Морфометрическая характеристика изменений в ретроспленальной коре мышей при моделировании социального стресса

Автор: Дорофеев Н.А., Смирнов А.В., Самаркин Д.Д., Тюренков И.Н., Быхалов Л.С.
Журнал: Волгоградский научно-медицинский журнал @bulletin-volgmed
Рубрика: Статьи
Статья в выпуске: 1 т.22, 2025 года.
Бесплатный доступ
Исследования морфологического субстрата воздействия стресса на организм помогают понять роль различных структур головного мозга в реакциях на стрессоры, однако детальные механизмы этих реакций остаются недостаточно изученными.
Социальное поражение, хронический стресс, мыши, ретроспленальная кора, морфометрия
Короткий адрес: https://sciup.org/142245448
IDR: 142245448 | УДК: 616.8-091 | DOI: 10.19163/2658-4514-2025-22-1-57-65
Morphometric characteristics of changes in the retrosplenial cortex of mice during social stress modeling
Studies of the morphological substrate of stress effects on the organism help to understand the role of various brain structures in reactions to stressors, but the detailed mechanisms of these reactions remain poorly understood. Purpse. To study morphofunctional changes in the retrosplenial cortex, one of the important structures in the brain's reactions to stress, when modeling social stress in mice in comparison with intact animals.
Текст научной статьи Морфометрическая характеристика изменений в ретроспленальной коре мышей при моделировании социального стресса
doi:
Согласно современным представлениям хронический умеренный стресс могут вызывать различные причины. У человека особое значение имеют социальные факторы, связанные как с конфликтами в повседневной жизни, так и с глобальными событиями. Эти негативные воздействия, затрагивающие широкие слои населения, могут привести к дезадаптации и развитию заболеваний психической сферы с развернутой симптоматикой, в том числе депрессии и посттравматического стрессового расстройства [1–4].
На текущий момент существует много методов лекарственной терапии как указанных заболеваний, так и более мягких психических расстройств и тревожных состояний. Однако детальные механизмы их действия во многом остаются недостаточно изученными [4]. Особенно данный факт относится к исследованиям морфологического субстрата воздействия хронического стресса на организм, что возможно только в рамках эксперимента с участием животных, преимущественно крыс и мышей. С помощью различных экспериментальных моделей депрессии изучается роль отдельных структур головного мозга в реакции организма на патологические воздействия различной природы, в том числе физической, химической, социальной. Среди наиболее значимых и изученных в контексте развития депрессивно- / тревожно-подобных состояний выделяют гиппокамп и медиальную префронтальную кору, в меньшей степени – ядра миндалевидного тела и иные структуры конечного мозга, например, переднюю поясную кору, зрительную кору, таламус, скорлупу, латеральную перегородку [1–11]. В последнее время накапливаются данные относительно участия в реакциях на подобные стрессоры ретроспленальной коры (РСК), описанную как у грызунов, так и у приматов, в том числе у человека [3, 9, 10, 12].
Структурно в РСК выделяют гранулярную и агранулярную (дисгранулярную) части, в которых выражены или, наоборот, не выражены II и IV зернистые (гранулярные) слои [9]. В головном мозге мыши гранулярная часть располагается на медиальной поверхности в межполушарной борозде, начинаясь вентрально от мозолистого тела и переходя дорсально в агранулярную часть. Последняя располагается на границе медиальной и верхнелатеральной поверхностей с продолжением латерально во вторичную моторную кору.
Топографическая близость к зрительным областям и структурам лимбической системы, в частности к гиппокампу, обеспечивают значимую роль РСК в пространственной ориентации, процессах запоминания, воображении и связанных когнитивных функциях [3, 8–10, 12]. В контексте реакций на стресс и развития депрессивно- / тревожно-подобных состояний РСК отводится роль в запоминании условий, в которых происходило патологическое воздействие, и информации о предшествовавших ему стимулах при моделировании контекстно-обусловленного страха [9, 10, 12]. При этом накапливаются данные, что разные ее области могут иметь различные и даже противоположные функции при моделировании указанных процессов [9, 12].
