Морфометрическая и молекулярно-генетическая дифференциация линий и семей медоносной пчелы Apis mellifera caucasica L., разводимых в районе Большого Сочи

Автор: Форнара М.С., Крамаренко А.С., Свистунов С.В., Любимов Е.М., Сокольский С.С., Зиновьева Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Геномное сканирование, контроль происхождения
Статья в выпуске: 6 т.50, 2015 года.
Бесплатный доступ
Создание специализированных линий - один из приемов генетического совершенствования и сохранения биоразнообразия пород и популяций медоносной пчелы. Серая горная кавказская порода ( Apis mellifera caucasica L.) выделяется среди прочих по комплексу хозяйственно полезных признаков (самый длинный хоботок, высочайшая работоспособность и продуктивность, использование широкого видового состава медоносов, чрезвычайно слабая ройливость, миролюбие и др.). Целью настоящего исследования была сравнительная оценка разнообразия и степени дифференциации линий A . m . caucasica на основании данных морфометрического анализа и микросателлитов (MС). Материалом для исследований служили рабочие пчелы пяти линий (I - линия № 19, с. Медовеевка; II - линия № 13, ПГТ Красная Поляна; III - линия № 12, ПГТ Красная Поляна; IV - линия № 49, с. Калиновое Озеро; V - линия № 34, с. Аибга; n = 728) с пяти пасек в районе Большого Сочи (Краснодарский край). Для морфометрического анализа измеряли длину хоботка (ДХ, мм), ширину 3-го тергита (ШТ, мм) и рассчитывали кубитальный индекс (КИ). Молекулярно-генетические исследования проводили по семи локусам МС (A024, A88, A113, AP043, HB-C16-05, HB-THE-03, HB-C16-01). Степень межсемейной изменчивости по морфометрическим показателям определяли методом двухфакторного иерархического дисперсионного анализа. Генетические различия между семьями по МС оценивали по показателям F st при парном сравнении. Полученную матрицу использовали для PCA-анализа (principal component analysis). Для количественной оценки межсемейной изменчивости внутри линий рассчитывали индексы F st, R st (AMOVA). Степень дифференциации линий по морфометрическим признакам определяли посредством расчета евклидовых дистанций. Полученные значения использовали для построения дендрограммы подобия методом одиночной связи (single linkage) в алгоритме иерархической кластеризации. Степень дифференциации линий по МС оценивали на основании расчета значений генетических дистанций по М. Nei. Дендрограмму подобия строили с помощью метода UPGMA. Для расчетов использовали программное обеспечение STATISTICA, GenAlEx (v. 6.5.1), PAST (v. 3.03). Результаты морфометрического анализа показали наличие достоверных различий между линиями по ДХ и Ш3Т, тогда как в отношении КИ различий не обнаружили. Установлена большая гетерогенность в отношении изучаемых признаков в линиях II и V и, напротив, большая консолидация в линиях III и IV. Пчелы линии I существенно отличались от остальных по обоим показателям, однако характеризовались значительными межсемейными различиями по ДХ. Анализ МС профилей выявил сходные тенденции при оценке степени внутрисемейной и межсемейной изменчивости. В линии I наблюдается избыток гетерозигот (F is = -0,048), что можно рассматривать как указание на высокую гетерогенность. Пчелы этой линии характеризовались минимальной индивидуальной (F it = 0,052) и максимальной межсемейной (F st = 0,124) изменчивостью. В линиях II-V наблюдали дефицит гетерозигот (F is= 0,062-0,128), относительно высокую индивидуальную изменчивость (F it = 0,143-0,189) и более низкие по отношению к линии I значения межсемейной изменчивости (F st = 0,095-0,104). Наименьшие межсемейные различия были отмечены в линиях III и IV (F st 0,096 и 0,095). Анализ дифференциации изучаемых линий по морфометрическим признакам и МС выявил различия в структуре генеалогического дерева. Дендрограмма, построенная по МС, была отражением географической удаленности изучаемых линий. Структура генеалогического дерева, построенного по морфометрическим признакам, не отражала ни географическую общность (различия) происхождения, ни сходство (различия) в хозяйственно полезных признаках у изучаемых линий. Таким образом, при исследовании морфометрических показателей и МС обнаружены сходные тенденции в оценке внутрисемейной и внутр
