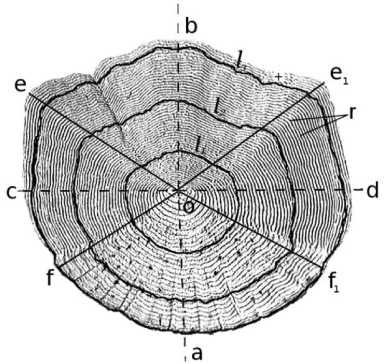
Рис. 2. Схема структуры чешуи трехгодовалого (четырехлетнего) сига Coregonus lavaretus плеса Йокостровская Имандра, 2011-12 гг. (по Галкину , 1958, с изменениями): о - центр, ab - продольный диаметр, cd - поперечный диаметр, bo - передний (базальный) радиус, eo и e 1 о - передние диагональные радиусы, со и do - боковые радиусы, fo и f _ о - задние диагональные радиусы, оа - задний (каудальный) радиус, еое 1 - передний сектор, ecf и ecf 1 - боковые сектора, fof 1 - задний сектор, r - склериты, 1 1 , 1 2, 1 3 -годовые зоны роста (по Смирнову, Смирновой-Залуми , 1993), + - незаконченный прирост
Для описания изменения секторов чешуи (переднего, бокового и заднего) относительно высоты тела рыбы с возрастом измеряли наибольшую высоту тела сигов ( qh ), мм ( Вовк , 1956; Чугунова , 1959).
Подсчет склеритов каждого годового кольца и незаконченного прироста у исследуемых рыб выборки проводили по переднему радиусу чешуи.
Обратные расчисления были сделаны для 243 рыб. Расчисления велись по формуле Розы Ли ( Чугунова , 1959; Брюзгин , 1969):
L i = Lc ( 5/5 с) + a(1 - 5 XZ 5 c ), (1)
где 5 c - радиус чешуи данной рыбы в момент ее поимки; 5 i - радиус чешуи этой особи в возрасте i лет; Lc - длина рыбы в момент поимки; L i - длина рыбы в возрасте i лет; а - постоянная величина. Для характеристики темпов роста использована формула удельной скорости роста Шмальгаузена-Броди, учитывающая нарастание длины по сложным процентам ( Шмальгаузен , 1935; Мина, Клевезаль , 1976; Дгебудзе , 2001):
с = lg l 2- lg l 1 ,
l 0.4343( t 2 t 1)
где С i - удельная скорость роста; 1 1 , 1 2 - значения удельной скорости роста; 0,4343 - модуль перехода от натуральных логарифмов к десятичным; t 1 , t 2 - показатели времени от начала роста рыбы (от выхода из икры) до начала и конца отрезка времени, за который вычисляется удельная скорость роста. Описание периодов роста проводилось по формуле константы роста И.И. Шмальгаузена, предполагающей, что произведение удельной скорости роста на время, протекшее с начала роста, является постоянной величиной для отдельных периодов роста, на которые распадается развитие всякого живого ( Шмальгаузен , 193 5; Чугунова , 1959 ; Мина, Клевезаль , 1976):
lg l 2 lg l 1 t 2 t 1
C , lt 0.4343( t t ) 2
где C1t - константа роста; 1 1 , 1 2, 0,4343, t 1 , t 2 - как к формуле (2).
3. Результаты и обсуждение
В наших уловах сиги в основном были представлены одной формой – малотычинковой: с числом жаберных тычинок на первой дуге от 15 до 31 (в среднем 23.1 ± 0.2), длина наибольшей тычинки в среднем составила 10.3 ± 0.2 % от длины жаберной дуги ( Решетников , 1980; Siwertsson et al ., 2008). Преднерестовые скопления сига были представлены особями в возрасте от 1+ до 7+ лет, длиной 161-460 (в среднем 263.8 ± 3.6) мм и массой 44-1660 (249.2 ± 12.5) г.
