Морфометрическая характеристика яиц птиц разных стратегий развития

Автор: Митяй Иван Сергеевич, Шатковская Оксана Вениаминовна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 4, 2015 года.
Бесплатный доступ
Изменчивость длины, диаметра и индексов формы яиц исследована на широком диапазоне птиц птенцово-выводкового спектра. Установлено, что птицы разных стратегий развития достоверно отличаются абсолютными размерами длины, диаметра и индексами формы яиц. Главными дискриминирующими факторами являются абсолютные размеры. В разграничении яиц по отрядам главными дискриминирующими факторами являются индексы клоакальной и инфундибулярной зон. Это указывает на необходимость использования радиусов поверхности яйца в описании его формы, а не ограничиваться использованием для данных целей лишь индекса удлиненности, который используется в большинстве литературных источников. В виду значительного влияния филогении на размеры и форму птичьих яиц их морфометрические параметры могут использоваться в качестве дополнительных критериев при решении вопросов эволюции и систематики птиц.
Выводковая, полувыводковая, полуптенцовая и птенцовая стратегии развития птиц, длина, диаметр и индексы формы яйца
Короткий адрес: https://sciup.org/146116602
IDR: 146116602 | УДК: 598.2.591.3.
Morphometric characteristic of bird eggs of different developmental modes
The variability of length, diameter and shape indexes of eggs was investigated in a wide range of altricial-precocial spectrum of birds. Birds with different developmental modes significantly differ by length, diameter and shape indexes of eggs. Length and diameter of eggs were the main discriminating factors. Indexes of cloacal and infundibular zones of eggs were the main discriminating factors between bird orders. The more complete description of the egg shape should contain besides the elongation indexes the values of the radii of the egg surface. Because the phylogeny significantly influences the size and shape of birds' eggs the morphometric parameters of the eggs can be used as additional criteria in the investigation of avian evolution and systematics.
Текст научной статьи Морфометрическая характеристика яиц птиц разных стратегий развития
Введение. Диапазон типов постэмбрионального развития птиц весьма разнообразен. Крайними полюсами спектра этих типов развития являются выводковые и птенцовые птицы. Такое распределение было совершено немецким орнитологом еще в XIX столетии (Oken, 1837). Между этими полюсами есть целый ряд промежуточных типов развития, значительно перекрывающихся между собой. Данное обстоятельство стало причиной значительного количества классификаций, которые активно разрабатывались в прошлом и нынешнем веке. А. Портман (1935) выделяет 7 групп развития, не давая им никакого названия. Позже М.М. Ницца (1962) выделяет 8 типов развития, которым дает следующие названия: выводковые (precocial) 1, 2, 3, 4; полувыводковые (semiprecocial), полунагнездные (semialtricial) 1, 2; нагнездные (altricial). Позже некоторые авторы сводили это количество до 4-5 (Skutch, 1976). В конце XX и начале XXI вв.
большинство орнитологов предпочитают использовать классификацию М.М. Ницца, иногда переименовывая крайние варианты в supercocial и superaltricial (Starck, 1993; Starck, Ricklefs, 1998; Dial, 2003, 2006; Schekkerman, 2008; Dyke, Kaiser, 2010).
В основе упомянутых классификаций лежит комплекс признаков, среди которых выделяют степень морфо-функционального развития птенцов, их активность, поведение, масштабы родительской опеки, срок нахождения в гнезде, на гнездовой территории и др. Максимальная форма выводковости (precocial 1) характерна для сорных кур (Megapodes), инкубация яиц которых происходит в специальных курганах из гниющих растительных остатков, а птенцы проявляют значительную самостоятельность. Максимально птенцовыми (altricial 2) считаются все Воробьинообразные (Passeriformes), Попугаеобразные (Psittaciformes) и Дятлообразные (Picifonnes), птенцы которых появляются голыми, слепыми и беспомощными. Все остальные типы развития в той или иной степени приближаются к указанным крайним вариантам. В литературе есть целый ряд публикаций, в которых описаны те или иные аспекты выводковости или птенцовости на основе сравнения значительного количества признаков. Отмечено, что стратегия развития и филогения оказывают доминирующее влияние на массу кладки (Birchard et al., 2013), различные репродуктивные характеристики птиц, включая состав яиц (Deeming, 2007а,b) и их энергетическую составляющую (Sibly et al., 2012). В некоторых работах, наряду с упомянутыми факторами, рассматривается влияние размеров самки и величины кладки на форму яйца (например, Olsen et al., 1994). Однако форма при этом рассматривалась лишь как отношение длины к диаметру, что, безусловно, недостаточно для полной характеристики.
