Morphological response and yield of rice cultivars to water deficit condition at different growth stages on sandy loam soil in Ibadan

Author: Akpoilih O.A., Dada O.A.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 2 т.21, 2025.
Free access
Drought is a major abiotic constraint in upland rice field, causing severe yield loss of more than 50%. Southwestern Nigeria, which covers almost 30% upland rice fields are purely rainfed and rainfed farming is no longer reliable due to fluctuation in rainfall pattern and volume causing drought in rice field. The need to understand response of upland rice cultivars to moisture deficit at different phenological stages is important to device appropriate drought management strategy in upland rice field. Information on response of upland rice cultivar to water deficit condition at different growth stages is scanty. Therefore, response of 12 Rice Cultivars (RC) to water deficit condition at tillering, panicle initiation and grain filling on sandy loam was investigated. The study was a 4×12 factorial, arranged in completely randomized design with six replicates was conducted in a screenhouse during the early and late dry seasons of 2018 in Ibadan. Treatments included twelve RC: OFADA, IGBEMO, FARO-16, FARO-44, FARO-60, IR-64, APO, NERICA-4, NERICA-5, NERICA-7, NERICA-8, water stressed at tillering, Panicle Initiation (PI) and Grain Filling (GF) and Well-Watered Soil (WWS, served as control). Three weeks old rice seedling was transplanted into pot containing 5 kg soil. Data were collected on plant height (cm), leaf area (cm2), number of leaves and tillers, leaf roll and dry, dry matter and yield components. Data were analyzed using analysis of variance (ANOVA) and means separated with LSD (p function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Drought, yield components, upland rice cultivars, growth and biomass yield
Short address: https://sciup.org/143184716
IDR: 143184716
Text of the scientific article Morphological response and yield of rice cultivars to water deficit condition at different growth stages on sandy loam soil in Ibadan
Globally rice is an important staple food crop for more than 3 billion people in the world (Khush, 2005 and Nguyen, 2010). Rice forms a major part of the Nigerian diet and most populations in developing countries ( akariyawo et al., 2013). Rice has been reported to generate more income for arable farmers than any other crop (FAO, 2017).
Harfold, (2011) reported that rice consumption in Nigeria is higher than that of any other staple crops due to increase in population, rapid urbanization and changing eating habits. Nigeria relied on importation to meet the increasing rice demand. Rice importation profile of Nigeria stands at approximately 3 MT annually, whereas local production stands at 2 MT (FAO, 2017). imilarly, Uduma et al . (2016) noted that the inability of local supply to meet up with rice demand has given rise to the high importation of rice in Nigeria.
tudies have shown that biotic and abiotic stresses are the major constraints affecting rice production (Ismaila et al., 2010). The abiotic factors including flooding, low soil fertility, iron toxicity and particularly drought have been reported to reduce global rice production by 50% (Nwilene et al., 2008; Mostajeran and Rashimi-Eichi, 2009; Pawar et al., 2017).Water deficit is an abiotic factor affecting arable crops particularly, rice production in Nigeria (Dada et al., 2018). It causes stomata closure ( antos and Carlesso, 1998), and other metabolic processes that limit normal growth and development (Amin et al ., 2009; Jalal et al ., 2012). ubstantial reduction in plant growth and yield has been reported to occur because of water deficit (Jalal et al ., 2012; Hayatu et al ., 2014).
Rice crop have been reported to be susceptible to drought stress which causes yield loss in rice field (Pantuwan et al., 2002). However, some varieties have been reported to be tolerant than others, out-yielding those subjected to the same level of drought (Dada et al., 2018). NERICA- New Rice for Africa has been known for higher yield, tolerant of major stress such as drought; it is higher in protein, with good taste and stable yield under different conditions compared to the traditional rice varieties (Africa Rice Center et al., 2008;
Wopereis et al., 2008; Arouna et al ., 2017).
Plant tolerance to drought is generally complex due to interactions between factors causing damage and various physiological responses within the plant (Ashraf and Hafeez, 2004; Manivannan et al ., 2008). An adequate understanding of rice response to water deficit is required to select the most appropriate varieties tolerant of this biotic limitation in order to increase rice productivity. Timing, duration, severity, and stage of exposure have roles in determining how plants respond to water deficit. Other factors include stage of plant development ( ingh et al ., 2012).
creening for drought tolerant varieties and determining the growth stage(s) at which water deficit is most detrimental to rice plant development is necessary to evolve appropriate drought mitigation approach that could be deployed for improving yield of upland rice. Equally, identification and selection of varieties with high level of tolerance to drought for further breeding programme is expedient for improving upland rice production.