Медоносная пчела, днк-маркеры, морфометрия, микросателлиты, биоразнообразие
Короткий адрес: https://sciup.org/142133641
IDR: 142133641 | УДК: 638.123.54:575.174:[577.2.08+591.4.08 | DOI: 10.15389/agrobiology.2015.6.776rus
Morphometric and molecular genetic differentiation of Apis mellifera caucasica L. honey bee lines reared in Sochi region
Creating specialized lines is one of the techniques of genetic improvement and conservation of breeds and populations of the honeybee. The aim of this study was a comparative assessment of the diversity and differentiation degree of A. m. caucasica lines based on morphometric analysis and microsatellites (MS). Material for this study was the worker bees of Gray Mountain Caucasian breed (lines I-V, n = 728) which were selected in five apiaries in the Greater Sochi of Krasnodar krai. Morphometric analysis included such measurements as the length of the proboscis (LP, mm), width of third tergite (W3T, mm) and cubital index (CI). Molecular genetic studies were based on seven MS loci (A024, A88, A113, AP043, HB-C16-05, HB-THE-03, HB-C16-01). The level of variation between families for morphometric parameters was determined by two-way hierarchical analysis of variance. Genetic differences between families for MS were estimated by paired comparison of F st values. F st matrix was used for PCA-analysis. To determine the quantitative estimation of variation between families within lines we calculated F st, R st (AMOVA). The degree of line's differentiation for morphometric characters was evaluated by calculating the Euclidean distances. The obtained values were used to construct a dendrogram of similarity by a single bond (single linkage) of the hierarchical clustering algorithm. The differentiation of lines for MS was based on calculating the values of Nei genetic distances. Similarity dendrogram was constructed using the method of UPGMA. We performed summary statistic using the software STATISTICA, GenAlEx (v. 6.5.1), PAST (v. 3.03). Morphometric analysis showed the presence of significant differences between the lines for LP and W3T whereas there was no difference between the lines for CI. The greater heterogeneity concerning studied traits in the lines II and V was revealed, and on the contrary, there was more consolidation in the lines III and IV. Bees of line I differed significantly from the rest of the lines on both traits, but they were characterized by significant differences between families in LP. Analysis of MS profiles showed similar trends in assessing the level of intra- and interfamily variability. We observed an excess of heterozygotes in the line I (F is = -0.048), which can be considered as an indication of the high heterogeneity. Bees of this line were characterized by a minimal individual (F it = 0.052) and the maximal interfamily variability (F st = 0,124). Lines II-V were characterized by a deficiency of heterozygotes (F is = 0.062-0.128), a relatively higher individual variability (F it = 0.143-0.189) and lower values of interfamily variability comparing to line I (F st = 0.095-0.104). The lowest interfamily differences were observed in the lines III and IV (F st = 0.096 and 0.095, respectively). Analysis of the differentiation of the studied lines for morphometric characteristics and MS revealed differences in the structure of the family tree. The dendrogram based on MS data is a reflection of the geographical origin of these lines. The structure of the family tree, based on morphometric characters, does not reflect the geographic closeness (differences) of origin or similarities (differences) in the economically useful traits of studied lines. Thus, the results of our studies of the morphometric parameters and MS show similar trends in assessing intra- and interline variation, but there are differences in assessing differentiation of lines using two methods. In the future complex approach will allow to identify not only breeds of bees with high accuracy, but also smaller taxonomic units. It is hoped that the research results in general can be used in breeding work to restore the purity of the honeybee breeds.