В связи с этим, мы предприняли попытку поисков взаимосвязи типов развития с формой яйца при более расширенном использовании пространственных характеристик скорлупы яиц. Как известно, скорлупа играет очень важную роль в обеспечении условий для развития зародыша вне организма птицы. Она является вместилищем, где находится зародыш и питательные вещества для него, выполняет терморегуляторную, транспираторную и дыхательную функции. Ее механические свойства (прочность и толщина) также тесно связаны с формой яйца. Общеизвестно, что максимальной прочностью оболочки при минимальном расходе вещества обладает сфера. Яйцо, как топологически преобразованная сфера, в той или иной степени является отступлением от этого правила, что отражается на толщине скорлупы, количестве строительного материала, площади поверхности, количестве капилляров и т.д.
Анализ взаимосвязи упомянутых пространственных характеристик с типом развития птиц является целью данной статьи.
Методика. В качестве исходных независимых параметров для описания форм птичьих яиц были использованы три радиуса полярных зон, диаметр и длина. На основании их нами рассчитаны следующие индексы: традиционный индекс удлиненности - lei=L/D и три индекса, предложенных нами: 112=гД9; Iiz=ri/D; Icz=rc/D, где: Ijz, 1ц, Icz - индексы инфундибулярной, клоакальной и латеральной зон; р, гс, ц - радиусы дуг, L - длина, D - диаметр. Методика снятия параметров и вычисления индексов была детально изложена нами ранее (Митяй, 2014). Наряду с индексами формы использовались полиномные коэффициенты, вычисленные Л.И. Францевичем (2010) для нашей базы данных.
Распределение птиц по моделям развития приведено согласно классификации Starck J.M., Ricklefs R.E. (1998), из которой были взяты четыре основные группы: выводковая, полувыводковая, полуптенцовая и птенцовая. Изменчивость морфологических параметров яиц исследована стандартными методами линейного дискриминантного анализа (Афифи, Эйзен, 1982). Для каждого варианта набора параметров яйца определено общее значение статистики Уилкса (А) с учетом всех задействованных переменных, рассчитаны статистики Фишера (F) и уровни значимости (р). Приводятся статистические характеристики, касающиеся каждой из дискриминантных переменных, и построены канонические переменные. Проверка значимости переменных осуществлена с использованием критерия согласия Пирсона (/2). Нами также проведен дискриминантный анализ в пределах каждой модели развития для того, чтобы проанализировать каким образом распределяются яйца по абсолютным размерам индексов формы среди представителей разных отрядов. Группирующей переменной в этом случае был отряд птиц.
Вычисления проведены с помощью статистического пакета Statistica for Windows, версия 7 (StatSoft, Inc., США). Достоверность разницы между показателями определялась с помощью One-way ANOVA и Kruskal-Wallis tests (PAST) (Hammer et al., 2001). Объем материала и основные морфометрические характеристики приведены в табл. 1.
Данные о количестве исследованных яиц птиц с разными стратегиями развития и результаты базовой статистической обработки параметров яиц также приведены в табл. 1.