More information is however, needed on morphological response of upland rice cultivars to drought stress at different stages of growth. Therefore, the aim of this study was to investigate the response of twelve upland rice cultivars to water deficit condition at various growth stages in Ibadan, southwestern Nigeria. MATERIALS AND METHODS
Study site
The study was a pot experiment, carried out between 2018-2019 in the screen house of the Department of Crop Protection and Environmental Biology, Faculty of Agriculture, University of Ibadan, Nigeria (7°.45′E; 3°.89′N, 227m above sea level). The ambient temperature of the screen house was 32-40 °C while the relative humidity was 66-85% and the average precipitation during the trial was1260 mm.
Soil collection and analysis
The soil was collected from a field at University of Ibadan Teaching and Research Farm. The field where the soil was collected had been used for cultivation of vegetables such as Solanum spp., Lycopersicum esculentum, Abelmoscuhs esculentus, Zea mays, Vigna unguiculata in the previous three planting seasons. Prior to transplanting, routine soil analysis were performed following standard analytical methods described by IITA (1979).
Sources of rice seeds
Twelve upland rice cultivars were investigated to establish their responses to drought. These included nine commonly cultivated lines: OFADA, IGBEMO, FARO-16, FARO-60, FARO-44, NERICA-4, NERICA-5, NERICA-7, NERICA-8, which were collected from rice farmers during preliminary field survey across southwestern, Nigeria. The remaining three cultivars IR-64, VANDANA and APO were collected from Africa Rice Center (WARDA), Nigeria sub-station, located at the International Institute of Tropical Agriculture (IITA), Ibadan.
Nursery and transplanting
The seeds of the cultivars were raised in nursery beds for two weeks after which they were transplanted to the prepared pots each, filled with 5 kg soil. One vigorous seedling with three fully formed leaves was transplanted per pot.
Treatments and experimental design
There were 48 treatments derived from combination of twelve upland rice cultivars and phenology at which water stress was imposed: tillering, panicle initiation and grain filling while, treatment without imposition of water stress served as the control. The treatments were arranged in completely randomized design and replicated six times. A total of 288 pots used for the experiment were arranged in the screen house at the crop garden of the Department of Crop Protection and Environmental Biology. A repeat trial was carried out following same procedures employed in the first trial. All cultural practices were applied as appropriate.
Imposition of water stress
Prior to imposition of water stress, transplanted seedlings were adequately nurtured for two weeks to ensure that they were well established and had uniform development. Drought was imposed at vegetative stage (active tillering) for 14 days only, while drought was imposed for ten days each at panicle initiation and grain filling stages.
Water stress treatment was imposed twice at tillering at seven days interval. In this treatment, irrigation was withheld for seven consecutive days till the 8th day when plants were irrigated to field capacity with 600 mL water representing100% of water required to attain field capacity. Field capacity was determined following the method of axton and Rawls (2006). At the 9th day, water stress treatment resumed for another seven days. Other plant sets, not subjected to water stress continued to receive water when soil moisture was low with 300 mL water representing 50% of water required to attain field capacity. Drought was imposed at panicle initiation and grain filling stages, each, for ten days at five days intervals. Whenever the stressed plants were to be unstressed after drought treatments, they were irrigated with 600 mL and subsequently with 300 mL based on soil moisture level test carried out, usually every other day till harvest. The moisture level was monitored daily at 9:00 hour and 15:00 hour by probing the moisture sensor at the center of the pots at 8-9 cm depth. Data Logger Em50 series was used to monitor the soil moisture level.
Data collection
Data were collected after the plants had been water stressed for a specific period of time based on the treatment. Morphological parameters such as plant height (cm), numbers of tillers, number of leaves, leaf area and number of tillers formed were assessed at tillering, panicle initiation and grain filling growth stages. Leaf area was evaluated following the method described by Gomez (1972). Dry matter yield was determined at the final harvest, each plant was uprooted carefully and washed thoroughly under running water to remove soil particles to determine dry matter yield at the final harvest. The samples were partitioned into root and shoot, kept in paper bags and oven dried at 80 °C until constant weight was attained after 48 hours. The dried samples were weighed using a measuring scale (Camry 202)
Soil moisture content determination
This was determined using the soil moisture meter (LM -714) which was done by probing the meter into the soil before and after irrigation.
Assessment of leaf response to water stress
Indicators of water stress such as leaf rolling and leaf drying were assessed using standard procedures described by International Rice Research Institute (IRRI, 2016).
Leaf rolling
The score ranged from 1-9. Where 1 = light folding, 3 = Half rolling, 5 = Full to tight rolling, 7 = Tight rolling, 9 = Tube like rolling (no unrolling).