Текст научной статьи Морфометрическая и молекулярно-генетическая дифференциация линий и семей медоносной пчелы Apis mellifera caucasica L., разводимых в районе Большого Сочи
В представляемой работе мы показали, что структура генеалогиче- ского дерева, построенного по морфометрическим признакам, не отражала ни географическую общность происхождения, ни сходство в хозяйственно полезных признаках у изученных линий. Следовательно, для их адекватной характеристики и дифференциации необходимо сочетание морфометрических и молекулярно-генетических методов. Использованный нами комплексный подход позволит с высокой точностью определять не только породы пчел, но и более мелкие таксономические группы. Полученные результаты могут быть использованы в работе по восстановлению чистоты пород медоносных пчел.
Целью настоящей работы стала сравнительная оценка разнообразия и степени дифференциации линий Apis mellifera caucasica на основании данных морфометрического анализа и микросателлитов.
Методика. Исследования проводили в 2014 году. В качестве материала использовали рабочих пчел серой горной кавказской породы различных линий, разводимых на пяти пасеках в районе Большого Сочи (Краснодарский край): I — линия ¹ 19 (с. Медовеевка), II — линия ¹ 13 (ПГТ Красная Поляна), III — линия ¹ 12 (ПГТ Красная Поляна), IV — линия ¹ 49 (с. Калиновое Озеро) и V — линия ¹ 34 (с. Аибга). Для анализа отбирали 5 семей каждой линии по 22-30 пчел в семье (25 семей, n = 728), в том числе I — n = 149, II — n = 147, III — n = 147, IV — n = 137, V — n = 148. Пчел консервировали в 96 % спирте и хранили при +4 ° C.
Измеряли длину хоботка (ДХ, мм), ширину 3-го тергита (ШТ, мм) и рассчитывали кубитальный индекс (КИ) как отношение между большей и меньшей сторонами третьей кубитальной ячейки переднего крыла (8).
ДНК выделяли с помощью экстракции перхлоратом (28), материалом служила голова рабочей пчелы. Семь локусов микросателлитов (МС) — A024, A88, A113, AP043, HB-C16-05, HB-THE-03, HB-C16-01 амплифици-ровали в одной мультиплексной реакции с использованием флуоресцентно меченных праймеров. Реакции проводили в конечном объеме 15 мкл. В пробирки вносили по 14 мкл реакционной смеси (1 мкл 10½ ПЦР-буфера, 0,5 мкл 2 мМ раствора dNTPs, 0,7 мкл 100 мМ раствора DMSO, 4,6 мкл 10 мМ смеси праймеров, 0,1 мкл Таq-полимеразы, 7,1 мкл бидистиллиро-ванной воды) и добавляли 1 мкл (50-100 нг) исследуемой геномной ДНК. Состав ПЦР-буфера: 16,6 мМ (NH 4 ) 2 SO 4 , 67,7 мМ Трис-НС1 (рН = 8,8), 0,1 объема Tween 20. ПЦР выполняли в следующем режиме: начальная денатурация 8 мин при 94 ° С; отжиг 2 мин при 55 ° С; полимеризация 2 мин при 72 ° С; 30 циклов — денатурация 30 с 94 ° С, отжиг 35 с при 55 ° С, элонгация 40 с при 72 ° С; элонгация 20 мин при 72 ° С. Разделение продуктов ПЦР выполняли на генетическом анализаторе АВI3130xl («Applied Biosystems», США) с использованием внутреннего стандарта ROX. Электрофореграммы обрабатывали с помощью программного обеспечения GeneScan v. 3.7 и Genotyper v. 3.7 («Applied Biosystems», США).
Межсемейную изменчивость по морфометрическим показателям оценивали методом двухфакторного иерархического дисперсионного анализа с помощью программного обеспечения Statistica v. 6 (29). В качестве фиксированных эффектов использовали принадлежность к линии и семье (иерархический подчиненный фактор). Рассчитывали Mst:
F - 1
F - 1 + n ’
M st =
где F — дисперсионное отношение, n* — среднее геометрическое объема выборок. Величина Mst может принимать значения от 0 до 1 и оценивать меру морфологической дифференциации между отдельными популяциями и, по сути, представляет собой аналог Fst (30).