-
3.1. Морфологические особенности чешуи
Чешуя малотычинкового сига циклоидная. По нашим наблюдениям, центральный склерит имеет овальную форму, чаще незамкнут; область центра содержит включения, которые напоминают участки разрушенного склерита (рис. 3а). Склериты имеются на всей поверхности чешуи, у исследуемых рыб они образуют чередующиеся комплексы из "концентрических" и следующих за ними "выклинивающихся" ("срезанных") склеритов, которые формируют годовые зоны роста (рис. 3а) ( Решетников , 1980; Смирнов , Смирнова-Залуми , 1993). Выклинивание склеритов начинается на границе бокового и заднего секторов чешуи и заканчивается на переднем секторе, поэтому здесь их большее число (рис. 3а). Это говорит о закладке и более интенсивном их росте в базальной части с вершины чешуи ( Галкин , 1958). Также в переднем секторе чешуи наблюдается удвоение склеритов (рис. 3а, б). В переднем секторе склериты слабо волнистые ( Галкин , 1958; Чернова , Дгебуадзе , 2008). Вдоль боковых секторов чешуи в результате утери склеритов расстояние между ними незначительно больше. Гораздо большие расстояния между склеритами наблюдаются на заднем секторе (рис. 3а), что, в первую очередь, объясняется дополнительной потерей некоторых склеритов на границе бокового и заднего секторов чешуи, они как бы "обрываются" (рис. 3а, в). Также боковой и задний радиусы чешуи у исследуемых рыб в первые два года имеют большие относительные размеры, по сравнению с передним радиусом (табл. 1).

Рис. 3. Морфологические особенности чешуи (а), удвоение склеритов на переднем секторе чешуи (б) и "обрывание" склеритов на границе бокового и заднего секторов чешуи (в) малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, в возрасте 3+ ( АС = 265.0 мм), 2012 г. ОЦ – область центра, ПС – передний сектор, БС – боковой сектор, ЗС – задний сектор. Увеличение: а – ок. 10 ×, об. 5 ×, б, в – ок. 10 ×, об. 20 ×
Учитывая все вышеописанные особенности в распределении склеритов на чешуе малотычинкового сига Йокостровской Имандры, их подсчет правильнее вести по переднему радиусу.
У 98 % исследуемых рыб во взятой пробе чешуи встречалась чешуя с разрушенной центральной областью (рис. 4), она составляла от 50 до 70 % от пробы. Такой картины не наблюдалось у малотычинкового сига из условно-фоновых районов оз. Имандра – Бабинской Имандры и других водоемов фоновой зоны региона. Это, возможно, говорит о более неблагоприятных экологических условиях плеса Йокостровская Имандра.
В данном плесе также у 15 % выловленных сигов наблюдалось ерошение чешуи (рис. 5). Как известно, ерошение чешуи является основным симптомом при таких бактериальных заболеваниях рыб, как аэромоноз, лепидортоз и т.д. ( Чернова, Дгебуадзе , 2008) и, возможно, свидетельствует о неблагоприятном бактериологическом режиме воды исследуемого плеса и ослабленном иммунитете рыб.


Рис. 4. Чешуя с разрушенной центральной областью малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра в возрасте 2+ ( АС = 232.0 мм), 2011-12 гг.
Увеличение: ок. 10 ×, об. 1.8 ×
Рис. 5. Ерошение чешуи на теле малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра (фотография П.М. Терентьева, 2012 г.)
-
3.2. Размерно-возрастные изменения морфологии чешуи
Относительный размер чешуи исследуемых рыб достоверно увеличивается с возрастом, при этом центр чешуи сдвигается к заднему (каудальному) краю ( P > 0.95) (табл. 1). Эти изменения происходят из-за различного роста переднего и заднего секторов чешуи сига в разном возрасте. Так, передний сектор чешуи, включающий передний и передний диагональный радиусы как относительно наибольшей высоты ( qh ), так и длины тела ( АС ), с возрастом увеличивается, при этом задний сектор, включающий, в нашем случае, измерения заднего радиуса – уменьшается ( P > 0.95) (табл. 2). Такие изменения заднего сектора чешуи с возрастом наиболее очевидны относительно наибольшей высоты тела исследуемых рыб.