Dried tip
The scores ranged from 1-9. Where 1 = No drying, 3 = Few tips drying (5-10%) basal leaves drying, 5 = 5% leaf tip drying extending to ¼ of leaf blade, 5-10% of lower leaves dried, 7 = Most upper leaves ½ – ¾ are dried; 60-80% lower leaves are dried, 9 = 100% of all leaves dried, plants generally approach permanent wilting point, some die. The various scores were logarithm transformed before they were analyzed statistically.
Yield and yield components such as numbers of panicles and spikelets, weight of panicles, filled grains, unfilled grains, 100-seed weight and grain yield (g)/plant, were determined following procedures of IRRI (2016).
Data Analysis
Data were analyzed using analysis of variance (ANOVA) of tatistical Analysis ystem ( A , 2000) and different means were separated by Least ignificant Difference (L D) at p≤0.05.
RESULTS
Physical-chemical properties of the soil
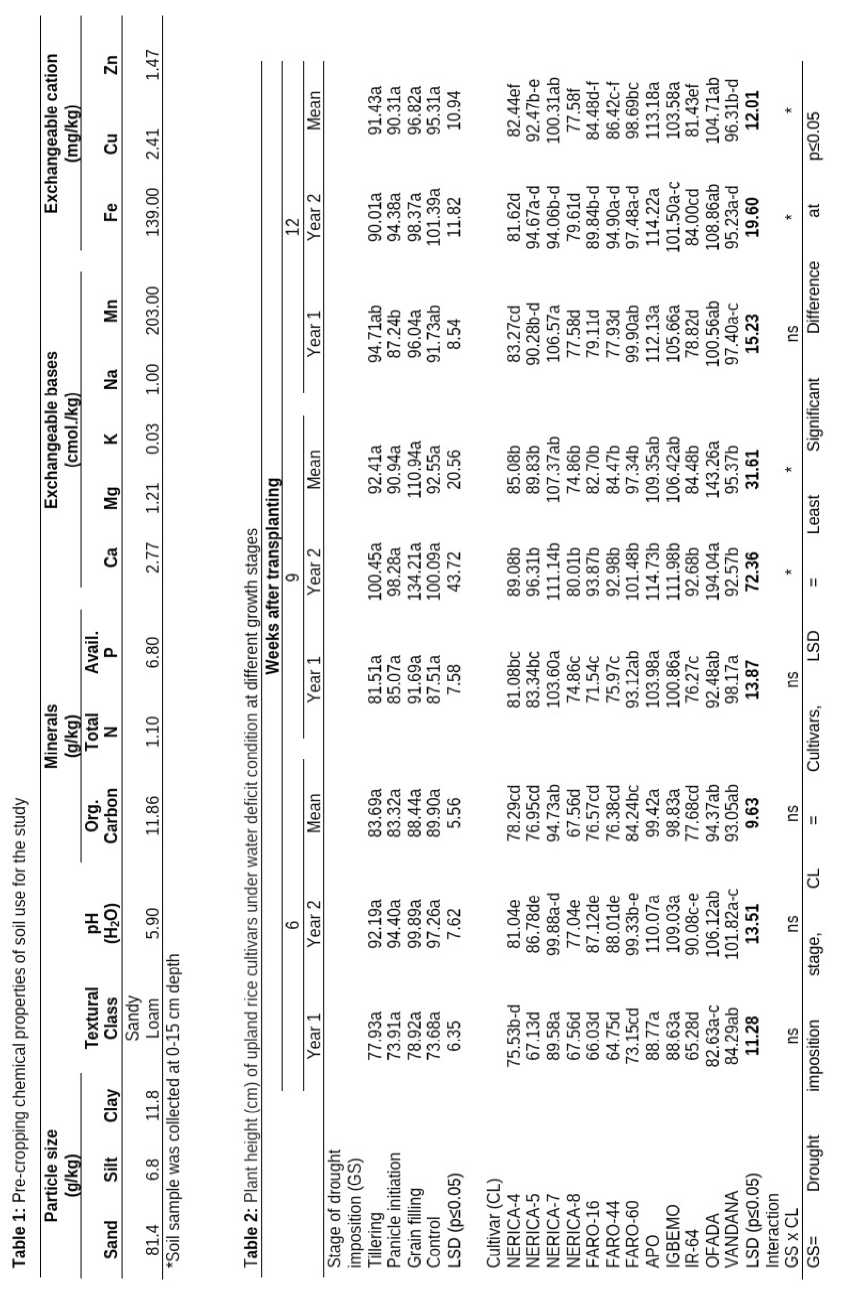
The result of physical and chemical analysis of the soil for the study is presented in Table 1. The textural class of the soil is sandy loam and the pH revealed that the soil was slightly acidic (5.9).The soil had low organic carbon, total nitrogen and potassium but had high phosphorus content. Other exchangeable bases and cations were within the optimum mineral range for upland rice growth. The manganese and iron threshold were extremely above the adequate range.
