Морфометрический подход в оценке величины площади поверхности таллома многоклеточной водоросли Callithamnion corymbosum (J. E. Smith) Lyngb
 Lyngb")
Автор: Празукин Александр Васильевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
На примере многоклеточной водоросли Callithamnion corymbosum рассматривается схема морфометрии и расчетов, позволяющих вычислить величину площади поверхности (S, см2) на уровне осевых структур (0k) и ветвей (k) разных порядков. Представлены регрессионные уравнения, определяющие связь между площадью поверхности ветвей второго (S2) и третьего (S3) порядка ветвления и длиной осевых структур соответствующего порядка ветвления ( l 02, l 03, см): l ogS2 = -0.702 + 1.105 log l 02; l ogS3 = -0.728 + 1.564 log l 03.
Каллитамнион, площадь поверхности, удельная поверхность, архитектоника слоевища, морфометрия слоевищ, иерархия, многоклеточные водоросли, таллом, черное море
Короткий адрес: https://sciup.org/146116670
IDR: 146116670 | УДК: 582.26:
Morphometric approach in defining the surface area of the thallus in multicellular alga Callithamnion corymbosum (J. E. Smith) Lyngb
The multicellular alga Callithamnion corymbosum was chosen as a model for the estimation of its surface area. Here we present the equations to calculate the surface area (S2, cm2) at the axis (0k) as well as at different levels of the branching (k). We provide regression equations, defining the relation between surface areas of the second- (S2) and third-order (S3) branches to the length of the axial structures of the corresponding orders of the branching ( l 02, l 03, см): l ogS2 = -0.702 + 1.105 log l 02; l ogS3 = -0.728 + 1.564 log l 03.
Текст научной статьи Морфометрический подход в оценке величины площади поверхности таллома многоклеточной водоросли Callithamnion corymbosum (J. E. Smith) Lyngb
Введение. В физиологических и экологических исследованиях часто проводятся расчеты поточных характеристик по отношению к физиологически активной поверхности растений (Мокроносов, 1981; Хайлов, 1984; Хайлов и др., 1992). Для водных многоклеточных водорослей такой поверхностью является вся поверхность таллома. По отношению к сложно-разветвленным слоевищам водорослей используется поэтапная технология определения площади их поверхности (Фирсов, 1984; Хайлов, Фирсов, 1985; Хайлов и др., 1992; Празукин, 2015), и, в соответствии с особенностями морфологии того или иного объекта, существуют определенные нюансы её исполнения.
Цель настоящей работы: на примере многоклеточной водоросли CaUithamnion corymbosum (J. Е. Smith) Lyngb. продемонстрировать морфометрический подход в оценке величины площади поверхности слоевища и оценить роль каждого класса морфологических структур С. corymbosum в формировании её ассимиляционной поверхности.
Материал и методика. В апреле-мае 2015 г., в период наших исследований, на преимущественно затененных участках горизонтальных и вертикальных стенках скал открытых участков Севастопольского прибрежья до глубин 30-40 см С. corymbosum формировала сплошные заросли (растительный покров).
Для подробной морфометрии слоевищ С. corymbosum отбирали отдельные кустики растений (рис. 1а), формируемые одним вертикальным «побегом» (Ко, рис. 1 б). В структуре вертикального побега отчетливо выделяется главная ось (Коо), которая является основой для всех последующих порядков осевых структур (Koi - Кою , рис. 1 б - г) и ветви разного уровня организации (Ki - Кд, рис. 1 в, г). «Ветвь» рассматривается нами как сложноразветвленный элемент слоевища (иерархически организованный ансамбль осевых структур), обладающий собственной морфофункциональной структурой, закономерно меняющейся во времени (Фирсов, 1984; Хайлов и др., 1992; Празукин 2008, 2015). В таком случае, ветвь первого порядка (Кь рис. 1 в, д) - это ось первого порядка (Koi) со всеми осевыми структурами последующих порядков ветвления (К02 - Кою), аналогично, ветвь второго порядка (К2, рис. 1 в) - это ось второго порядка (К02) со всеми осевыми структурами последующих порядков ветвления (Коз - Кою)- Ветвь четвертого порядка (Кд) состоит из осевых структур Код - Кою, ветвящихся дихотомически (рис. 1 г).