Генетические различия между семьями медоносной пчелы в отношении МС оценивали по показателю Fst (31) при парном сравнении и на основании генетических дистанций по M. Nei (32). Полученные попарные матрицы использовали для построения распределения центроидов семей в пространстве первых двух главных координат (PCA, principal component analysis). Для количественной оценки степени межсемейной изменчивости внутри линий рассчитывали идексы Fst, Rst (AMOVA) (33), используя программное обеспечение GenAlEx v. 6.5 (34).
Степень дифференциации пяти изучаемых линий A. m. caucasica по морфометрическим признакам оценивали посредством расчета евклидовых дистанций. Полученные значения использовали для построения дендрограммы подобия методом одиночной связи (single linkage) в алгоритме иерархической кластеризации (29). Оценку степени дифференциации линий по МС проводили на основании расчета значений генетических дистанций по M. Nei (32). Дендрограмму подобия строили методом UPGMA при помощи программного обеспечения PAST (v. 3.03). Дендрограмму проверяли на устойчивость топологии кластеров посредством проведения 999 перму-таций bootstrap-методом.
Результаты. При формировании каждая из изученных линий пчел была строго ориентирована на получение материала с заданными хозяйственно полезными признаками. Матки линии III обладают высокой яйценоскостью в период подготовки к основному медосбору, а семьи — повышенной продуктивностью по меду. Линии IV и V характеризуются ранним развитием по сравнению с линиями I, II и III. Пчелы линии IV менее зимостойки, в то время как линия II выделяется повышенной, а линия I — высокой зимостойкостью.
Были выявлены достоверные различия между линиями по двум из трех оцененных признаков — длине хоботка и ширине 3-го тергита, тогда как по значениям кубитального индекса различий между линиями мы не обнаружили (табл. 1). У пчел линии I по сравнению с другими изучаемыми линиями величина ДХ оказалась на 0,05-0,07 мм меньше (p < 0,001), а Ш3Т — на 0,04-0,11 мм больше (p < 0,001). Максимальную длину хоботка отмечали у пчел линии III. У пчел линий III и IV Ш3Т была на 0,08-0,11 мм меньше, чем у других линий (p < 0,001). Обнаруженные различия могут быть следствием разной стратегии при селекционно-племенной работе с этими линиями.
1. Морфометрические показатели у различных линий медоносных пчел серой горной кавказской породы ( Apis mellifera caucasica L.) ( X ± x , Краснодарский край, 2014)
|
Линия |
Морфометрический признак |
||
|
ДХ, мм |
1 Ш3Т, мм 1 |
КИ |
|
|
I |
6,88±0,006 a, b, c, d |
4,72±0,010a, b, c, d |
0,502±0,008 |
|
II |
6,94±0,005b |
4,67±0,010b, e, f |
0,509±0,007 |
|
III |
6,95±0,004c, i |
4,64±0,008c, f, g |
0,513±0,007 |
|
IV |
6,94±0,004d, i |
4,61±0,009d, e, h |
0,515±0,008 |
|
V |
6,93±0,005a |
4,68±0,010a, g, h |
0,524±0,008 |
П р и м еч а ни е. ДХ — длина хоботка, Ш3Т — ширина 3-го тергита, КИ — кубитальный индекс. Описание линий см. в разделе «Методика».
a, b, c, d, e, f, g, h p < 0,001, i p < 0,01 (достоверность различий между значениями, обозначенным одной и той же буквой).
Результаты двухфакторного иерархического дисперсионного анализа подтвердили наличие достоверных различий по признакам ДХ и Ш3Т как между линиями, так и между семьями в пределах каждой из линий.