Таблица 1. Относительный размер чешуи ( J ), %, форма чешуи ( В ) и относительное положение ядра чешуи ( br ), % малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, 2011-12 гг.
|
Возраст, лет |
J |
br |
B |
|
1+ |
1.38 ± 0.06 (12) |
43.4 ± 1.0 (12) |
0.96 ± 0.01 (12) |
|
2+ |
1.46 ± 0.07 (9) |
47.3 ± 1.4 (9) |
0.95 ± 0.03 (9) |
|
3+ |
1.60 ± 0.03 (51) |
51.4 ± 0.6 (51) |
0.97 ± 0.01 (51) |
|
4+ |
1.65 ± 0.03 (39) |
54.9 ± 1.4 (39) |
0.99 ± 0.03 (39) |
|
5+ |
1.68 ± 0.03 (49) |
52.7 ± 0.6 (49) |
0.99 ± 0.02 (49) |
|
6+ |
1.81 ± 0.04 (9) |
57.1 ± 0.9 (9) |
0.98 ± 0.03 (9) |
|
7+ |
1.78 ± 0.15 (4) |
59.9 ± 2.9 (4) |
1.0 ± 0.08 (4) |
Примечание. В таблице представлено среднее значение, его ошибка и число рыб (в скобках).
Что же касается бокового сектора чешуи сига, включающего боковой радиус, его размеры относительно наибольшей высоты тела увеличиваются до возраста 4+, затем начинают уменьшаться, относительно длины тела – они увеличиваются до возраста 6+ (табл. 1).
В результате роста рыбы увеличивается поверхность, которую надо покрыть чешуей. Естественно, что увеличивается передний сектор чешуи, который располагается внутри чешуйного кармана. В нашем случае, это можно отнести и к боковому сектору, который у исследуемых сигов располагается также внутри чешуйного кармана (рис. 6). Рост разных частей чешуи может также зависеть от эпидермиса вне чешуйного кармана, который бывает сильно развит ( Чернова, Дгебуадзе , 2008).
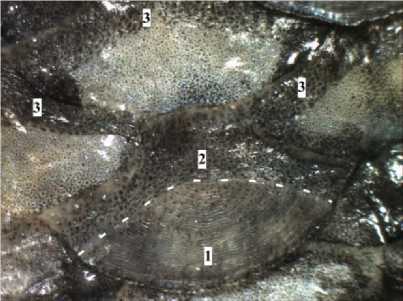
Рис. 6. Расположение чешуи в чешуйном кармане малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра в возрасте 6+ ( АС = 464.0 мм), 2012 г. 1 – задний (каудальный) сектор чешуи, 2 – чешуйный карман, 3 – чешуйные карманы прилегающих чешуй.
Увеличение: а, б – ок. 10 ×, об. 1 ×
Несмотря на описанные изменения роста секторов чешуи, форма чешуи ( В ) исследуемых сигов остается постоянной на протяжении всей жизни рыб, т.е. пропорции продольного диаметра чешуи к поперечному диаметру с возрастом практически не меняются (табл. 1).
В основе обратных расчислений длины лежит утверждение о наличии связи между длиной тела и размерами чешуи. Мы определили корреляцию размеров чешуи по рассматриваемым радиусам с длиной ( АС ) малотычинкового сига. Из табл. 3 видно, что наиболее сильно связаны с длиной тела размеры чешуи по переднему и переднему диагональному радиусу. У исследуемых сигов во всех возрастах по переднему и переднему диагональному радиусу чешуи число годовых колец одно и то же, поэтому мы выбираем тот радиус для обратных расчислений, по которому связь "длина рыбы – размер чешуи" самая высокая. В нашем случае, это передний диагональный радиус ( r = 0.91). Также анализ исследованной нами чешуи показал, что на переднем диагональном радиусе чешуи возможно более четкое фиксирование годовых зон у исследуемых рыб (рис. 7).
Таблица 3. Коэффициенты корреляции между длиной ( АС ), мм тела и размерами чешуи по различным радиусам малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, 2011-12 гг.
|
Радиусы |
r |
n |
|
Передний |
0.90 |
173 |
|
Передний диагональный |
0.91 |
173 |
|
Боковой |
0.83 |
173 |
|
Задний |
0.89 |
173 |

Рис. 7. Годовые зоны роста ( l 1 , l 2 , l 3 ) в районе переднего диагонального радиуса чешуи малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, в возрасте 3+ ( АС = 265.0 мм), 2012 г. Увеличение: ок. 10 ×, об. 5 ×
-
3.3. Рост
Зависимость между размерами тела ( АС ) и чешуи малотычинкового сига лучше описывается уравнением линейной регрессии Ln = 111.60 + 5.92 Rn ( n = 243) (рис. 8). Линия регрессии не проходит через начало координат, поэтому предпочтительнее использовать для обратных расчислений длины формулу Р. Ли (метод регрессии) ( Брюзгин , 1969). Отсюда находим формулу для обратного расчисления длины рыб: Li = 111.60 + ( Ln – 111.60) × ( Ri/Rn ). Результаты обратных расчислений длины по регрессионному методу приведены в табл. 4, где "феномен" Розы Ли не проявляется.