ЦЕЛЬ РАБОТЫ
Изучить качественные и количественные морфофункциональные изменения в РСК головного мозга мышей при моделировании хронического социального стресса поражения в сравнении с интактными животными.
МЕТОДИКА ИССЛЕДОВАНИЯ
Исследование выполнено на мышах ( n = 13) линии CBA (lac) возрастом 4 месяца массой 20– 25 граммов, а также животных линии C57BL/6 и нелинейных мышах. Работу с животными проводили в соответствии с Европейской конвенцией о защите позвоночных животных, используемых в экспериментальных и других научных целях (Страсбург, 1986). На проведение исследования получено разрешение локального этического комитета ФГБОУ ВО ВолгГМУ Минздрава России (№ 2022/160 от 3.11.2022).
Эксперимент производился в парадигме хронического социального стресса поражения [2, 6, 7, 13, 14]. Для этого использовались специальные клетки, разделенные пополам прозрачным барьером со сквозными отверстиями, которые позволяли мышам видеть, слышать и чувствовать запахи друг друга, но предотвращали физический контакт между ними.
Предварительно, перед началом моделирования депрессивного состояния, осуществлялся отбор животных «агрессоров» – победителей. Для этого в домашнюю клетку с животными (линии C57BL/6 и нелинейными) помещалось одно незнакомое животное из другой клетки и оценивался характер их взаимодействия. В группу «агрессоры» выбирались те мыши, которые активно защищали свою территорию, проявляли агрессивные действия в отношении «чужака». После выявления с целью обосноваться в новой среде животное «агрессор» на 3 суток перемещалось в свою часть клетки, разделенной барьером, где заранее были подготовлены опилки и материалы, необходимые для строительства домика и обогащения среды.
В группу «жертв» выбирались самцы линии CBA (lac) возрастом 4 месяца, каждый из которых спустя 3 суток перемещался в свою часть клетки, разделенной перегородкой. В последующем ежедневно, на протяжении 10 суток, побежденное животное перемещали в новую клетку к неизвестному ему ранее агрессору и убирали барьер, отделяющий их друг от друга, провоцируя, таким образом, агрессора защищать собственную территорию от новичка и вступать в физические конфронтации с ним. Если физический контакт (драка) между агрессором и жертвой продолжались непрерывно 3 минуты, то между ними обратно устанавливали преграду, в противном случае продолжительность столкновений длилась 10 минут. На 4–5-е сутки ежедневных столкнове- ний животные со статусом «жертвы» в большей части прекращали оказывать должное сопротивление на акты агрессии со стороны победителя, в результате чего продолжительность конфронтаций между мышами сокращали до 30 секунд, с целью предотвращения травмирования животных.
Далее начинался этап 10-дневного лечения, которое проводилось на фоне продолжения ежедневных социальных конфронтаций. В данной статье рассматриваются группы интактных животных (не подвергающихся воздействую стрессовых факторов) и стрессированные животные, которым во втором периоде вводился 0,9%-й раствор NaCl. Спустя 10 дней терапии стрессиро-вание прекращалось, животных выводили из эксперимента методом цервикальной дислокации с последующей декапитацией.
Для дальнейшего анализа использовались образцы головного мозга 6 мышей интактной группы и 7 – группы стресса. Морфологическое исследование проводилось на фронтальных срезах на уровне -1,255 : -2,555 мм от Брегмы, из которых изготавливались микропрепараты, окрашенные тионином по методу Ниссля по стандартной процедуре. Далее проводилась фотофиксация с использованием микроскопа «Axio Lab. A1» (Carl Zeiss Microscopy GmbH, Germany), фото- / видеокамеры «AxioCam 105 color» (Carl Zeiss Microscopy GmbH, Germany). На полученных микрофотографиях выполнена морфометрия с подсчетом площади (мкм2) и периметра (мкм) ядер и перикарионов нейронов, площади (мкм2) цитоплазмы перикарионов на 30 клетках. Также были рассчитаны относительные площади ядер, перикарионов нейронов и цитоплазмы перикарионов по формуле « Относительный показатель = Площадь структуры / Площадь тест системы », где под тест-системой подразумевается анализируемая область интереса. Кроме того, представлен показатель ядерно-цитоплаз-матического отношения (ЯЦО), рассчитанный по формуле « ЯЦО = площадь ядра / площадь цитоплазмы перикариона ». Измерения выполнялись в программе Image-Pro Plus (Media Cybernetics, USA) каждой из следующих структур: II/III, V слоях гранулярной (РСГК) и агранулярной / дисграну-лярной (РСАК) частей РСК.