Таблица 1
Объем материала и основные статистические характеристики параметров яиц у представителей разных стратегий развития
|
Параметры |
Стратегия развития |
Valid N |
Mean |
Min |
Max |
Std. Dev. |
Coeff. var |
|
L |
выводковая |
3215 |
53,7 |
21,9 |
168,2 |
19,32 |
36,0 |
|
птенцовая |
6786 |
24,6 |
11,8 |
116,3 |
10,10 |
41,1 |
|
|
полувыводковая |
1338 |
54,4 |
24,8 |
134,3 |
15,35 |
28,2 |
|
|
полуптенцовая |
2047 |
51,5 |
28,5 |
103,9 |
11,17 |
21,7 |
|
|
D |
выводковая |
3216 |
37,9 |
18,5 |
131,2 |
12,18 |
32,1 |
|
птенцовая |
6787 |
18,0 |
10,1 |
64,9 |
6,29 |
35,0 |
|
|
полувыводковая |
1338 |
38,4 |
18,7 |
87,6 |
9,92 |
25,8 |
|
|
полуптенцовая |
2048 |
39,7 |
21,0 |
78,9 |
7,98 |
20,1 |
|
|
ICZ |
выводковая |
3892 |
0,27 |
0,08 |
0,45 |
0,08 |
29,0 |
|
птенцовая |
8611 |
0,29 |
0,11 |
0,49 |
0,06 |
20,6 |
|
|
полувыводковая |
1611 |
0,24 |
0,08 |
0,45 |
0,06 |
26,8 |
|
|
полуптенцовая |
2376 |
0,33 |
0,13 |
0,47 |
0,06 |
16,8 |
|
|
I1Z |
выводковая |
3892 |
0,96 |
0,59 |
1,75 |
0,16 |
17,0 |
|
птенцовая |
8611 |
0,89 |
0,58 |
1,80 |
0,15 |
16,4 |
|
|
полувыводковая |
1611 |
1,01 |
0,60 |
1,80 |
0,21 |
21,1 |
|
|
полуптенцовая |
2376 |
0,77 |
0,58 |
1,47 |
0,09 |
11,8 |
|
|
liz |
выводковая |
3892 |
0,46 |
0,25 |
0,50 |
0,04 |
8,2 |
|
птенцовая |
8611 |
0,47 |
0,28 |
0,50 |
0,02 |
5,3 |
|
|
полувыводковая |
1611 |
0,47 |
0,33 |
0,50 |
0,02 |
4,9 |
|
|
полуптенцовая |
2376 |
0,45 |
0,28 |
0,50 |
0,03 |
7,1 |
Результаты и обсуждение. Представители разных стратегий развития различаются как абсолютными размерами яиц (L, D), так и их формой, которая описывается индексами формы яйца и коэффициентами полиномы (табл. 2). Однако если в дискриминантном анализе используются только индексы формы яйца или только коэффициенты полинома, качество определения принадлежности яиц ухудшается. Дискриминантные функции в этом случае позволяют правильно определить принадлежность 54-56% яиц. Использование только абсолютных размеров (L, D) дает хороший результат верных определений в целом (72%). Однако приводит к ухудшению определения принадлежности яиц по некоторым группам птиц. В частности, яйца птиц с полувыводковой стратегией развития не определяются вовсе. Компромиссным вариантом является использование в дискриминантном анализе и абсолютных размеров яиц (L, D), и индексов формы. В этом случае дискриминантные функции, учитывающие вариацию и ковариацию указанных параметров, позволяют правильно определить принадлежность 72% яиц. На много лучше, в сравнении с вышеуказанными вариантами, определяется принадлежность яиц к определенной стратегии развития (табл. 2). Этот вариант набора параметров, как наиболее полно дискриминирующий яйца птиц с разными стратегиями развития, будет использован в дальнейшем анализе.