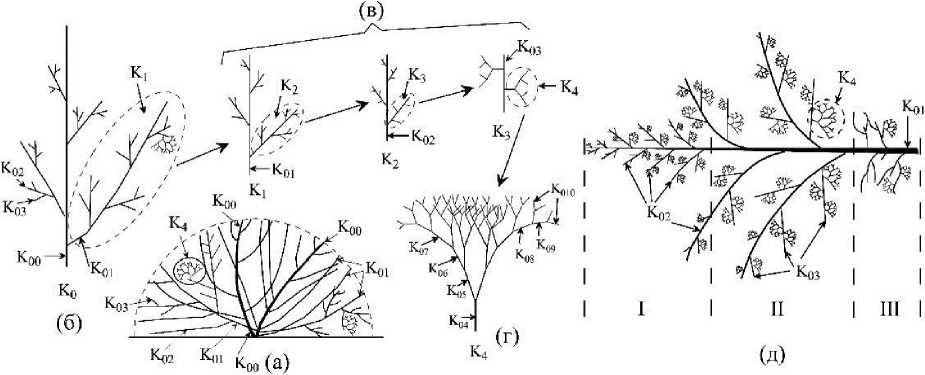
Рис. 1. Различные уровни организации Callithamnion corymbosum; а -куст, образованный двумя вертикальными «побегами; схема осевого аппарата вертикального «побега» (б) и ветвей первого - третьего (в) и четвертого (г) порядков; д - схема осевой структуры ветви первого порядка Коо, К01 - Кою - осевые структуры нулевого («ствол»), первого -десятого порядка ветвления; Ко - вертикальный «побег»; К! - К4 - ветви первого - четвертого порядка; I - III - морфологические отделы ветви первого порядка выделяемые вдоль оси К0]
Особенностью структуры слоевища С. corymbosum является то, что основная масса растения и его фотосинтезирующая поверхность (S) создается за счет относительно небольшого числа крупных ветвей первого порядка (Ki, рис. 1 в, д ), отрастающих от самой нижней части оси Коо. Это позволило нам сосредоточить внимание на морфометрии крупных ветвей первого порядка. Ветви первого порядка расчленяли на ось Koi и ветви второго порядка (К2). Последние, в соответствии с длиной оси К02, делили на три условные группы: «мелкие» (0.2 - 0.6 см), «средние» (>0.6 - 0.8 см) и «крупные» (1.3 — 1.8 см). Крупные ветви К2 были, как правило, немногочисленными и располагались в среднем отделе ветви Ki (рис. 1 д). Далее, из каждой размерной группы ветвей отбирали по три-пять К2 и каждую ветвь расчленяли на ось К02 и ветви К3. Подробно морфометрировали ветви четвертого порядка (К4, рис. 1 г).
4k’ СМ
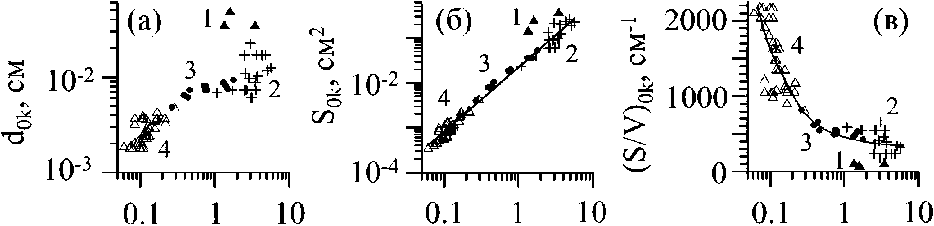
Рис. 2 . Связь между диаметром (dOk), площадью поверхности (80к) и удельной поверхностью ((S/V)ok) осевых структур разного порядка ветвления Callithamnion corymbosum, - с одной стороны, и, соответственно, с их длиной (70к), с другой Осевые структуры нулевого, «ствол» (1), первого (2), второго (3) и третьего (4) порядка ветвления
Во всех случаях с помощью бинокулярной лупы МБС-1 измеряли длину (/ок) и диаметр (dok) осевых структур нулевого, первого - десятого порядка ветвления. Величина диаметра осевых структур рассчитывалась как средняя по результатам нескольких измерений на разных участках структур. Подсчитывали общую численность структур последующего порядка (nok+i) на осевых структурах предыдущего (0k).