Формирование кластеров по линейному принципу наблюдалось для трех из пяти исследованных линий (рис. 1). Только семьи линии I распре- делялись внутри одного обособленного 1-го кластера, однако они характеризовались наибольшей изменчивостью по показателю ДХ (CvДХ = 1,11 %) по сравнению с другими линиями (CvДХ = 0,69-0,86 %) при средней гетерогенности по Ш3Т (CvШ3Т = 2,49 %). Семьи линий III и IV формировали два консолидированных кластера (соответственно CvДХ 0,74 и 0,69 % и CvШ3Т 2,15 и 2,33 %), которые частично перекрывались между собой. У пчел линий II и V сильно варьировали оба признака (CvДХ — 0,79 и 0,86 %, CvШ3Т — 2,66 и 2,52 %). Достоверные различия (p < 0,05) между семьями внутри линий наблюдались по признаку ДХ в линии II (Mst = 0,1359) и признаку Ш3Т в линиях II (Mst = 0,2869) и V (Mst = 0,1525). Межлинейные раз- личия были достоверны (p < 0,05) для показателей ДХ (Mst = 0,1453) и Ш3Т
(Mst = 0,1097), в то время как по КИ достоверных различий мы не выявили.
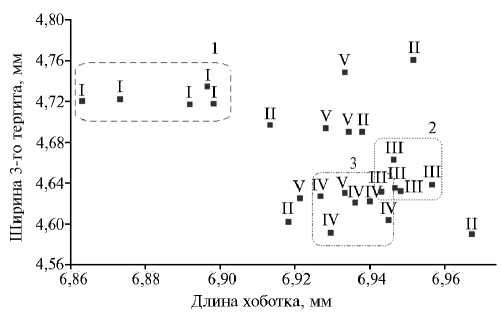
Рис. 1. Распределение 25 семей медоносной пчелы серой горной кавказской породы ( Apis mellifera caucasica L.) в пространстве двух измерений по длине хоботка и ширине 3-го тергита: I-V — изучаемые линии медоносной пчелы; 1-3 — формирующиеся кластеры (Краснодарский край, 2014). Описание линий см. в разделе «Методика».
В целом следует отметить большую гетерогенность в отношении изучаемых признаков у пчел линий II и V и, напротив, большую консолидацию у пчел линий III и IV. Пчелы линии I существенно отличались от остальных по обоим показателям, однако характеризовались значительными межсемейны- ми различиями по ДХ.
Анализ МС профилей выявил сходные тенденции в определении степени внутрисемейной и межсемейной изменчивости (табл. 2).
В линии I (единственной из исследованных) наблюдался избыток гетерозигот (отрицательное значение Fis), что можно рассматривать как
2. Степень внутрисемейной и межсемейной (внутри линий) генетической изменчивости у медоносной пчелы серой горной кавказской породы ( Apis mellifera caucasica L.) по семи локусам микросателлитов (Краснодарский край, 2014)
сравнению с выборкой в целом и более низкая по отношению к линии I межсемейная изменчивость. Наименьшие межсемейные различия, оцененные по показателям Fst и Rst (AMOVA), выявили в линиях III и IV.
Результаты PCA-анализа показали наибольшее сходство семей внутри линии IV, в то время как линии I и II характеризуются наибольшими межсемейными различиями (рис. 2).
|
□ □ |
• & де О А О А |
|
□ о + о □ |
+ • + • + |
Координата 1
Рис. 2. Распределение центроидов семей медоносной пчелы серой горной кавказской породы ( Apis mellifera caucasica L.) в пространстве первых двух главных координат, рассчитанных на основе матрицы Fst при парном сравнении для семи локусов микросателлитов: — линия I, □ — линия II, + — линия III, А — линия IV, • — линия V (Краснодарский край, 2014). Описание линий см. в разделе «Методика». Расчеты проводили для выборки в целом с использованием семьи в качестве субпопуляции (число субпопуляций = 25).