Таблица 2 Результативность определения яиц по разным наборам параметров
|
Параметры |
A |
F |
p,% |
% верных определений* |
||||
|
1 |
2 |
3 |
4 |
5 |
||||
|
L, D, Icz, 1ц, 1ц |
0,28 |
1448,16 |
<0,0001 |
72 |
46 |
94 |
23 |
71 |
|
L, D |
0,33 |
3327,76 |
<0,0001 |
72 |
56 |
95 |
0 |
66 |
|
Icz, Ilz, liz, % |
0,72 |
383,02 |
<0,0001 |
54 |
12 |
90 |
8 |
28 |
|
ko, kb k2, k3** |
0,71 |
409,9 |
<0,0001 |
56 |
24 |
90 |
9 |
20 |
Примечание. *1 - общий процент, 2 - выводковые, 3 - птенцовые, 4 -полувыводковые, 5 -полуптенцовые; ** к0, кь к2, к3 - коэффициенты полиномы.
Значение статистики Уилкса (А) для дискриминирующих переменных отличаются незначительно (от 0,28 до 0,34) (табл. 3).
Таблица 3 Результаты дискриминантного анализа яиц представителей разных стратегий развития
|
Параметры |
Wilks' Lambda |
Partial Lambda |
F-remove |
Toler. |
1-Toler. |
|
L |
0,31 |
0,90 |
500,92 |
0,03 |
0,97 |
|
D |
0,34 |
0,82 |
1005,04 |
0,03 |
0,97 |
|
Icz |
0,29 |
0,95 |
227,46 |
0,88 |
0,12 |
|
I1Z |
0,28 |
0,99 |
66,94 |
0,48 |
0,52 |
|
Ilz |
0,29 |
0,97 |
158,69 |
0,60 |
0,40 |
Примечание. р<0,0001.
С одной стороны, это свидетельствует о том, что все параметры примерно в одинаковой степени участвуют в определении принадлежности яиц птицам разных моделей развития, с другой - все рассматриваемые переменные важны для проведения дискриминантного анализа. Вместе с тем из рассмотренных дискриминирующих переменных наибольшее значение в разграничении имеют абсолютные размеры яиц. Как длина, так и диаметр яиц птиц имеют высокий коэффициент вариации (табл. 1). Из рассмотренных индексов формы наиболее изменчив индекс клокальной зоны и наименее изменчив индекс инфундибулярной зоны (табл. 1).
Коэффициенты множественной корреляции параметров, анализируемых в модели, свидетельствуют, что наибольше связаны с остальными параметрами длина и диаметр яйца (табл. 3). Наименее сильно связан с другими параметрами индекс клоакальной зоны.
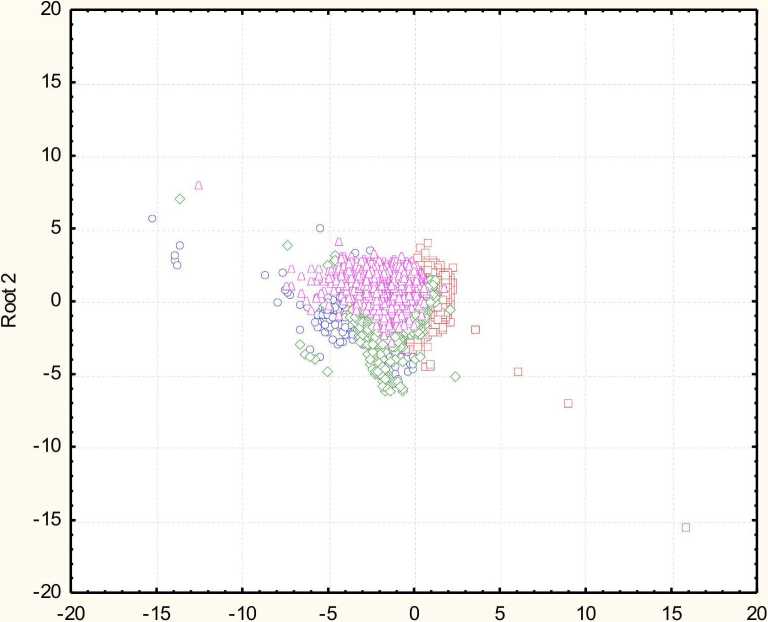
Распределение изученных экземпляров в пространстве первых двух канонических переменных показано на рис. 1. Из рисунка следует, что яйца представителей разных стратегий развития не разграничиваются отчетливо по значениям ни 1-й, ни 2-й канонических переменных.
