Морфология порово-канальной системы в альбидной ткани некоторых позднедевонских конодонтовых элементов

Автор: Журавлев А.В.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 1 (277), 2018 года.
Бесплатный доступ
Комплексом методов (световая и электронная микроскопия, рентгеновская микротомография) изучено распределение и морфология пор и «каналов» в альбидной ткани конодонтовых элементов, принадлежащих к позднедевонско-раннекаменноугольным представителям отрядов Ozarkodinida Dzik 1976 ( Mehlina Youngquist, Youngquistognathus Myshkina et Zhuravlev, Polygnathus Hinde, Bispathodus Müller), Prioniodontida Dzik 1976 ( Jablonnodus Dzik) и Priniodinida Sweet 1988 ( Ligonodina Bassler). Морфология порово-канальной системы конодонтовых элементов демонстрирует различия на уровне таксонов родового ранга, что позволяет предположить возможность использования этого признака в систематике конодонтов. Отмеченное для рода Youngquistognathus единство в строении порово-канальной системы в различных элементах аппарата дает основания предполагать, что данный признак перспективен при реконструкции аппаратов по разрозненным элементам.
Конодонты, гистология, поздний девон
Короткий адрес: https://sciup.org/149129271
IDR: 149129271 | УДК: 56.016.3 | DOI: 10.19110/2221-1381-2018-1-19-24
Morphology of the pores-and-canals system in the albid tissue of some Late Devonian conodont elements
Distribution and morphology of pores and “canals” in the albid tissue of conodont elements of orders Ozarkodinida Dzik 1976 ( Mehlina Youngquist, Youngquistognathus Myshkina et Zhuravlev, Polygnathus Hinde, Bispathodus Müller), Prioniodontida Dzik 1976 ( Jablonnodus Dzik), and Priniodinida Sweet 1988 ( Ligonodina Bassler) are studied by light microscopy, SEM, and X-ray micro-CT. Taxonomical differences (at the generic level) of morphology of the pore-and-canal system of conodont elements promise systematical application of these features. Similarity in the composition of the pore-and-canal system in elements of the apparatus of Youngquistognathus suggests a chance to use the composition in reconstructing apparatuses on the basis of disjuncted conodont elements.
Текст научной статьи Морфология порово-канальной системы в альбидной ткани некоторых позднедевонских конодонтовых элементов
Конодонтовые элементы представляют собой сильно минерализованные части ротового аппарата конодонтов — полностью вымершей в конце триаса группы морских организмов. Все конодонтовые элементы состоят из двух частей, отличающихся по степени минерализации, — базального заполнения (basal callus) с низким содержанием минеральной компоненты и сильно минерализованной кроны (crown, более 96 % фторгидроксилапатита). Крона конодонтовых элементов образована четырьмя типами твердых тканей: ламеллярной, интерламеллярной, парала-меллярной и альбидной [1]. Альбидная ткань, наличие которой рассматривается в качестве аутапоморфно-го признака конодонтов [5], характеризуется высокой степенью кристалличности минеральной компоненты, представляющей собой пористый мезокристалл [7]. Одним из специфических признаков альбидной ткани наряду с кристаллическим строением является присутствие в ней многочисленных и относительно крупных полостей различной морфологии [10,4, 6,2], которые обуславливают ее непрозрачность в проходящем свете и белый цвет в отраженном. Несмотря на то что этот признак известен уже давно, еще с классической работы X. Пандера [10], особенности морфологии полостей остаются слабоизученными. В частности, существуют противоречивые данные об их размере и форме [4, 6], что обусловлено трудностью изучения этих аспектов в шлифах и пришлифовках.
Данная работа посвящена морфологической характеристике «порового пространства» альбидной тка ни и предварительной оценке таксономического значения этого признака. При этом под «порами» понимаются относительно изометричные изолированные полости, а под «каналами» — цепочки обычно удлиненных и единообразно ориентированных, часто изолированных друг от друга полостей.