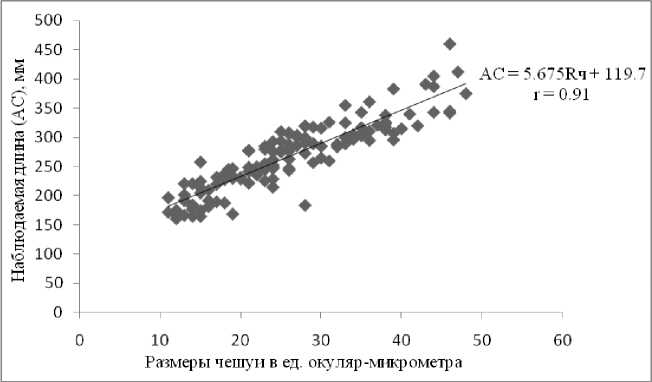
Рис. 8. Линейная зависимость длины тела ( АС ) малотычинкового сига
C. lavaretus lavaretus плеса Йокостровская Имандра от размеров переднего диагонального радиуса чешуи, 2011-12 гг.
По нашим данным, у исследуемых сигов с 1 по 5 год отсутствует компенсаторный рост, так как изменчивость в длине тела рыб с возрастом увеличивается (табл. 5), что, возможно, приводит к неодновременному наступлению созревания рыб. Значения коэффициента вариации снижаются на 6 и 7-й год жизни – после массового нереста сигов (см. ниже).
Cамый высокий темп роста был характерен для первого года жизни сигов (табл. 4). Начиная со второго года, идет снижение значений как средних годовых приростов, так и удельной скорости роста рыб. То же самое можно сказать и о количестве сформированных склеритов в каждом годовом кольце у исследуемых сигов (табл. 6).
У изученных нами сигов половые различия в темпе роста не наблюдались (табл. 7). Значения константы роста по Шмальгаузену ( Сlt ) у обоих полов увеличиваются с 1 по 5-й год (табл. 7), что, возможно, соответствует периоду быстрого роста сигов ( Шмальгаузен , 1935; Чугунова , 1959). Затем происходит постепенное снижение значений константы роста (период медленного роста). При этом созревающие самцы сига наблюдались в выборке в возрасте от 3+ и до 6+, самки – от 4+ до 7+, массовый нерест происходил в возрасте 4+ и 5+ соответственно.
Таким образом, выявить влияние наступления полового созревания на темпы линейного роста малотычинковых сигов Йокостровской Имандры при сравнении усредненных показателей сложно из-за неодновременного растянутого созревания и неежегодности нереста ( Решетников , 1966; 1980; Решетников, Богданов , 2011; Шарова, Лукин , 2000). Поэтому мы проанализировали раздельно особенности роста самцов сига в возрасте 3+, 4+ и самок сига в возрасте 4+ и 5+, готовящихся к нересту в текущем году, подразумевая, что они созревают впервые, и пропускающих его (табл. 8).
Готовящиеся к нересту самцы в возрасте 3+ росли достоверно быстрее не нерестящихся на протяжении всей жизни. В итоге особи, готовящиеся к нересту, имели большие наблюдаемые длины по сравнению с не нерестящимися (табл. 8). В возрасте 4+ рост участвующих и не участвующих в нересте сигов в основном достоверно не отличался (табл. 8), что также характерно и для самок сига в возрасте 5+. Среднее количество склеритов текущего года у впервые нерестящихся сигов во всех возрастах достоверно не отличалось от количества склеритов у не нерестящихся рыб (табл. 8).