Геометрический объем (Vok, см3) и геометрическую поверхность (Sok, см2) осевых структур рассчитывали по формулам, подходящим для аппроксимации их геометрических фигур: цилиндра, усеченного конуса, полусферы и их комбинаций (Выгодский, 1965).
Результаты и обсуждение. Вначале рассмотрим структурную организацию слоевища С. corymbosum, что позволит на следующем этапе рассмотрения представить алгоритм расчета площади поверхности этого вида водорослей.
Слоевище С. corymbosum в своей структуре сочетает моноподиальное (осевые структуры нулевого - четвертого порядка, рис. 1 а, б) и дихотомическое (ветви четвертого порядка, рис. 1 г) типы ветвления. Для осевых структур каждого порядка ветвления характерны собственные диапазоны варьирования значений параметров: длины (Zok), диаметра (dok), объёма (Уок), площади поверхности (Sok) и удельной поверхности (S/V)ok (рис. 2, 3). В ряду порядков, от первого до четвертого, длина, диаметр, площадь поверхности и объем осевых структур быстро снижаются, а удельная поверхность повышается. Особенность такой динамики lok и dok состоит в том, что в ряду порядков длина уменьшается в 130 раз быстрее диаметра. Для цистозиры косматой (Cystoseira crinita (Desf.) Вогу) этот показатель равен четырем (Хайлов и др., 1992), а для бриопсиса перистого (Bryopsis plumosa (Huds.) Ar.) - трем (Празукин, 2007). У С. corymbosum моноподиальное ветвление ограничено четырьмя порядками ветвления, у цистозир - четырьмя, реже - пятью-шестью, у бриопсиса перистого - пятью. Можно предположить, что существуют минимальные размеры структур (lok, dok) и их соотношения, при которых дальнейшее моноподиальное ветвление становится невозможным, и в случае с С. corymbosum осевые структуры пятого и последующих порядков ветвления ветвятся дихотомически (рис. 1 г).
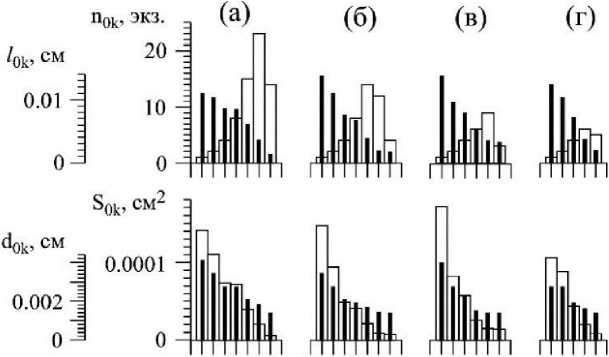
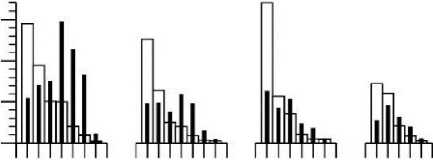
Возможно, и при дихотомическом ветвлении существует ограничение в количестве последовательных генераций осевых структур. В структуре ветвей четвертого порядка С. corymbosum их насчитывается 4-6 (рис. 3). При дихотомическом ветвлении численность осевых структур в каждой последующей генерации удваивается, но это правило нарушается на завершающем акте ветвления, и в этом случае численность структур снижается (рис. 3).
В ряду порядков структур (Ко4 - Кою) происходит одновременно как уменьшение значений длины (/ок), так и диаметра (dok) без существенного изменения их пропорций (рис. 3). Наблюдаемое снижение значений /ок и dok нельзя рассматривать как прямое следствие дихотомического ветвления. Так, например, у церамиума шиповидного (Ceramium echionotum J. Ag.) при аналогичном типе ветвления у осевых структур уменьшение значений длины и диаметра обнаруживается только на завершающих этапах ветвления (Хайлов и др., 1992).