Материал
Для данного исследования были использованы позднедевонские конодонтовые элементы различной морфологии (конические, рамиформные, листовидные и платформенные) и систематической принадлежности (отряд Ozarkodinida Dzik 1976, роды Mehlina Youngquist, Youngquistognathus Myshkina et Zhuravlev, Polygnathus Hinde; отряд Prioniodontida Dzik 1976, род Jablonnodus Dzik; отряд Priniodinida Sweet 1988 род Ligonodina Bassler) из местонахождений Главного и Центрального девонского поля (ВосточноЕвропейская платформа). Коллекция конодонтовых элементов родов Mehlina , Youngquistognathus , Polygnathus, Ligonodina происходит из среднефран-ских (конодонтовая зона Pal. punctata) ильменских и бурегских слоев в разрезах на южном берегу оз. Ильмень [14], а также раннефранских отложений на р. Сясь (Главное девонское поле). Раннефаменские Jablonnodus изучены из задонского горизонта в верхней части разреза Русский Брод (Центральное девонское поле) [12]. В качестве сравнительного материала были использованы оистодиформные (конические) раннеордовикские конодонтовые элемен- 19
ты из разреза на р. Поповка (Ленинградская область, г. Павловск), а также раннекаменноугольные (турней-ские) конодонтовые элементы Ligonodina sp. из разреза на р. Каменка (Печорская гряда) [13] и позднефа-менские конодонтовые элементы Bispathodus bispathodus Ziegler, Sandberg, Austin из разреза на р. Талота (северо-восток Тимано-Печорской провинции) [3].
Все гистологически изученные конодонтовые элементы характеризуются хорошей сохранностью (индекс окраски 1—2), отсутствием следов вторичных изменений, что практически исключает возможность посмертного формирования пор и каналов. Выбор указанных родов обусловлен их хорошей гистологической изученностью [1, 8] и представительностью в использованном в данном исследовании коллекционном материале.
Методы
Конодонтовые элементы изучались в иммерсионных препаратах, толстых шлифах и пришлифовках с использованием оптической микроскопии, а также в полированных пришлифовках с применением электронной сканирующей микроскопии. Кроме того, ряд элементов был изучен с помощью рентгеновской микротомографии с пространственным разрешением 1—2 мкм на рентгеновском микротомографе SkyScan 1272 (НМСУ «Горный», оператор А. А. Штырляева). Морфометрические исследования по томограммам осуществлялись с помощью программного обеспечения CTAn (Brucker), Voxler (Golden Software) и PAST [9]. Органическое вещество в альбидной ткани конодонтовых элементов изучалось с помощью трихром-ного окрашивания по Ван Гизону [2, 8].
Детальная информация о морфологии пор и «каналов» в альбидной ткани получена по томограммам, а уже на ее основе интерпретировались результаты изучения иммерсионных препаратов, шлифов и при-шлифовок.
Результаты
Массивы альбидной ткани слагают исключительно ядра зубцов срединных гребней и их гомологов в первичных отростках конодонтовых элементов [1]. У конических элементов альбидная ткань формирует ядро зубца. В других морфологических образованиях (платформы различного происхождения и формы, вторичные гребни, дополнительные зубцы) альбидная ткань отсутствует. Исключение представляет распределение альбидной ткани в эхелонах некоторых спа-тогнатид ( Skeletognathus , некоторые Mehlina ). Однако это исключение кажущееся, так как эхелоны гомологичны срединному гребню.
Величина пористости альбидной ткани, определенная рентгенотомографическим методом (учтены поры крупнее 2 мкм), достигает 3—4 %. Распределение пор и каналов в массивах альбидной ткани в изученных конодонтовых элементах различно.
В конических элементах рода Jablonnodus альбидная ткань образует большую часть элемента. Крупные поры в массиве альбидной ткани формируют продольную центральную цепочку («канал»), окруженную более мелкими порами (рис. 1). При этом сред- 20

Рис. 1. Центральный «канал» в альбидной ткани конического элемента рода Jablonnodus Dzik; нижний фамен, разрез Русский Брод, Центральное девонское поле: A — рентгенотомографическое изображение элемента; B — виртуальный срез элемента; поры канала отмечены стрелками
Fig. 1. Central «canal» in the albid tissue of the coniform element of the genus Jablonnodus Dzik; the Lower Famennian, Russkij Brod section, Central Devonian Field: A — mirco-CT image of the element; B — Virtual section of the element; pores composing the canal are marked by arrows ний объем пор составляет 2 мкм3. Мелкие поры не демонстрируют отчетливой упорядоченности.
Раннеордовикские конические оистодиформные элементы (“ Oistodus ” s. f.), альбидная ткань которых обладает менее упорядоченным распределением кристаллитов [8], несут хаотически расположенные относительно крупные (в среднем 6 мкм3) поры и не имеют «каналов».