Таким образом, прямых данных о замедлении темпов роста в связи с созреванием гонад у сигов из исследуемых выборок нет, что также отмечалось Ю.С. Решетниковым (1966) для сигов малокормных озер Лапландского заповедника. Он указывал на наличие у самцов и самок сига, после массового их созревания, чередования годов быстрого и медленного роста. У сигов плеса Йокостровская Имандра 2011-12 гг. чередования годов быстрого и медленного роста не наблюдается. Это, возможно, связано с тем, что большая часть участвующих в нересте (III-IV стадия развития гонад) экземпляров были впервые нерестящимися, и в условиях эвтрофирования озера обеспеченность пищей достаточна для быстрого восполнения энергетических затрат.
При исследовании впервые нерестящихся сигов была выявлена закономерность: чем выше темпы роста рыб, тем в более раннем возрасте они созревают (табл. 8). Описываемая зависимость является одной из форм связи между темпом роста и скоростью полового созревания, описанная у рыб в природных условиях ( Никольский , 1956; Земская , 1964; Кошелев , 1971; Решетников , 1980; Trudel et al ., 2000).
Вышеописанные особенности и закономерности темпов линейного роста сига Йокостровской Имандры близки к таковым малотычинкового сига из условно-фоновых районов оз. Имандра – Бабинской Имандры ( Зубова, Кашулин , 2013).
Таблица 5. Значения коэффициента корреляции расчисленной длины ( АС ) малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра в разном возрасте, 2011-12 гг.
|
– |
l 1 |
l 2 |
l 3 |
l 4 |
l 5 |
l 6 |
l 7 |
|
Cv |
6.45 |
9.54 |
11.52 |
11.96 |
14.16 |
12.32 |
12.02 |
Таблица 6. Среднее количество склеритов в каждом годовом кольце малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, 2011-12 гг.
|
Возраст, годы |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
– |
13.9 ± 0.2 |
13.8 ± 0.3 |
13.0 ± 0.3 |
13.0 ± 0.4 |
12.4 ± 0.5 |
12.2 ± 1.0 |
12.9 ± 2.0 |
Примечание. Как к табл. 4.
Таблица 7. Средние значения расчисленной длины ( АС ), мм, удельной скорости роста ( Cl ) и константы роста по Шмальгаузену ( C lt ) малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, 2011-12 гг.
|
Возраст, годы |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
n |
|||||
|
Cамцы |
|||||||||||||
|
Среднее |
153.7 |
187.6 |
220.1 |
249.0 |
281.2 |
308.6 |
– |
||||||
|
Прирост |
32.9 |
32.5 |
28.9 |
32.2 |
27.4 |
– |
106 |
||||||
|
C l |
0.20 |
0.16 |
0.12 |
0.12 |
0.09 |
– |
|||||||
|
C lt |
0.30 |
0.40 |
0.43 |
0.55 |
0.51 |
– |
|||||||
|
Самки |
|||||||||||||
|
Среднее |
154.5 |
189.9 |
224.4 |
256.9 |
286.5 |
307.4 |
325.1 |
||||||
|
Прирост |
35.4 |
34.5 |
32.5 |
29.6 |
20.9 |
17.7 |
133 |
||||||
|
C l |
0.21 |
0.17 |
0.14 |
0.11 |
0.07 |
0.06 |
|||||||
|
C lt |
0.31 |
0.42 |
0.47 |
0.49 |
0.39 |
0.36 |
|||||||
4. Заключение
При исследовании морфологических особенностей чешуи малотычинкового сига Йокостровской Имандры было выявлено, что различные сектора данной минерализованной структуры в течение жизни относительно высоты и длины тела растут неодинаково: передний и боковой сектора чешуи увеличиваются с возрастом, задний сектор – уменьшается. При этом форма чешуи остается постоянной. Подсчет склеритов на чешуе правильнее вести по переднему (базальному) радиусу, так как здесь их максимальное число. Размеры годовых колец рекомендуется снимать с переднего диагонального радиуса чешуи.
По нашим данным, зависимость между размерами тела и чешуи малотычинкового сига описывается уравнением линейной регрессии. Регрессионный метод обратных расчислений длины ( АС ) может быть рекомендован как оптимальный при исследованиях линейного темпа роста сига Йокостровской Имандры.
Вариабельность в длине тела сигов увеличивается с возрастом и достигает максимальных значений на 5-й год жизни, что, возможно, приводит к неодновременному наступлению созревания сигов. Компенсаторный рост наблюдается на 6 и 7-й годы жизни – после массового созревания самцов и самок.