Root 1 vs. Root 2
0 precocial
□ altricial о semi-precocial semi-altricial
Root 1
Рис. 1 . Распределение птиц с разной стратегией развития в пространстве первых двух канонических переменных
Можно наблюдать лишь некоторое смещение показателей канонических переменных для каждой модели развития относительно некоторого общего центра. Это свидетельствует в пользу консервативности размеров и формы яиц для птиц в целом. Несколько с других позиций к подобным выводам пришли Риклефс и Старк (1998) при исследовании вариации массы яиц. Вероятно, изменчивость размера и формы яиц ограничена рядом факторов, среди которых можно выделить размер птиц, как летающих животных, размеры кладки, общие репродуктивные усилия, направленные на откладывание яиц (Olsen, Cunningham, 1994), характеристики яичной скорлупы (Birchard, Deeming, 2009) и др.
Среди птиц с разной стратегией развития наилучшим образом определяются яйца птенцовых (94%) и полуптенцовых птиц (72%), и хуже - выводковых (40%) и полувыводковых (24%). Для исследуемого комплекса признаков вычислены классификационные функции (табл. 4). Коэффициенты классификационных функций, вероятно, можно предложить для практического применения в дальнейших исследованиях.
Таблица 4 Классификационные функции, построенные по размерам и индексам формы яиц
|
Параметры |
выводковые |
птенцовые |
полувыводковые |
полуптенцовые |
|
L |
2,55 |
2,76 |
2,55 |
2,19 |
|
D |
-2,64 |
-3,22 |
-2,62 |
-2,08 |
|
Icz |
132,87 |
139,12 |
125,31 |
139,86 |
|
hz |
-17,05 |
-20,66 |
-16,75 |
-19,11 |
|
hz |
760,65 |
765,13 |
782,38 |
745,10 |
|
Constant |
-203,64 |
-195,63 |
-214,04 |
-200,11 |
Рассмотрим результаты дискриминантного анализа в пределах каждой модели развития. Из птенцовых птиц по анализируемым параметрам определяются яйца представителей четырех отрядов: Passeriformes (99%), Pelecaniformes (99%), Columbiformes (71%), Coraciformes (70%). Большинство яиц представителей Cuculiformes, Apodiformes, Pssitaciformes, Upupiformes, которые также имеют птенцовую модель развития, определяются как яйца представителей Passeriformes. Яйца представителей Phoenicopteriformes определяются, главным образом, как яйца представителей Pelecaniformes. Если для птиц в целом главным дискриминирующим признаком являются абсолютные размеры, то для птенцовых птиц - это индекс клоакальной зоны.
Root 1 vs. Root 2
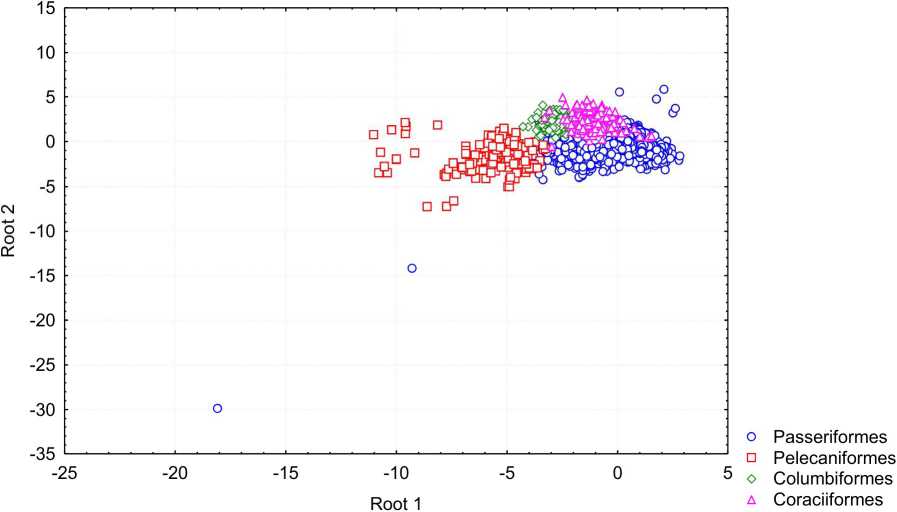
Рис. 2. Распределение птиц с птенцовой стратегией развития в пространстве первых двух канонических переменных
Распределение экземпляров в пространстве первых двух канонических переменных приведено на рис. 2. 1-я каноническая переменная отчетливо выделяет представителей Pelecanifonnes. Значения 2-ой канонической переменной менее четко разграничивают Pelecaniformes, Columbifonnes и Coraciformes.