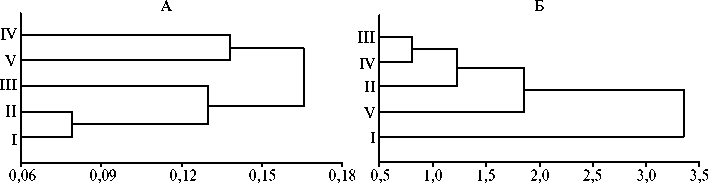
Анализ дифференциации изучаемых линий выявил различия в структуре генеалогического дерева, построенного для семи локусов МС и трех морфометрических признаков (рис. 3). Дендрограмма, построенная по МС, отражала географическую удаленность изучаемых линий (см. рис. 3, А). Формирование двух главных кластеров могло быть следствием географической изолированности линий I, II и III, расположенных в окрестностях Красной Поляны (первая пасека находится на южном склоне хребта Ачи- шхо высотой 2391 м, вторая и третья расположены в ущелье реки Мзымта), от линии IV (пасека расположена на расстоянии 15-25 км от трех перечисленных и отделена от них хребтом Алек высотой 1000 м) и линии V (находится в верховьях реки Псоу, удалена от первой, второй и третьей пасек на 10-16 км и отделена от них хребтом Аибга высотой 2460 м). Существенные генетически различия между линиями IV и V могли быть обусловлены их географической изолированностью. Пасеки, на которых разводят эти линии, расположены на расстоянии около 30 км друг от друга и отделены хребтом Алек, который протянулся на 20 км между реками Мзымта и Сочи на расстоянии 15 км от берега Черного моря, и хребтом Аибга, протянувшимся с северо-запада на юго-восток на 23 км и разделяющим среднюю часть долины Мзымты и верховья Псоу.
Попарные генетические дистанции Евклидова дистанция
Рис. 3. Дендрограмма подобия пяти линий медоносной пчелы серой горной кавказской породы ( Apis mellifera caucasica L.), рассчитанная для семи локусов микросателлитов (А) и трех морфометрических признаков (Б) (Краснодарский край, 2014). Попарные генетические дистанции рассчитаны по M. Nei (32). В обоих случаях применен метод кластеризации UPGMA. Описание линий см. в разделе «Методика».
Структура генеалогического дерева, построенного по морфометри- ческим признакам, не отражала ни географическую общность (различия) происхождения, ни сходство (различия) в хозяйственно полезных признаках изучаемых линий.
Таким образом, нами обнаружены сходные тенденции при оценке внутрисемейной и внутрилинейной (между семьями) изменчивости на основе морфометрических показателей и микросателлитов. Однако данные о степени дифференциации и характере генеалогических связей между линиями, полученные с использованием двух систем анализа, существенно различались. Возможно, подобные расхождения обусловлены тем, что большинство различий по традиционным экстерьерным признакам (морфологические промеры) укладываются в пределы сезонных колебаний и малопригодны для идентификации локальных популяции. Кроме того, они в большей степени подвержены давлению отбора факторов внешней среды, не связанных с воздействием направленного искусственного отбора. Микросателлиты в большинстве случаев селекционно нейтральны и отражают более отдаленные эволюционные события. Следовательно, для получения наиболее полной информации о состоянии и динамике популяций необходимо учитывать результаты, полученные с использованием обоих методов.
Л И Т Е Р А Т У Р А
Sel’skokhozyaistvennaya biologiya [ Agricultural Biology ], 2015, V. 50, ¹ 6, pp. 776-784
MORPHOMETRIC AND MOLECULAR GENETIC DIFFERENTIATION OF Apis mellifera caucasica L. HONEY BEE LINES REARED IN SOCHI REGION
M.S. Fornara1, A.S. Kramarenko1, 2, S.V. Svistunov3, E.M. Lyubimov3,S.S. Sokol’skii3, N.A. Zinovieva1
Acknowledgements:
The equipment of Bioresources and Bioengineering Center of L.K. Ernst All-Russian Research Institute of Animal Husbandry was used.
Supported by the Russian Science Foundation, project 14-36-00039
Список литературы Морфометрическая и молекулярно-генетическая дифференциация линий и семей медоносной пчелы Apis mellifera caucasica L., разводимых в районе Большого Сочи
- Брандорф А.З. Популяционно-генетическая дифференциация медоносных пчел Кировской области. Пчеловодство, 2012, 7: 14-16.
- Ruttner F. Biogeography and taxonomy of honey bees. Germany, Berlin, Springer, 1988.
- Билаш Г.Д., Кривцов Н.И. Селекция серых горных кавказских пчел. Пчеловодство, 1984, 2: 12-13.