EVOk, см3 VOk, см3
РЮ"7
0.5*10-7
SSOk, см2 (S/V)Ok, см"1 0.0004 2000 4 0.0002 0^ 0
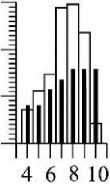
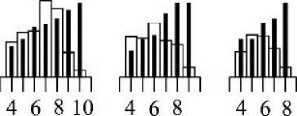
Рис.
порядок ветвления
3 . Средние значения индивидуальных параметров и значения кумулятивных характеристик осевых структур разных порядков ветвления у четырех ветвей четвертого порядка (К4) Callithamnion corymbosum, взятых с ветвей второго порядка разного размерного диапазона: «крупные» (а), «средние» (б, в), «мелкие» (г) (см. табл. 1)
VOk- объём осевой структуры; nok, ZVok, Z^ok, - соответственно, численность, суммарный объём и суммарная площадь поверхности осевых структур каждого порядка ветвления. Обозначение остальных параметров те же, что на рис. 2
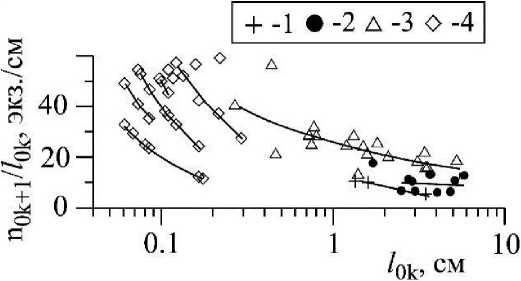
Рис. 4 . Плотность размещения осевых структур последующего порядка на осевых структурах предыдущего порядка (nOk+i//ok) как функция длины осевых структур (Zok) Callithamnion corymbosum
1 - 4 - плотность размещения осевых структур первого - четвертого порядка ветвления на структурах предыдущего порядка
Суммарная поверхность (ZSok) и суммарный объём (ZVok) структур в ряду порядков ветвления ветви четвертого порядка С. corymbosum изменяются по одновершинной зависимости, а их соотношение (S/V)ok увеличивается (рис. 3). По отношению ко всей ветви К4 величина площади поверхности укладывается в диапазоне от 0.00116 до 0.0018 см2, а удельная поверхность - от 1600 до 1800 см"1 (табл. 1, 2).
В табл. 1 и 2 представлены средние значения параметров осевых структур и ветвей первого и четвертого порядка ветвления, что позволяет проследить последовательность «сборки» ветвей, начиная с третьего и заканчивая первым порядком ветвления.
Общий объем (Усл) и общая поверхность (8СЛ) слоевища складываются, соответственно, из объема и поверхности ствола (оси нулевого порядка, Voo, Soo) и объема и поверхности всех ветвей первого порядка (ZVb ZSi):
Усл = Уоо + ЕУь
SM = Soo + ZSb
Таблица!