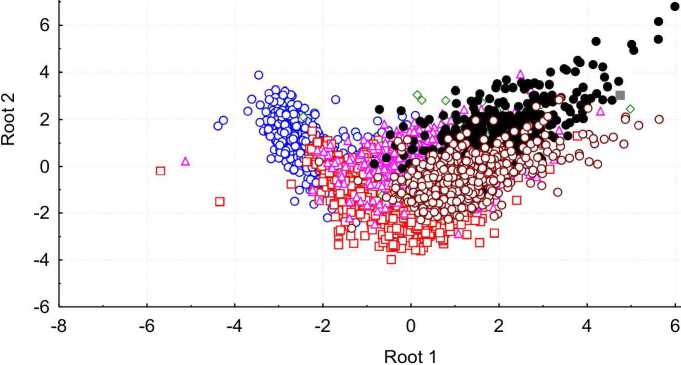
Из выводковых птиц по абсолютным размерам и индексам формы определяются яйца представителей семи отрядов: Charadriifonnes (84%), Galliformes (79%), Gaviifonnes (72%), Gruiformes (49%), Podicipediformes (68%), Tinamifonnes (50%), Anseriformes (59%). He определяются яйца представителей Columbiformes и плохо определяются яйца представителей Struthioniformes (2,5%). Главным дискриминирующим признаком среди выводковых птиц является индекс инфундибулярной зоны. В пространстве первых двух канонических переменных ни один из отрядов не занимает отдельного региона (рис. 3). Однако распределение исследуемых экземпляров не является однородным; для каждого отряда можно выделить обособленный участок. По значениям 1-й канонической переменной отличаются представители Charadriifonnes от представителей Anseriformes и Podicipediformes, по значениям 2-й канонической переменной отличаются представители Galliformes и Podicipediformes.
Root 1 vs. Root 2
о Charadriiformes
-
□ Galliformes
-
о Gaviiformes
-
a Gruiformes
-
• Podicipediformes
8 ■ Tinamiformes о Anseriformes
Рис. 3 . Распределение птиц с выводковой стратегией развития
в пространстве первых двух канонических переменных
Root 1 vs. Root 2
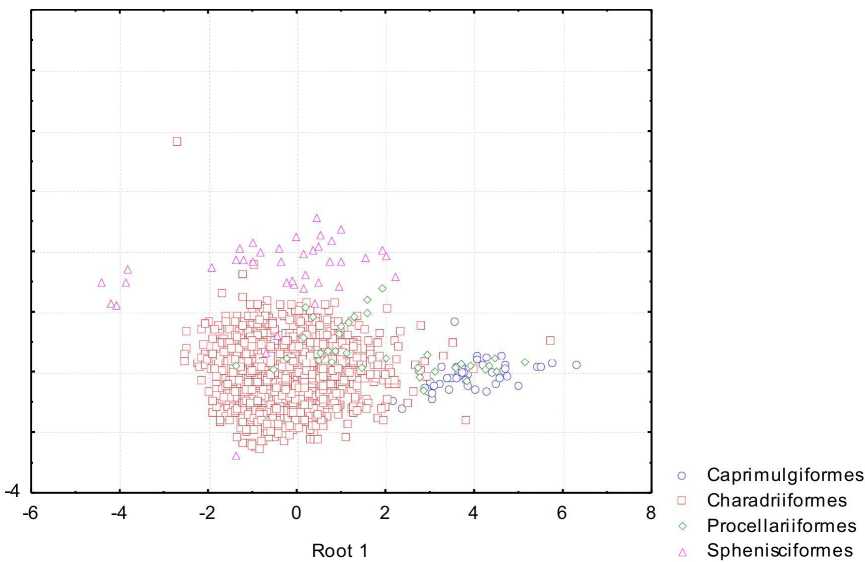
Рис. 4 . Распределение птиц с полувыводковой стратегией развития в пространстве первых двух канонических переменных
У полувыводковых птиц хорошо определяются яйца представителей Caprimulgiformes (96%), Charadriifonnes (99%), Spheniscifonnes (82%), хуже представителей Procellariiformes (6%). Главными дискриминирующими признаками являются индексы клоакальной и инфундибулярной зон. По значениям 1-й канонической переменной отличаются Caprimulgiformes от Charadriiformes и Sphenisciformes (рис. 4), по значениям 2-й канонической переменной отличия есть между Spheniscifonnes и представителями других отрядов.