В элементах рода Youngquistognathus ядра альбидной ткани, образующие зубцы срединного гребня, характеризуются наличием крупных, часто удлинённых пор. Поры формируют конические структуры, ориентированные вершиной конуса внутрь элемента, иногда с центральным «каналом» (цепочкой пор) (рис. 2). Наиболее крупные поры располагаются на границе альбидной и параламеллярной тканей (рис. 2). Подобное распределение пор и каналов установлено в Pa-, Pb-, Sb- и M-элементах аппарата Youngquistognathus (рис. 2). Средний объем пор в Sb-элементе составляет 3.2 мкм3, в Pa-элементах 2.5 мкм3. Сходная морфология пор и каналов наблюдается у представителей предкового по отношению к Youngquistognathus рода Mehlina и у переходных форм — Y. praeangustidiscus Zhuravlev (рис. 3). В зубцах осевого гребня Pa-элементов Mehlina gradata Youngquist средний диаметр пор составляет около 1 мкм (вариации от 0.6 до 1.5 мкм), а сами поры упорядочены в конические структуры. В осевом гребне Pb-элементов этого вида поры крупнее, в среднем их диаметр около 1.7 мкм (от 0.9 до 3.5 мкм). Поры в альбидной ткани близкого по морфологии к Mehlina фаменско-турней-ского рода Bispathodus распределены более равномерно и не образуют конических структур (рис. 3).
В изученных Pa-элементах рода Polygnathus ( P. la-nei Kuzmin, P. ljaschenkoi Kuzmin, P. webbi Stauffer) поры в альбидной ткани мелкие, близкие к шарообраз-
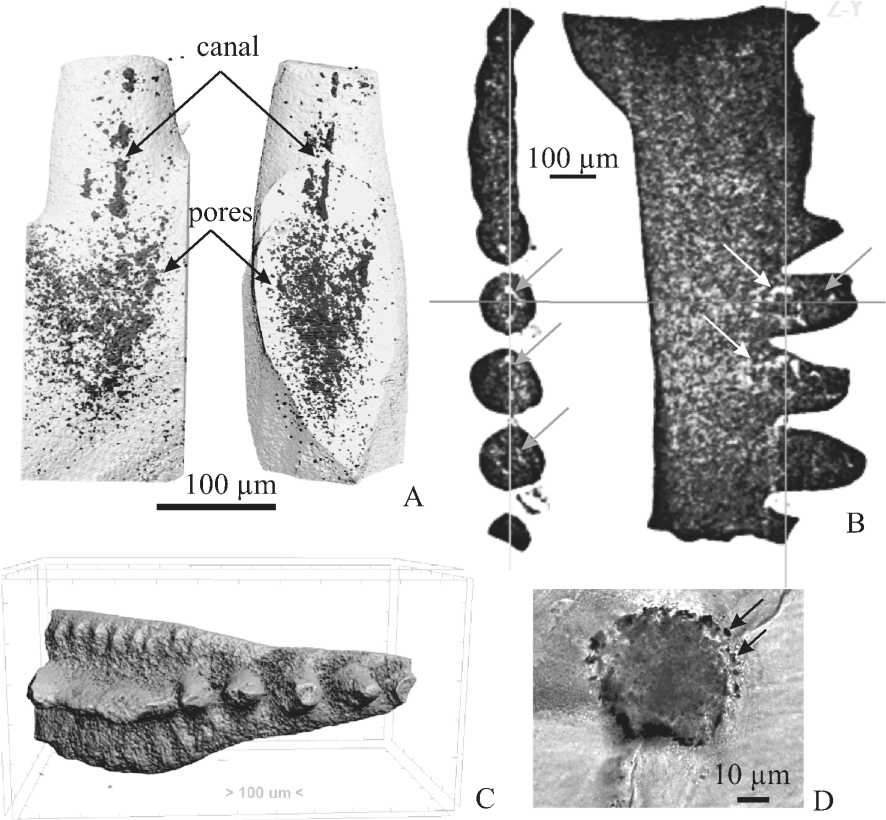
Рис. 2. Распределение пор и «каналов» в альбидной ткани в элементах Youngquistognathus rossicus (Zhuravlev), средний фран, разрез на оз. Ильмень, Главное девонское поле: A — поры и центральный «канал» в зубце Sb-элемента, рентгенотомографическое изображение; B — ортогональные сечения томограммы Pa-элемента; C — томографическое изображение Pa-элемента; D — поперечный шлиф зубца Sa-элемента. Поры и «каналы» отмечены стрелками
Fig. 2. Distribution of the pores and «canals» in the albid tissue of elements of Youngquistognathus rossicus (Zhuravlev), the Middle Frasnian, Il’men Lake section, Main Devonian Field: A — Pores and central «canal» in the denticle of Sb element; B — Orthogonal sections of the tomogram of Pa element; C — microCT image of Pa element; D — Transverse thin section of the denticle of Sa element. Pores and «canals» are marked by arrows
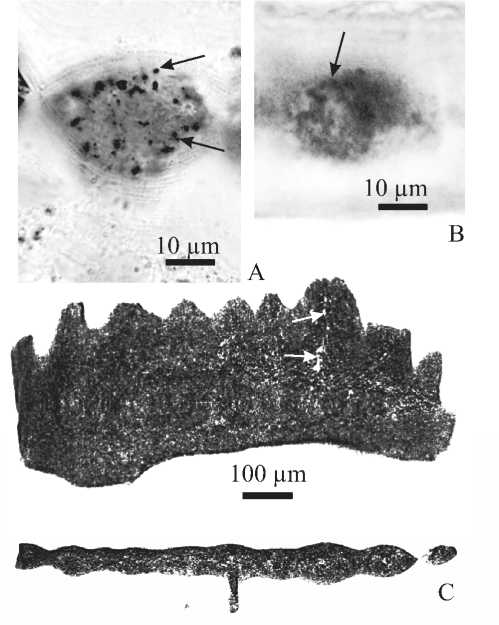