Средние значения параметров осевых структур и ветвей ________________1-4 порядков (К! - К4)
|
Ветви первого порядка (К,) ’ |
Ветви второго порядка (К2) |
Ветви третьего порядка (Кз) |
К, |
||||||||||
|
Si, см2 |
Soi, см2 |
Размерные группы ветвей К2 по Lq2 , СМ |
Ветви второго порядка (К2) |
s2, см2 |
S02, см2 |
СМ |
поз/ ^02, экз./см |
s3, см2 |
8оз, см2 |
СМ |
noV /оз экз./см |
s4, см2 |
|
|
П |
ES, см2 |
||||||||||||
|
6,271 |
246 Е-3 |
Мелкие (0.2-0.6) |
62 |
373 Е-2 |
602 Е-4 |
83 Е-4 |
0,39 |
393 Е-1 |
338 Е-5 |
70 Е-5 |
97 Е-3 |
412 Е-1 |
7 Е-4 |
|
Средние (>0.6-0.8) |
4 |
76 Е-2 |
1893 Е-4 |
168 Е-4 |
0,76 |
289 Е-1 |
784 Е-5 |
86 Е-5 |
И Е-2 |
542 Е-1 |
12 Е-4 |
||
|
Крупные (1.3-1.8) |
5 |
154 Е-2 |
3070 Е-4 |
222 Е-4 |
1,58 |
216 Е-1 |
833 Е-5 |
113 Е-5 |
15 Е-2 |
263 Е-1 |
18 Е-4 |
||
|
V,, |
Voi, см3 |
Lq2 , СМ |
п |
XV, |
v2. |
Vq2> |
СМ |
Поз/ /о2 |
v3. |
V03, |
СМ |
П04/ /оз |
v4. |
|
5189 Е-6 |
846 Е-6 |
Мелкие (0.2-0.6) |
62 |
28 Е-4 |
451Е-7 |
148Е-7 |
0,39 |
393 Е-1 |
198Е-8 |
447Е-9 |
97 Е-3 |
412 Е-1 |
38 Е-8 |
|
Средние (>0.6-0.8) |
4 |
5 Е-4 |
133Е-6 |
299Е-7 |
0,76 |
289 Е-1 |
470 Е-8 |
553Е-9 |
И Е-2 |
542 Е-1 |
69 Е-8 |
||
|
Крупные! 1.3 1.8) |
5 |
10 Е-4 |
202Е-6 |
250Е-7 |
1,58 |
216 Е-1 |
518Е-8 |
827Е-9 |
15 Е-2 |
263 Е-1 |
И Е-7 |
||
В свою очередь, объем (Vi) и поверхность (Si) ветвей первого порядка складываются, соответственно, из объема и поверхности оси первого порядка (V01, S01) и объема, и поверхности всех ветвей второго порядка (ZV2, £S2):
Vi = Voi+ZV2,(1)
Si = S01 + ZS2, (2)
Последовательно раскрывая уравнения (1) и (2), получаем:
V 1 - Vol + n02 (V02 + Поз (Vo3 + n4JV4)), (3)
Si = Soi + П02 (So2 + Поз (8оз + n4 S4)). (4)
По отношению к ветвям второго порядка можно записать:
V2 = V02 + Поз (V03 + n4 V4), (5)
§2 = §02 + Поз (8оз + n4 S4), (6)
а по отношению к ветвям третьего порядка:
Уз = У0з + п4у4, (7)
S3 = So3 + n4S4. (8)
Таблица2
Средние значения удельной поверхности (S/У) осевых структур и ветвей первого - четвертого порядка
|
Осевые структуры и ветви первого порядка |
Размерные группы ветвей К2 ПО L02 , см |
Осевые структуры второго - третьего порядка |
Ветви второго - четвертого порядка |
||||
|
(S/V)oi, см"1 |
(S/V)b см"1 |
(S/V)02, см"1 |
(8/У)оз, см"1 |
(S/V)2, см"1 |
(S/V)3, см"1 |
(S/V)4, см"1 |
|
|
291 |
1209 |
Мелкие (0.2-0.6) |
562 |
1560 |
1334 |
1709 |
1752 |
|
Средние (>0.6-0.8) |
562 |
1561 |
1421 |
1669 |
1683 |
||
|
Крупные (1.3-1.8) |
891 |
1365 |
1519 |
1608 |
1654 |
||
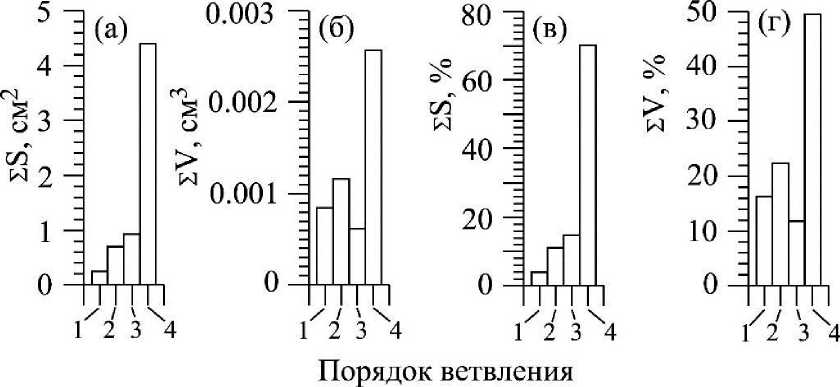
Рис. 5 . Суммарная площадь поверхности (а) и суммарный объем (б) осевых структур (1 - 3 - KOi - К03) и ветвей четвертого порядка (4 - К4) в структуре ветви первого порядка (КО Callithamnion corymbosum и вклад названных структур в общую поверхность (в) и объем (г) ветви К,