Самые высокие значения скорости роста характерны для первого года жизни сигов, что также характерно и для количества сформированных склеритов в этот год. Период снижения темпа роста рыб наступает после массового их нереста. Для впервые нерестящихся сигов за исследуемый период более высокие темпы роста приводят к более раннему возрасту вступления в нерестовое стадо.
Таблица 2. Изменение величины переднего, переднего диагонального, бокового и заднего радиусов чешуи относительно максимальной высоты ( qh ) и длины ( АС ) тела, % с возрастом малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, 2011-12 гг.
|
– |
1+ |
2+ |
3+ |
4+ |
5+ |
6+ |
7+ |
|
относительно qh |
|||||||
|
Передний |
3.15 ± 0.24 (12) |
3.40 ± 0.25 (9) |
3.77 ± 0.12 (51) |
4.02 ± 0.11 (39) |
3.94 ± 0.10 (49) |
4.12 ± 0.18 (9) |
4.39 ± 0.53 (4) |
|
Передний диагональный |
3.92 ± 0.22 (12) |
4.02 ± 0.22 (9) |
4.26 ± 0.11 (51) |
4.43 ± 0.10 (39) |
4.35 ± 0.09 (49) |
4.51 ± 0.18 (9) |
4.87 ± 0.39 (4) |
|
Боковой |
3.76 ± 0.20 (12) |
3.86 ± 0.18 (9) |
3.87 ± 0.09 (51) |
3.89 ± 0.15 (39) |
3.77 ± 0.09 (49) |
3.66 ± 0.15 (9) |
3.68 ± 0.25 (4) |
|
Задний |
4.04 ± 0.13 (12) |
3.73 ± 0.13 (9) |
3.51 ± 0.08 (51) |
3.50 ± 0.08 (39) |
3.48 ± 0.07 (49) |
3.03 ± 0.08 (9) |
2.94 ± 0.36 (4) |
|
относительно АС |
|||||||
|
Передний |
0.60 ± 0.04 (12) |
0.70 ± 0.05 (9) |
0.83 ± 0.02 (51) |
0.90 ± 0.02 (39) |
0.89 ± 0.02 (49) |
1.04 ± 0.03 (9) |
1.08 ± 0.13 (4) |
|
Передний диагональный |
0.75 ± 0.03 (12) |
0.81 ± 0.05 (9) |
0.93 ± 0.02 (51) |
0.99 ± 0.02 (39) |
0.98 ± 0.02 (49) |
1.13 ± 0.03 (9) |
1.18 ± 0.08 (4) |
|
Боковой |
0.72 ± 0.03 (12) |
0.77 ± 0.03 (9) |
0.83 ± 0.01 (51) |
0.86 ± 0.03 (39) |
0.86 ± 0.02 (49) |
0.92 ± 0.02 (9) |
0.89 ± 0.02 (4) |
|
Задний |
0.78 ± 0.02 (12) |
0.76 ± 0.03 (9) |
0.77 ± 0.01 (51) |
0.75 ± 0.02 (39) |
0.79 ± 0.01 (49) |
0.77 ± 0.02 (9) |
0.71 ± 0.04 (4) |
Примечание. Как к табл. 1.
Таблица 4. Результаты обратного расчисления длины ( АС ) по методу регрессии, значения удельной скорости роста ( C l ) малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра, 2011-12 гг.