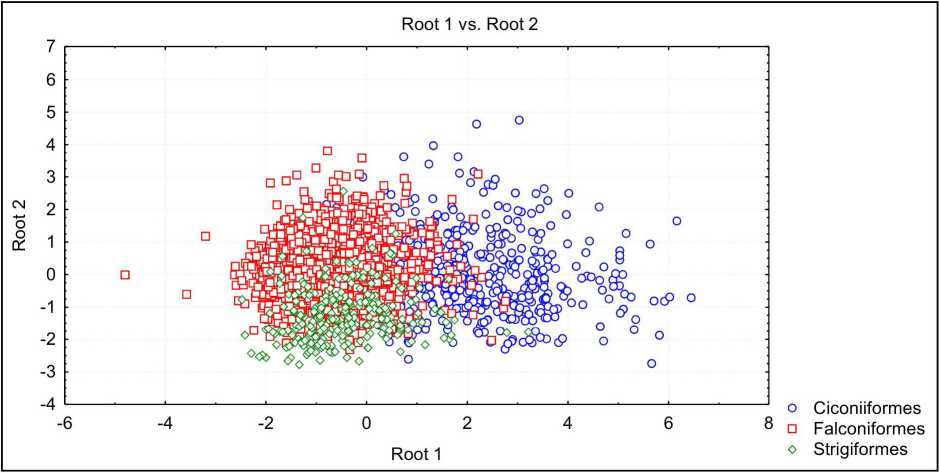
Рис. 5 . Распределение птиц с полуптенцовой стратегией развития в пространстве первых двух канонических переменных
У полуптенцовых птиц хорошо определяются яйца представителей Falconiformes (96%) и Ciconiifonnes (76%), хуже представителей Strigiformes (28%), большинство из них определяются как яйца представителей Falconiformes. Не определяется принадлежность яиц представителям Charadriiformes. Основным разграничивающим признаком у полуптенцовых птиц является индекс инфундибулярной зоны. По значениям 1-й канонической переменной Ciconiiformes отличаются от Falconiformes и Strigiformes (рис. 5).
Заключение. Из приведенных выше результатов исследования видно, что при разграничении отрядов в пределах каждой модели развития главными дискриминирующими факторами являются не столько абсолютные размеры, как индексы формы яйца. Это указывает на важность использования радиусов кривизны яйца для описания его формы, а не ограничиваться использованием для данных целей лишь индекса удлиненности, который обычно используется в литературе (Olsen, Cunningham, 1994).
В виду того, что яйца птиц разграничиваются не только по принадлежности к определенным моделям развития птиц, но также и по отрядам, параметры формы птичьих яиц, по нашему мнению, могут использоваться в качестве дополнительного критерия при решении вопросов систематики и филогении птиц.