По сути, мы имеем дело с процессом формирования ветвей и, в целом, всего слоевища. Его можно представить как результат иерархического «обрастания»: на растущей осевой структуре нулевого порядка («стволе») появляются дочерние оси-эпифиты (Koi), а на них, в свою очередь, - дочерние оси (Ко2) и т. д. (рис. 1). В этом случае, при расчетах общей поверхности ветви или всего слоевища становится важной такая характеристика как плотность размещения осевых структур последующего порядка на осевых структурах предыдущего (nOk+i//ok). Как следует из рис. 4, эта характеристика является функцией длины осевых структур предыдущего порядка ветвления (/ок), и названная связь хорошо обнаруживается по отношению к осевым структурам третьего и четвертого порядка ветвления. Плотность размещения К04 на структурах третьего порядка во многом зависит и от индивидуальных размеров структур второго порядка (/02, табл. 1), что проявляется в широком диапазоне варьирования значений (11.4 -59.2 экз. см'1) этого показателя (noV/оз, рис. 4). В целом, в ряду порядков от нулевого до третьего плотность размещения осевых структур увеличивается с 5.5 экз. см"1 на осевых структурах нулевого порядка до 59.2 экз. см"1 на осевых структурах третьего порядка.
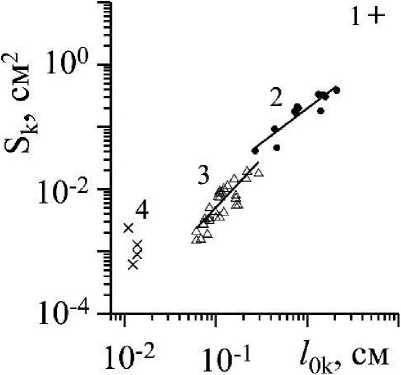
Рис. 6 . Зависимость площади поверхности (Sk) ветвей первого -четвертого порядка ветвления от длины осевых структур соответствующего порядка ветвления (ZOk) 1 - 4 - ветви первого - четвертого порядка ветвления
В композиционной структуре ветви первого порядка (Ki, рис. 1 д) С. corymbosum выделяются две группы морфологических объектов: собственно осевые структуры первого - третьего порядка ветвления (Koi - Коз, рис. 1 д ) и ветви четвертого порядка (Кд, рис. 1 д). Вторые, по сравнению с первыми, отличаются относительно небольшими значениями индивидуальных характеристик (/оь Vok, Sok), но, в то же время, они превосходят осевые структуры первого - третьего порядка по величине удельной поверхности (S/V)ok, рис. 2 в, табл. 2) и по численности их в структуре ветви Ki, а значит и по суммарной площади поверхности (£S). За счет ветвей четвертого порядка формируется 70% площади поверхности ветви первого порядка (рис. 5). Учитывая и тот факт, что ветви четвертого порядка обладают наиболее активной фотосинтезирующей поверхностью (Празукин, 2015), по сравнению со структурами предыдущих порядков ветвления (Koi - КОД то можно заключить, что эти структуры выполняют основную роль в формировании общего потока карбонатного углерода ветви Ki и всего растения в целом. Аналогичная картина наблюдается и в структуре ветвей Ki В. plumosa. Поверхность ветви Ki этого вида на 72.6 % складывается из поверхностей «коротких» (мелкоразмерных) осевых структур, доля которых в общей массе ветви составляет 53 % (Празукин, 2007).
Теперь обратимся непосредственно к вопросу об определении величины площади поверхности слоевища С. Corymbosum и его морфологических структур.