- Алпатов В.В. Породы медоносной пчелы. М., 1948.
- Сокольский С.С. Селекционно-разведенческая работа Краснополянской опытной станции пчеловодства. Мат. Межд. науч. конф. «Современные направлении научно-технического прогресса в пчеловодстве». Рыбное, 2007: 80-82.
- Сокольский С.С., Савушкина Л.H. Сохранение генофонда серых горных кавказских пчел. Пчеловодство, 2006, 2: 17-18.
- Ruttner F., Tassencourt L., Louveaux J. Biometrical-statistical analysis of the geographic variability of Apis mellifera L. Apidologie, 1978, 9: 363-381 ( ) DOI: 10.1051/apido:19780408
- Руттнер Ф. Техника разведения и селекционный отбор пчел. М., 2006.
- Oleksa A., Tofilski A. Wing geometric morphometrics and microsatellite analysis provide similar discrimination of honey bee subspecies. Apidologie, 2015, 46: 49-60 ( ) DOI: 10.1007/s13592-014-0300-7
- Rahimi A., Mirmoayedi A., Kahrizi D., Abdolshahi R., Kazemi E., Yari K. Microsatellite genetic diversity of Apis mellifera meda skorikov. Mol. Biol. Rep., 2014, 41(12): 7755-7761 ( ) DOI: 10.1007/s11033-014-3667-7
- Ivanova E., Bienkowska M., Panasiuk B., Wilde J., Staykova T., Stoyanov I. Allozyme variability in populations of A. mellifera mellifera (Linnaeus 1758.), A. m. carnica (Pollman, 1879) and A. m. caucasica (Gorbachev, 1916) from Poland. Acta Zool. Bulg. Suppl., 2012, 4: 81-88.
- Калашников А.Е., Масленников И.В., Колбина Л.М., Удина И.Г. Генетическая дифференциация популяций медоносной пчелы (Apis mellifera L.) и распространение РНК-содержащих вирусов пчел на фоне эпизоотии клеща Varroa destructor на территории Удмуртии. Сельскохозяйственная биология, 2013, 4: 88-92 ( , 10.15389/agrobiology.2013.4.88eng) DOI: 10.15389/agrobiology.2013.4.88rus
- Ilyasov R.A., Kutuev I.A., Petukhov A.V., Poskryakov A.V., Nikolenko A.G. Phylogenetic relationships of dark European honeybees Apis mellifera mellifera L. from the Russian Ural and West European populations. J. Apic. Sci., 2011, 55: 67-76.
- Gruber K., Schöning C., Otte M., Kinuthia W., Hasselmann M. Distinct subspecies or phenotypic plasticity? Genetic and morphological differentiation of mountain honey bees in East Africa. Ecol. Evol., 2013, 3(10): 3204-3218 ( ) DOI: 10.1002/ece3.711
- Whitfield C.W., Behura S.K., Berlocher S.H., Clark A.G., Johnston J.S., Sheppard W.S., Smith D.R., Suarez A.V., Weaver D., Tsutsui N.D. Thrice out of Africa: Ancient and recent expansions of the honey bee Apis mellifera. Science, 2006, 314: 642-645 ( ) DOI: 10.1126/science.1132772
- Harpur B.A., Kenta C.F., Molodtsova D., Lebonb J.M.D., Alqarnic A.S., Owayssc A.A., Zayed A. Population genomics of the honey bee reveals strong signatures of positive selection on worker traits. PNAS USA, 2013, 111(7): 2614-2619 ( ) DOI: 10.1073/pnas.1315506111
- Jensen A.B., Palmer K.A., Boomsma J.J., Pedersen B.V. Varying degrees of Apis mellifera ligustica introgression in protected populations of the black honeybee, Apis mellifera mellifera, in northwest Europe. Mol. Ecol., 2005, 14: 93-106 ( ) DOI: 10.1111/j.1365-294X.2004.02399.x
- Bodur C., Kence M., Kence A. Genetic structure of honeybee, Apis mellifera L. (Hymenoptera: Apidae) populations of Turkey inferred from microsatellite analysis. J. Apic. Res., 2007, 46: 50-56.