Митяй И.С. Морфометрическая характеристика яиц птиц разных стратегий развития / И.С. Митяй, О.В. Шатковская // Вести. ТвГУ. Сер.: Биология и экология. 2015. № 4. С. 109-120.
Список литературы Морфометрическая характеристика яиц птиц разных стратегий развития
- Афифи А., Эйзен С. 1982. Статистический анализ: Подход с использованием ЭВМ. М.: Мир. 488 с.
- Митяй И.С. 2014. Описание форм птичьих яиц с помощью геометрических эталонов//Вестн. ТвГУ. Сер.: Биология и экология. № 1. С. 125-147.
- Францевич Л.И. 2010. Планиметрия параметров формы птичьего яйца . Режим доступа: http://www.biometrica.tomsk.ru/planirus.htm (дата обращения: 25.09.2015)
- Birchard G.F., Deeming D. C. 2009. Avian eggshell thickness: scaling and maximum body mass in birds//Journal of Zoology. V. 279. P. 95-101.
- Birchard G.F., Ruta M., Deeming D.C. 2013. Evolution of parental incubation behaviour in dinosaurs cannot be inferred from clutch mass in birds//Biology Letters. V. 9. P. 1-5.
- Deeming D.C. 2007a. Effects of phylogeny and hatchling maturity on allometric relationships between female body mass and the mass and composition of bird eggs//Avian and Poultry Biology Reviews. V. 18. P. 21-37.
- Deeming D.C. 2007b. Allometry of mass and composition in bird eggs: effects of phylogeny and hatchling maturity//Avian and Poultry Biology Reviews. V. 18. P. 71-86.
- Dial K.P. 2003. Evolution of avian locomotion: correlates of flight style, locomotor modules, nesting biology, body size, development, and the origin of flapping flight//Auk. V. 120 (4). P. 941-952.
- Dial K.P. 2006. Patterns among avian flight style, nesting biology, development, body size, and locomotor modularity//Acta Zoologica Sinica. V. 52 (Supplement). P. 502-504.
- Dyke G.J, Kaiser G.W. 2010. Cracking a developmental constraint: Egg size and bird evolution//Records of the Australian Museum. V. 62. P. 207-216.
- Hammer O., Harper D.A.T., Ryan P.D. 2001. PAST: Paleontological Statistics software package for education and data analysis//Palaeontologia Electronica. V. 4. № 1. P. 1-9.
- Nice M.M. 1962. Development of behavior in precocial birds//Transactions of the Linnean Society of New York. V. 8. P. 1-211.
- Olsen P.D., Cunningham R.B. 1994. Avian egg morphometrics: Allometric models of egg volume, clutch volume and shape//Australian Journal of Zoology. V. 42. P. 307-321.
- Oken L. 1837. Allgemeine Naturgeschichte für alle Stände. Bd. 7 Abt.1. Stuttgar: Hoffmann’sche Verlagsbuchhandlung. 685 S.
- Portmann A. 1935. Die Ontogenese der Vögel als Evolutionsproblem//Acta Biotheoretica. Bd. 1A. S. 59-90.
- Schekkerman H. 2008. Precocial problems: Shorebird chick performance in relation to weather, farming and predation. Alterra Scientific Contributions. 228 p.
- Sibly R.M., Witt C.C., Wright N.A., Venditti C., Jetze W., Brown J.H. 2012. Energetics, lifestyle, and reproduction in birds//Proceedings of the National Academy of Sciences of the USA. V. 109. P. 937-1941.
- Skutch A. F. 1976. Parent birds and their young. Austin and London: University of Texas press. 503 p.
- Starck J.M. 1993. Evolution of avian ontogenies//Current ornithology. 1993. V. 10. P. 275-366.
- Starck J.M., Ricklefs R.E. 1998. Avian Growth and Development. Evolution within the Altricial-Precocial Spectrum. New York: Oxford University Press. 380 p.