Статистическая обработка осуществлялась с использованием языка программирования R (версия 4.3.1) и среды разработки RStudio. Производился расчет параметров описательной статистики, включая среднее (M, Mean), стандартное отклонение (SD, Standard Deviation), медиану (Me, Median), 1-й и 3-й квартили (Q1, Q3).
Характер распределения указанных количественных непрерывных переменных оценивался с помощью приведенных показателей описательной статистики, теста Shapiro-Wilk (SW-test, тест на нормальность распределения), а также графической визуализации с построением «box-plot» («ящик с усами»).
На следующем этапе, в зависимости от характера распределения, проводился либо Welch Two Sample t-test (t-критерий Уэлча, модификация t-критерия Стьюдента, не предполагающий равенства дисперсий в сравниваемых выборках), либо его непараметрический аналог – Wilcoxon rank sum test (критерий суммы рангов Уилкоксона, син. тест Манна – Уитни) – с расчетом уровня значимости «p-value».
Для показателей и локализаций, в которых установлены статистически значимые различия, рассчитана разность средних между группой стресса и группой интакта с построением 95 % доверительного интервала (ДИ) методом классического бутсрэпа (bootstrap) с представлением в виде «Средняя разность (Нижняя граница 95 % ДИ – Верхняя граница 95 % ДИ)». Также указанные показатели разности представлены в формате процентного изменения относительно значений в интактной группе в виде «Средний % изменений [Нижняя граница 95 % ДИ (%) – Верхняя граница 95 % ДИ (%)]».
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В работах, посвященных исследованию участия РСК в реакции организма на стрессовые воздействия, наиболее часто исследовались II и V слои гранулярной части, а также II/III и V слои агранулярной части, так как в данной области отсутствует оформленный II слой.
При гистологическом исследовании препаратов головного мозга, окрашенных по Нисслю, в гранулярной части ретроспленальной коры не отмечено значительной вариабельности формы и расположения нейронов II/III и V слоев между животными разных групп (см. рис.). В группе стрессированных мышей нейроны данной зоны имели тенденцию к более компактному и упорядоченному расположению. В то же время заметные различия в морфологии слоев агранулярной части ретроспленальной коры не были обнаружены при визуальной качественной оценке микропрепаратов.
По данным морфометрии в каждой из описанных 4 зон (по 2 слоя нейронов в двух частях) ретроспленальной коры головного мозга получены результаты, представленные в табл. 1–4.
Согласно анализу характера распределений полученных данных в качестве метода сравнения t-критерий Уэлча был выбран для показателей «Периметр перикариона нейрона» в зонах РСГК V, РСАК II/III, РСАК V; «Площадь ядра нейрона» в зонах РСГК V, РСАК V; «ЯЦО нейрона» в зонах РСГК II/III, РСАК V. Для остальных комбинаций «Показатель-Зона ретроспленальной коры» сравнение производилось с помощью теста Манна – Уитни.