Рис. 3. Распределение пор в альбидной ткани зубцов осевого гребня листовидных Pa-элементов: A — поперечный шлиф зубца Mehlina gradata (Youngquist), средний фран, разрез на оз. Ильмень, Главное девонское поле; B — поперечный шлиф зубца Bispathodus bispathodus Ziegler, Sandberg, Austin, верхний фамен, разрез на р. Талота, северо-восток Тимано-Печорской провинции; C — ортогональные сечения томограммы Youngquistognathus praeangustidiscus Zhuravlev, нижний фран, разрез на р. Сясь, Главное девонское поле. Поры отмечены стрелками
Fig. 3. Distribution of the pores in the albid tissue of the carina denticles of the blade-like Pa elements. A — Transverse thin section of the denticle of Mehlina gradata (Youngquist), the Middle Frasnian, Il’men Lake section, Main Devonian Field; B — Transverse thin section of the denticle of Bispathodus bispatho-dus Ziegler, Sandberg, Austin, Upper Famennian, the Talota River section, NE Timan-Pechora Province; C — Orthogonal sections of the tomogram of Youngquistognathus praeangustidis-cus Zhuravlev, the Lower Frasnian, Syas’ River section, Main Devonian Field. Pores are marked by arrows ным, не формируют центральных каналов и образуют конические структуры с вершиной конуса, направленной к вершине зубца (рис. 4). В зубцах осевого гребня Pa-элемента Polygnathus lanei Kuzmin диаметр пор составляет 0.2—0.3 мкм, в Pa-элементах P. ljaschenkoi Kuzmin — 0.5—1.0 мкм (средний 0.7 мкм), а в Pa-элементах P. webbi Stauffer — 0.8—1.0 мкм. При этом объем пор редко превышает 0.5 мкм3.
В Sa-элементе турнейского представителя рода Ligonodina (отряд Prioniodinida) поры формируют цепочку («канал») в средней и верхней частях главного зубца (рис. 5, A). Средний объем пор составляет
10 цт
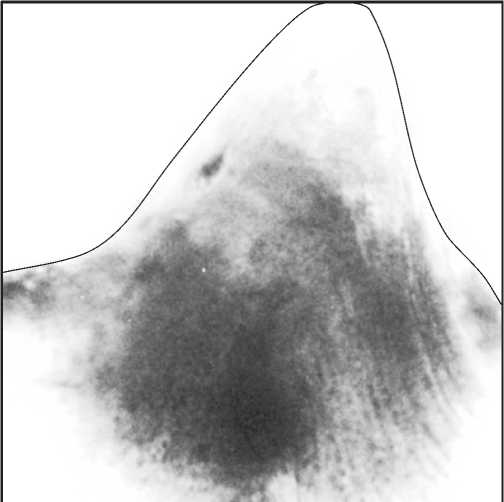
Рис. 4. Распределение пор в альбидной ткани зубца осевого гребня Pa-элемента Polygnathus lanei Kuzmin, средний фран, разрез на оз. Ильмень, Главное девонское поле
Fig. 4. Distribution of pores in the albid tissue of carina denticles of the Pa-element of Polygnathus lanei Kuzmin, the Middle Frasnian, Il’men Lake section, Main Devonian Field 21
100 мкм3, а их диаметр около 4.4 мкм. При этом в количественном отношении преобладают мелкие поры (около 0.8 мкм). Аналогичное строение демонстрируют и Sc-элементы среднефранских представителей этого рода. Размер пор, составляющих «канал», достигает у них 13 мкм (в среднем 5 мкм) в верхней части зубцов и 2 мкм в нижней (рис. 5, B). Поры вне «канала» имеют диаметр от 0.3 до 2.5 мкм, и их размер также увеличивается вверх внутри зубца.