- Кривцов Н.И., Горячева И.И., Удина И.Г., Бородачев А.В., Монахова М.А. Идентификация пород и популяций медоносной пчелы с использованием метода ПЦР. Сельскохозяйственная биология, 2010, 6: 26-29.
- Зиновьева Н.А., Кривцов Н.И., Форнара М.С., Гладырь Е.А., Бородачев А.В., Лебедев В.И. Микросателлиты как инструмент для оценки динамики аллелофонда при создании приокского типа среднерусской породы медоносной пчелы Apis mellifera. Сельскохозяйственная биология, 2011, 6: 75-79.
- Кривцов Н.И., Гладырь Е.А., Волкова В.В., Форнара М.С., Лебедев В.И., Зиновьева Н.А. Характеристика аллелофонда трех пород медоносной пчелы России с использованием микросателлитов. Проблемы биологии продуктивных животных, 2011, 1: 41-45.
- Островерхова Н.В., Конусова О.Л., Кучер А.Н., Погорелов Ю.Л., Белых Е.А., Воротов А.А. Популяционно-генетическая структура медоносной пчелы (Apis mellifera L.) в районе д. Леботер Чаинского района Томской области // Вестник Томского государственного университета. Биология, 2013, 1(21). С. 161-172.
- Garnery L., Franck P., Baudry E., Vautrin D., Cornuet J.M., Solignac M. Genetic diversity of the west European honey bee (Apis mellifera mellifera and A. m. iberica). II. Microsatellite loci. Genet. Sel. Evol., 1998, 30: 49-74 ( ) DOI: 10.1186/1297-9686-30-S1-S49
- Pritchard J.K., Stephens M., Donnelly P. Inference of population structure using multilocus genotype data. Genetics, 2000, 155: 945-959.
- Oleksa A., Chybicki I.J., Tofilski A., Burczyk J. Nuclear and mitochondrial patterns of introgression into native dark bees (Apis mellifera mellifera) in Poland. J. Apic. Res., 2011, 50: 116-129 ( ) DOI: 10.3896/IBRA.1.50.2.03
- Alburaki M., Bertrand B., Legout H., Moulin S., Alburaki A., Sheppard W.S., Garnery L. A fifth major genetic group among honeybees revealed in Syria. BMC Genetics, 2013, 14: 117 ( ) DOI: 10.1186/1471-2156-14-117
- Muñoz I., Dall'Olio R., Lodesani M., De La Rúa P. Population genetic structure of coastal Croatian honeybees (Apis mellifera carnica). Apidologie, 2009, 40: 617-626 ( ) DOI: 10.1051/apido/2009041
- Зиновьева Н.А., Попов А.Н., Эрнст Л.К., Марзанов Н.С., Бочкарев В.В., Стрекозов Н.И., Брем Г. Методические рекомендации по использованию метода полимеразной цепной реакции в животноводстве. Дубровицы, 1998.
- Халафян А.А. STATISTICA 6. Статистический анализ данных. М., 2007.
- Крамаренко С.С. Особенности внутри-и межпопуляционной структуры конхиометрической изменчивости наземного моллюска Brephulopsis cylindrica (Gastropoda, Pulmonata, Buliminidae). Вестник зоологии, 2009, 43(1): 51-58.
- Weir B.S., Cockerham C.C. Estimating F-statistics for the analysis of population structure. Evolution, 1984, 38: 1358-1370 ( ) DOI: 10.2307/2408641
- Nei M., Koehn R. Genetic polymorphism and the role of mutation in evolution. In: Evolution Genes and Proteins. Sinauer, Sunderland, MA, 1983: 165-190.
- Slatkin M. A measure of population subdivision based on microsatellite allele frequencies. Genetics, 1995, 139: 457-462.
- Peakall R., Smouse P.E. GenAlEx 6.5: genetic analysis in Excel: population genetic software for teaching and research -an update. Bioinformatics, 2012, 28: 2537-2539 ( ) DOI: 10.1093/bioinformatics/bts460