По результатам попарных сравнений и расчетов p-value по описанной выше методике достоверные различия между интактной и стрессированной группами установлены для следующих показателей:
-
- «Периметр перикариона нейрона» – зоны РСГК V [увеличение на 3,31 (1,76–4,85) мкм либо 9,5 % (5,1–14,0 %) в группе стресса], РСАК II/III [уменьшение на 2,4 (1,27–3,53) мкм либо 7,1 % (3,7–10,4 %) в группе стресса];
-
- «Периметр ядра нейрона» – зоны РСГК V [увеличение на 1,56 (0,65–2,46) мкм либо 7,3 % (3,0–11,5 %) в группе стресса], РСАК II/III [уменьшение на 1,66 (0,96–2,36) мкм, 7,8 % (4,5–11,1 %) в группе стресса];
-
- «Относительная площадь перикариона нейрона» – зона РСГК V [уменьшение на 0,17 (0,09– 0,25) либо 16,5 % (8,7–24,3 %) в группе стресса];
-
- «Площадь ядра нейрона» – зона РСАК II/III [уменьшение на 3,87 (2,31–5,43) мкм2 либо 15,9 % (9,5–22,3 %) в группе стресса];
-
- «Относительная площадь ядра нейрона» – зона РСГК V [уменьшение на 0,07 (0,04–0,11) либо 16,3 % (9,3–25,6 %) в группе стресса], РСАК II/III [уменьшение 0,07 (0,04–0,1) либо на 13,2 % (7,5–18,9 %) в группе стресса].
Для показателей «Площадь перикариона нейрона», «Площадь цитоплазмы перикариона нейрона», «Относительная площадь цитоплазмы перикариона нейрона», «ЯЦО нейрона» не обнаружено достоверных различий в исследуемых зонах ретроспленальной коры.
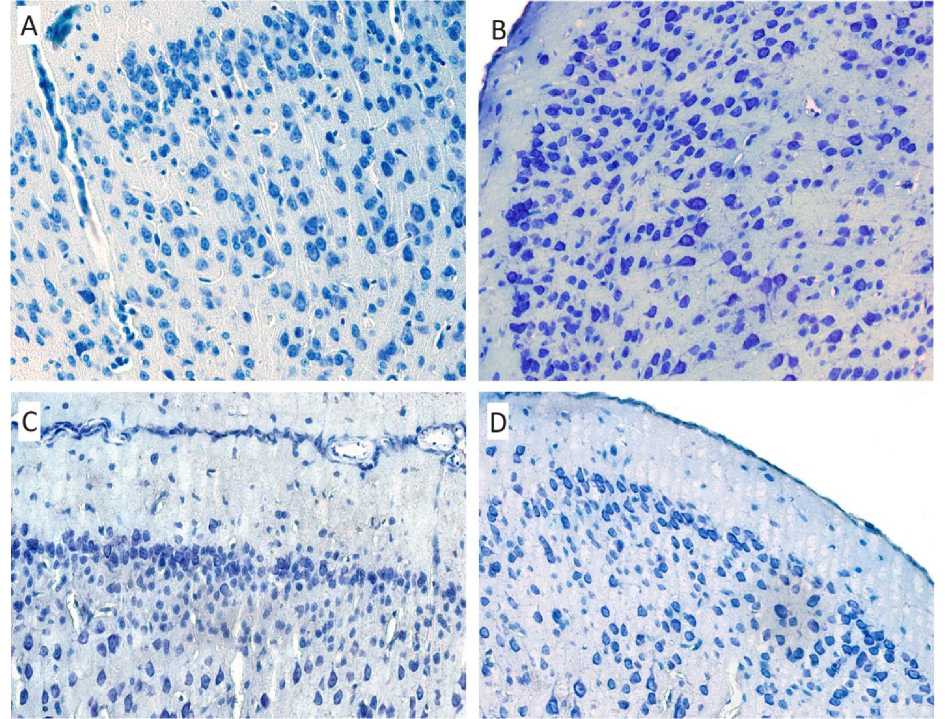
Рис. Гистологическое строение ретроспленальной коры головного мозга мыши, окраска тионином по методу Ниссля, ×400:
A, C – гранулярная ретроспленальная кора головного мозга; B, D – агранулярная ретроспленальная кора головного мозга; A, B – группа интактных мышей; C, D – группа стрессированных мышей
Таблица 1
Морфометрические показатели нейронов гранулярной ретроспленальной коры головного мозга II–III слоев
|
Показатель |
Интакт |
Стресс |
|
Периметр перикариона нейрона, мкм |
28,84 ± 7,43, 25,89 (23,36; 33,31) |
28,22 ± 5,56, 27,1 (24,69; 30,25) |
|
Периметр ядра нейрона, мкм |
18,32 ± 4,86, 16,5 (14,79; 21,64) |