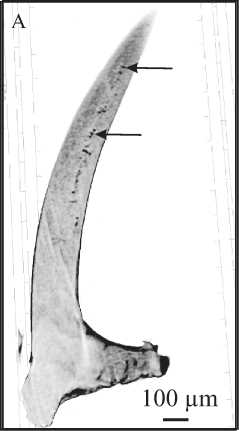
Рис. 5. Центральный «канал» и поры в альбидной ткани S-элементов Ligonodina: A — рентгенотомографический виртуальный срез Sa-элемента; поры канала отмечены стрелками; верхнее турне, разрез на р. Каменка, южная часть Печорской гряды; B — СЭМ-изображение поперечного сечения нижней части зубца Sc-элемента; средний фран, разрез на оз. Ильмень, Главное девонское поле
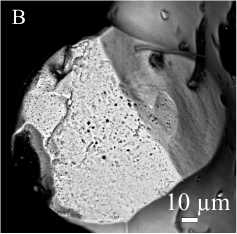
Fig. 5. Central canal and pores in the albid tissue of the S elements of Ligonodina: A— mirco-CT virtual section of Sa element; pores composing the canal are marked by arrows; the upper Tournaisian, Kamenka River section, the southern part of the Pechora Swell; B — SEM image of the cross-section of the lower part of the denticle of SC element; the middle Frasnian, Il’men Lake section, Main Devonian Field.
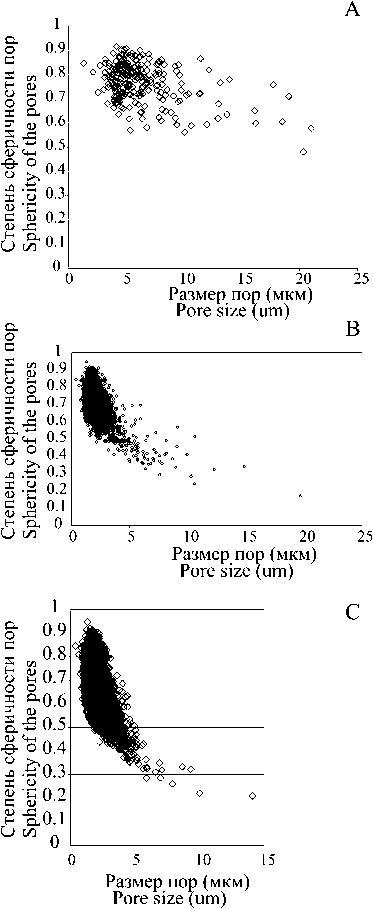
Рис. 6. Зависимость степени сферичности от размера пор в альбидной ткани на основе данных микротомографии: A — Pa-элемент Youngquistognathus rossicus (Zhuravlev); B — Sb-элемент Youngquistognathus sp.; C — конический элемент Jablonnodus sp.
У всех изученных конодонтовых элементов наблюдается сходная зависимость формы (степени сферичности) пустот в альбидной ткани от объема этих пустот. Как видно из графиков (рис. 6), чем крупнее пора, тем дальше ее форма от сферы. Крупные поры, как правило, сильно удлиненные, и именно они формируют центральные «каналы» в зубцах.
Обсуждение результатов
Морфология порово-канальной системы в зубцах конодонтовых элементов демонстрирует различия на уровне таксонов родового ранга. При этом для рода Youngquistognathus отмечается сходство в строении и размерах порово-канальной системы в пределах аппарата: S-, M- и P-элементы обладают крупными порами и несут центральные «каналы» в зубцах осевого гребня (рис. 2). Также наблюдается сходство поровоканальной системы в зубцах Pa-элементов родствен- 22
Fig. 6. Sphericity and size of pores in the albid tissue based on micro-CT. A — Pa element of Youngquistognathus rossicus (Zhuravlev); B — Sb element of Youngquistognathus sp.; C — coniform element of Jablonnodus sp.