|
Возраст, лет |
l 1 |
l 2 |
l 3 |
l 4 |
l 5 |
l 6 |
l 7 |
n |
|
1+ |
154.6 ± 2.2 |
– |
– |
– |
– |
– |
– |
12 |
|
2+ |
149.8 ± 1.9 |
180.0 ± 3.0 |
– |
– |
– |
– |
– |
35 |
|
3+ |
158.3 ± 1.3 |
196.4 ± 2.2 |
230.7 ± 3.2 |
– |
– |
– |
– |
69 |
|
4+ |
151.8 ± 1.2 |
186.9 ± 2.2 |
219.3 ± 3.1 |
254.6 ± 3.7 |
– |
– |
– |
58 |
|
5+ |
155.0 ± 1.1 |
187.4 ± 2.3 |
220.4 ± 3.7 |
256.6 ± 4.6 |
289.0 ± 5.8 |
– |
– |
51 |
|
6+ |
155.3 ± 1.7 |
181.7 ± 3.6 |
211.3 ± 4.7 |
254.6 ± 8.9 |
289.8 ± 11.0 |
319.5 ± 11.8 |
– |
11 |
|
7+ |
147.3 ± 1.2 |
171.7 ± 1.9 |
198.2 ± 5.0 |
223.3 ± 7.0 |
252.7 ± 7.5 |
289.1 ± 11.0 |
325.1 ± 15.5 |
7 |
|
Среднее |
154.0 ± 0.6 |
188.1 ± 1.2 |
222.4 ± 1.8 |
253.7 ± 2.7 |
285.5 ± 4.8 |
307.7 ± 8.9 |
325.1 ± 15.5 |
|
|
Прирост |
34.1 34.3 31.3 31.8 22.2 17.4 |
243 |
||||||
|
C l |
0.20 0.17 0.13 0.12 0.07 0.06 |
|||||||
Примечание. В таблицах представлено среднее значение и его ошибка.
Таблица 8. Сравнение темпа линейного роста нерестящихся (III, IV стадия зрелости) и не нерестящихся (II, II-III стадия зрелости) особей малотычинкового сига C. lavaretus lavaretus плеса Йокостровская Имандра в разном возрасте, 2011-12 гг.
|
– |
Стадия зрелости |
Расчисленная длина рыб ( АС ), мм |
Наблюдаемая длина ( АС ), мм |
n |
|||||
|
l 1 |
l 2 |
l 3 |
l 4 |
l 5 |
l 6 |
||||
|
Возраст |
3+ |
||||||||
|
II |
155.8 ± 1.6 |
189.8 ± 2.5 |
222.4 ± 3.7 |
8.2 ± 0.7 |
– |
– |
242.8 ± 5.4 |
34 |
|
|
Самцы |
III, IV |
169.4 ± 3.1 |
224.4 ± 1.7 |
273.0 ± 6.4 |
11.2 ± 0.9 |
– |
– |
301.4 ± 5.4 |
5 |
|
t |
3.09 |
5. 1 9 |
4.99 |
1.57 |
– |
– |
4.87 |
– |
|
|
Возраст |
4+ |
||||||||
|
II |
149.4 ± 2.3 |
178.5 ± 3.2 |
208.8 ± 4.9 |
242.6 ± 6.7 |
9.4 ± 0.9 |
– |
265.4 ± 7.3 |
21 |
|
|
Самцы |
III, IV |
155.2 ± 2.8 |
194.4 ± 4.3 |
230.3 ± 6.8 |
268.8 ± 5.6 |
8.4 ± 1.3 |
– |
284.3 ± 8.5 |
6 |
|
t |
1.27 |
2.48 |
2.15 |
2.01 |
0.50 |
– |
1.15 |
– |
|
|
II |
151.9 ± 1.6 |
190.2 ± 2.9 |
224.2 ± 4.2 |
260.5 ± 4.7 |
8.1 ± 0.8 |
– |
278.2 ± 5.1 |
29 |
|
|
Самки |
III, IV |
172.7 |
227.8 |
252.3 |
276.8 |
6.0 |
– |
289.0 |
1 |
|
t |
– |
– |
– |
– |
– |
– |
– |
– |
|
|
Возраст |
5+ |
||||||||
|
II |
154.1 ± 1.4 |
186.3 ± 3.8 |
220.2 ± 4.5 |
256.8 ± 6.5 |
287.5 ± 7.6 |
7.0 ± 0.9 |
304.4 ± 8.9 |
21 |
|
|
Самки |
III, IV |
159.4 ± 1.9 |
197.1 ± 6.2 |
233.8 ± 12.3 |
272.1 ± 12.2 |
305.8 ± 16.3 |
6.4 ± 1.1 |
321.6 ± 16.5 |
10 |
|
t |
2.20 |
1.56 |
1.28 |
1.22 |
1.16 |
0.40 |
1.00 |
– |
|
Примечание. В таблице представлено среднее значение и его ошибка, жирным выделено среднее количество склеритов, которое образовалось в текущем году, t – коэффициент Стьюдента, линиями подчеркнуты достоверные различия (одной – P > 0.5, двумя – P > 0.9, пунктиром – P > 0.99).