17,80 ± 3,68, 17,04 (15,63; 19,18) |
|
Площадь перикариона нейрона, мкм2 |
42,10 ± 21,45, 32,93 (27,47; 52,08) |
38,89 ± 15,49, 35,14 (29,8; 42,34) |
|
Относительная площадь перикариона нейрона |
1,16 ± 0,46, 1,08 (0,86; 1,36) |
1,08 ± 0,37, 1,02 (0,78; 1,33) |
|
Площадь цито плазмы перикариона нейрона, мкм2 |
23,21 ± 12,00, 19,27 (14,61; 28,44) |
21,78 ± 8,80, 19,94 (16,09; 24,82) |
|
Относительная площадь цитоплазмы перикариона нейрона |
0,64 ± 0,27, 0,58 (0,45; 0,75) |
0,61 ± 0,23, 0,54 (0,43; 0,77) |
|
Площадь ядра нейрона, мкм2 |
18,88 ± 10,07, 14,68 (12,06, 23,98) |
17,11 ± 7,53, 15,53 (12,88; 18,61) |
|
Относительная площадь ядра нейрона |
0,52 ± 0,21, 0,48 (0,38; 0,61) |
0,47 ± 0,17, 0,46 (0,34;0,57) |
|
ЯЦО нейрона |
0,83 ± 0,19, 0,81 (0,69; 0,96) |
0,80 ± 0,19, 0,78 (0,67; 0,92) |
Примечание: данные в ячейках представлены в формате M ± SD, Me (Q1; Q3).
Таблица 2
|
Показатель |
Интакт |
Стресс |
|
Периметр перикариона нейрона, мкм |
34,75 ± 7,88, 34,39 (28,16; 40,75) |
38,06 ± 7,56, 37,36 (33,83; 42,37) |
|
Периметр ядра нейрона, мкм |
21,33 ± 4,86, 21,11 (17,68; 25,11) |
22,89 ± 4,10, 22,93 (20,4; 25,68) |
|
Площадь перикариона нейрона, мкм2 |
59,05 ± 25,39, 55,90 (38,94; 75,93) |
65,44 ± 23,23, 63,67 (50,52; 76,99) |
|
Относительная площадь перикариона нейрона |
1,03 ± 0,46, 0,99 (0,67; 1,29) |
0,86 ± 0,33, 0,8 (0,63; 1,03) |
|
Площадь цитоплазмы перикариона нейрона, мкм2 |
34,58 ± 16,43, 32,25 (21,79; 44,58) |
38,65 ± 15,86, 36,94 (27,94; 46,24) |
|
Относительная площадь цитоплазмы перикариона нейрона |
0,60 ± 0,29, 0,56 (0,39; 0,75) |
0,51 ± 0,21, 0,47 (0,36; 0,60) |
|
Площадь ядра нейрона, мкм2 |
24,47 ± 10,65, 22,56 (16,03; 32,04) |
26,79 ± 9,25, 25,97 (21,23; 31,22) |
|
Относительная площадь ядра нейрона |
0,43 ± 0,20, 0,40 (0,27; 0,56) |
0,36 ± 0,14, 0,33 (0,26; 0,42) |
|
ЯЦО нейрона |
0,75 ± 0,21, 0,73 (0,60; 0,87) |
0,74 ± 0,21, 0,72 (0,59; 0,88) |
Примечание: данные в ячейках представлены в формате M ± SD, Me (Q1, Q3).