По результатам измерений морфологических параметров ветвей К2 и Кз (табл. 1), взятых с одной и той же ветви Ki С. Corymbosum, была проведена численная «сборка» вышеназванных объектов, рассчитана их площадь поверхности (Sk, см2) и построены зависимости (Sk = f(/ok), рис. 6), описываемые следующими уравнениями регрессии:
LogS2 = -0.702 + 1.105 log/02 (R2 = 0.84, n = 12), (9)
LogS3 = -0.728 + 1.564 log/03 (R2 = 0.68, n = 35). (10)
Таким образом, площади поверхностей ветвей второго и третьего порядка можно легко определить, рассчитав её по регрессионным уравнениям: (9) и (10). Для определения площади поверхности ветвей первого порядка потребуются дополнительные расчеты, учитывающие особенности её структуры (рис. 1 д).
Согласно уравнению (2) площадь поверхности ветви первого порядка (Si) равна сумме площадей поверхностей оси первого порядка (Soi) и всех ветвей второго порядка (£82). Для установления величины Soi достаточно измерить длину (Zoi) и диаметр (doi) осевой структуры и рассчитать Soi по формуле усеченного конуса (Выгодский, 1965). Для расчета величины £S2 вначале необходимо по уравнению (9) рассчитать величину 82 каждой ветви К2, а затем полученные значения суммировать.
Определение величины площади поверхности ветви К2 можно осуществлять и в результате ее индивидуальной «сборки». 82 - как сумма площадей поверхностей оси второго порядка (S02) и всех ветвей третьего порядка (£83): S2 = S02 + £83. В таком случае, площадь поверхности каждой ветви третьего порядка (S3) рассчитывается по уравнению (10).
Заключение. Можно сказать, что для установления величины площади поверхности ветвей первого - третьего порядка С. corymbosum достаточно измерить такие легко определяемые морфологические параметры, как длину (Zok) и диаметр (dok) осевых структур соответствующего порядка ветвления и воспользоваться регрессионными уравнениями (9) и (10).
Празукин А.В. Морфометрический подход в оценке величины площади поверхности таллома многоклеточной водоросли CaUithamnion corymbosum (J. Е. Smith) Lyngb / А.В. Празукин // Вести. ТвГУ. Сер.: Биология и экология. 2016. № 4. С. 195-205.
Список литературы Морфометрический подход в оценке величины площади поверхности таллома многоклеточной водоросли Callithamnion corymbosum (J. E. Smith) Lyngb
- Выгодский М.Я. 1965. Справочник по элементарной математике. М.: Наука. 424 с.
- Мокроносов А.Т. 1981. Онтогенетический аспект фотосинтеза. М.: Наука. 195 с.
- Празукин А.В. 2007. Различные приемы описания фитосистем разного уровня организации на примере зеленой водоросли бриопсис (Bryopsis Lamouroux) Черного моря//Морской экологический журнал. Т. 6. № 1. C. 66-87.
- Празукин А.В. 2008. Морфофункциональные образы растения (на примере многоклеточных водорослей)//Вестн. ТвГУ. Сер. Биология и экология. Вып. 9. С. 206-218.
- Празукин А.В. 2015. Экологическая фитосистемология. М.: Перо. 375 с.
- Фирсов Ю.К. 1984. Иерархический структурно-функциональный анализ морской многоклеточной водоросли Cystoseira barbata: автореф. дис.... канд. биол. наук. Севастополь. 20 с.
- Хайлов К.М. 1984. Два способа выражения интенсивности фотосинтеза у морских макрофитов в связи с их функциональной морфологией//Биология моря. № 6. С. 36-39.
- Хайлов К.М., Празукин А.В., Ковардаков С.А., Рыгалов В.Е. 1992. Функциональная морфология морских многоклеточных водорослей. Киев: Наукова думка. 280 с.
- Хайлов К.М., Фирсов Ю.К. 1985. Расчет величины ассимиляционной поверхности Sargassum на разных уровнях организации//Ботанический журнал Т. 70. № 2. С. 215-221.