ных родов Mehlina и Youngquistognathus (рис. 3, A, C). У сходных с Youngquistognathus по общей морфологии Pa-элементов представителей рода Polygnathus поровоканальная система существенно отличается и по размеру пор, и по их распределению (рис. 4). Аналогичная ситуация наблюдается при сравнении морфологически близких родов Mehlina и Bispathodus . Представители этих родов демонстрируют различия в строении порово-канальной системы: у Bispathodus менее упорядочено распределение пор, которые не образуют конических структур (рис. 3, A, B).
Следует, однако, отметить, что у всех изученных конодонтовых элементов «каналы» формируются на поздних стадиях онтогенеза (присутствуют толь- ко в средней и верхней частях зубцов). Поэтому данный признак не может использоваться для диагностики элементов на ранних стадиях онтогенеза.
Все поры, даже те, которые формируют «каналы», изолированы друг от друга. Таким образом, их образование могло происходить исключительно на этапе роста соответствующей части конодонтового элемента. В дальнейшем эти поры, в силу своей полной изолированности, не могли содержать живые клетки, что подтверждается отсутствием следов клеточного вещества в деминерализованных конодонтовых элементах [2]. По данным спектроскопии комбинационного рассеивания и трихромного окрашивания, поры и каналы в альбидной ткани содержат органическое вещество, близкое по составу к органической матрице конодонтового элемента [2].
Наиболее вероятно, что полости и каналы в альбидной ткани образовывались на месте нерезорбиро-ванной части органической (предположительно коллагеновой) матрицы, вокруг которой формировалась минеральная компонента ткани. Размер пор в целом недостаточен для расположения в них эукариот-ных клеток, однако он вполне подходит как вместилище белковых структур. Образование пор и каналов, скорее всего, происходило на этапе преобразования ткани «внешней каймы» с низкой упорядоченностью кристаллитов и высокой нанопористостью [2] в аль-бидную ткань с высокой степенью упорядоченности и плотной упаковкой кристаллитов [11, 2].
Выводы
Различия в строении порово-канальной системы морфологически сходных конодонтовых элементов различных родов ( Oistodus s. f и Jablonnodus , Youngquistognathus и Polygnathus ; Bispathodus и Mehlina ) позволяют предположить возможность использования этого признака как для уточнения систематической (родовой) принадлежности конкретных видов, так и для таксономической диагностики (на родовом уровне) фрагментов конодонтовых элементов. Общность в строении порово-канальной системы у элементов в пределах аппарата, отмеченная для представителей рода Youngquistognathus , дает основания предполагать, что данный признак перспективен при реконструкциях аппаратов по разрозненным элементам.
Автор выражает благодарность С. С. Шевчуку, А. С. Шуйскому и В. Н. Филиппову за помощь в электронно-микроскопических исследованиях в ЦКП «Геонаука», А. А. Штырляевой за содействие при рентгено-томографическом изучении конодонтовых элементов. Исследования выполнены при финансовой поддержке Программы УрО РАН 18-5-5-50.
Список литературы Морфология порово-канальной системы в альбидной ткани некоторых позднедевонских конодонтовых элементов
- Журавлев А. В. Гистология и микроскульптура позднепалеозойских конодонтовых элементов. СПб.: Геосервис-Плюс, 2002. 94 с.
- Журавлев А. В., Герасимова А. И. Альбидная ткань конодонтовых элементов: строение и модель формирования//Вестник Института геологии. 2015. № 10. С. 21-27.
- Разрез верхнего девона - нижнего карбона на р. Талота (Тимано-Печорская провинция) - результаты литологических, биостратиграфических и палеомагнитных исследований/Н. М. Еременко, А. В. Журавлев, А. Г. Иосифиди, В. В. Попов, А. Н. Храмов, Я. А. Вевель, А. И. Николаев //Нефтегазовая геология. Теория и практика. 2009. 4(4). URL: http://www.ngtp.ru/rub/2/45_2009.pdf
- Barnes C. R, Sass D. B., Monroe E. A. Ultrastructure of some Ordovician conodonts//The Geological Society of America Special Paper. 1973. 141. P. 1-30.
- Donoghue P. C. J. & Chauffe K. M. Conchodontus, Mitrellataxis and Fungulodus: Conodonts, fishes or both?//Lethaia. 1998. 31. P. 283-292.