Таблица 3
|
Показатель |
Интакт |
Стресс |
|
Периметр перикариона нейрона, мкм |
33,96 ± 6,41, 32,87 (29,19; 38,03) |
31,56 ± 4,60, 31,36 (28,13; 34,65) |
|
Периметр ядра нейрона, мкм |
21,21 ± 3,79, 20,83 (18,4; 23,65) |
19,54 ± 3,13, 19,27 (17,56; 21,46) |
|
Площадь перикариона нейрона, мкм2 |
55,49 ± 19,73, 49,45 (41,48; 65,45) |
48,44 ± 14,05, 47 (38,9; 56,21) |
|
Относительная площадь перикариона нейрона |
1,21 ± 0,41, 1,14 (0,92; 1,48) |
1,08 ± 0,31, 1,06 (0,88;1,27) |
|
Площадь цитоплазмы перикариона нейрона, мкм2 |
31,11 ± 12,39, 28,73 (22,65; 37,7) |
27,92 ± 9,02, 27,14 (21,39; 33,88) |
|
Относительная площадь цитоплазмы перикариона нейрона |
0,68 ± 0,26, 0,65 (0,50; 0,82) |
0,62 ± 0,19, 0,61 (0,49; 0,72) |
|
Площадь ядра нейрона, мкм2 |
24,38 ± 8,9, 22,10 (18,50; 28,40) |
20,51 ± 6,29, 19,59 (16,41; 23,39) |
|
Относительная площадь ядра нейрона |
0,53 ± 0,18, 0,50 (0,41; 0,66) |
0,46 ± 0,15, 0,45 (0,35; 0,53) |
|
ЯЦО нейрона |
0,82 ± 0,23, 0,81 (0,67; 0,94) |
0,76 ± 0,21, 0,72 (0,62; 0,89) |
Примечание: данные в ячейках представлены в формате M ± SD, Me (Q1, Q3).
Таблица 4
|
Показатель |
Интакт |
Стресс |
|
Периметр перикариона нейрона, мкм |
35,00 ± 8,46, 35,01 (28,81; 40,75) |
36,90 ± 9,70, 35,51 (30,54; 43,21) |
|
Периметр ядра нейрона, мкм |
20,87 ± 4,47, 21,10 (17,79; 23,82) |
21,37 ± 4,50, 21,85 (18,05; 24,56) |
|
Площадь перикариона нейрона, мкм2 |
60,87 ± 26,29, 57,92 (40,40; 78,65) |
65,07 ± 31,19, 57,10 (42,18; 85,92) |
|
Относительная площадь перикариона нейрона |
1,00 ± 0,44, 0,96 (0,65; 1,29) |
0,96 ± 0,45, 0,86 (0,61; 1,19) |
|
Площадь цитоплазмы перикариона нейрона, мкм |
37,14 ± 18,62, 32,94 (22,32; 50,28) |
40,61 ± 23,32, 33,73 (23,59; 54,47) |
|
Относительная площадь цитоплазмы перикариона нейрона |
0,61 ± 0,30, 0,55 (0,36; 0,82) |
0,59 ± 0,31, 0,51 (0,36; 0,74) |
|
Площадь ядра нейрона, мкм2 |
23,73 ± 9,39, 23,53 (16,42; 29,92) |
24,46 ± 9,56, 23,95 (17; 30,77) |
|
Относительная площадь ядра нейрона |
0,39 ± 0,16, 0,38 (0,27; 0,48) |
0,37 ± 0,16, 0,33 (0,25; 0,47) |
|
ЯЦО нейрона |
0,69 ± 0,19, 0,69 (0,56; 0,82) |
0,68 ± 0,23, 0,64 (0,51; 0,85) |
Примечание: данные в ячейках представлены в формате M ± SD, Me (Q1, Q3).
При этом имелись противоположные изменения во II/III слое агранулярной РСК в виде процессов атрофии нейронов, подтверждающихся достоверным уменьшением размеров нейронов – площади ядер на 15,9 % (9,5–22,3 %), периметра ядер на 7,8 % (4,5–11,1 %) и периметра перикарионов на 7,1 % (3,7–10,4 %), которым также сопутствовало снижение плотности нейронов в ви-
де снижения относительной площади их ядер на 13,2 % (7,5–18,9 %).
Таким образом, получены дополнительные морфологические доказательства участия ретро-спленальной коры головного мозга в ответе организма на хронический социальный стресс поражения в виде реципрокных изменений в ее субструктурах.