Effect of water deficit condition at different growth stages on height of rice cultivars
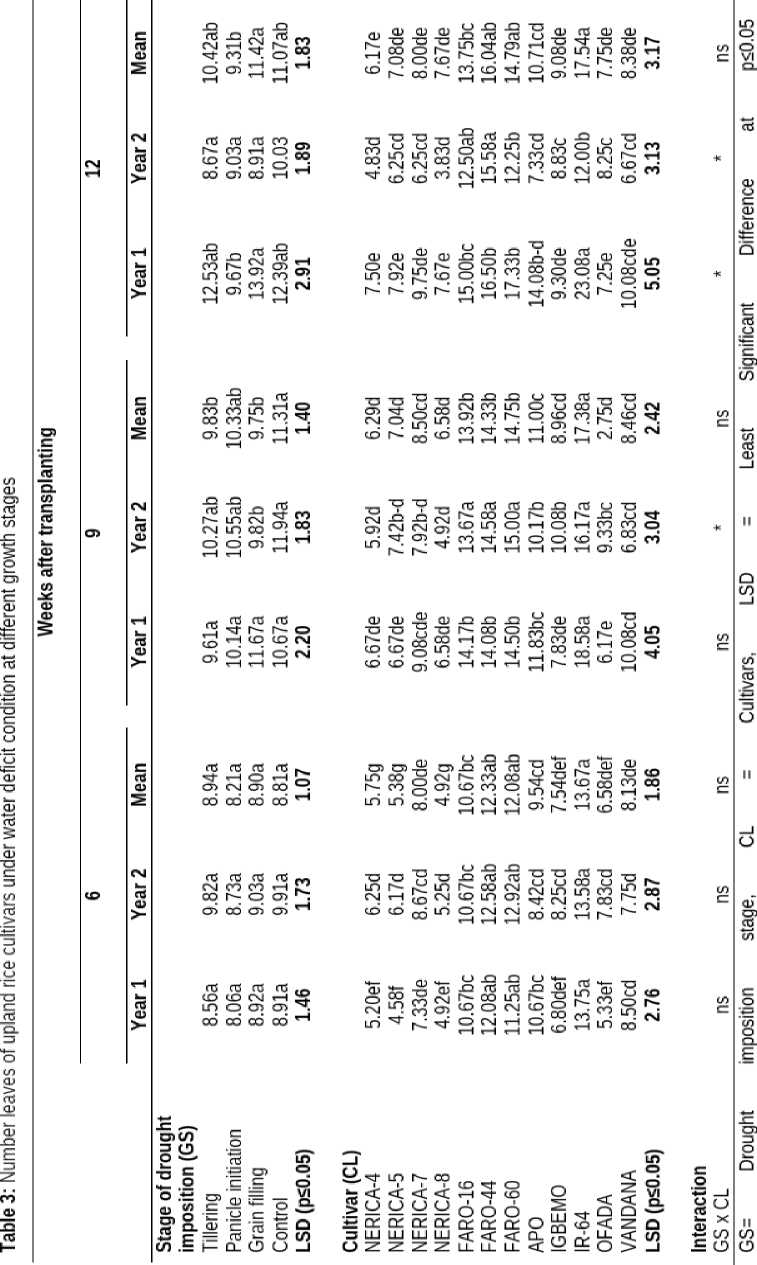
Generally, imposition of drought at all the growth stages had no significant effect on the height of the plants; however the plants were taller at grain filling than any other growth stage. The height of the different rice cultivars differed significantly in response to water limiting conditions. Among the twelve cultivars, APO had significantly taller plant (99.42 and 113.18 cm) but this was not significantly (p≤0.05) taller than IGBEMO, OFADA and NERICA-7 all through the growing periods except at 9 weeks after transplanting (WAT) when OFADA had the tallest height (143.26 cm) as shown in Table 2.
The interaction between the two factors showed that water deficit condition had no significant influence on height of different rice cultivars at 6 WAT, however, stage of drought occurrence had significant influence on the height of upland rice cultivars at 9 and 12 WAT (Table 2).
Effect of water deficit condition at different growth stages on number of leaves formed by upland rice cultivars
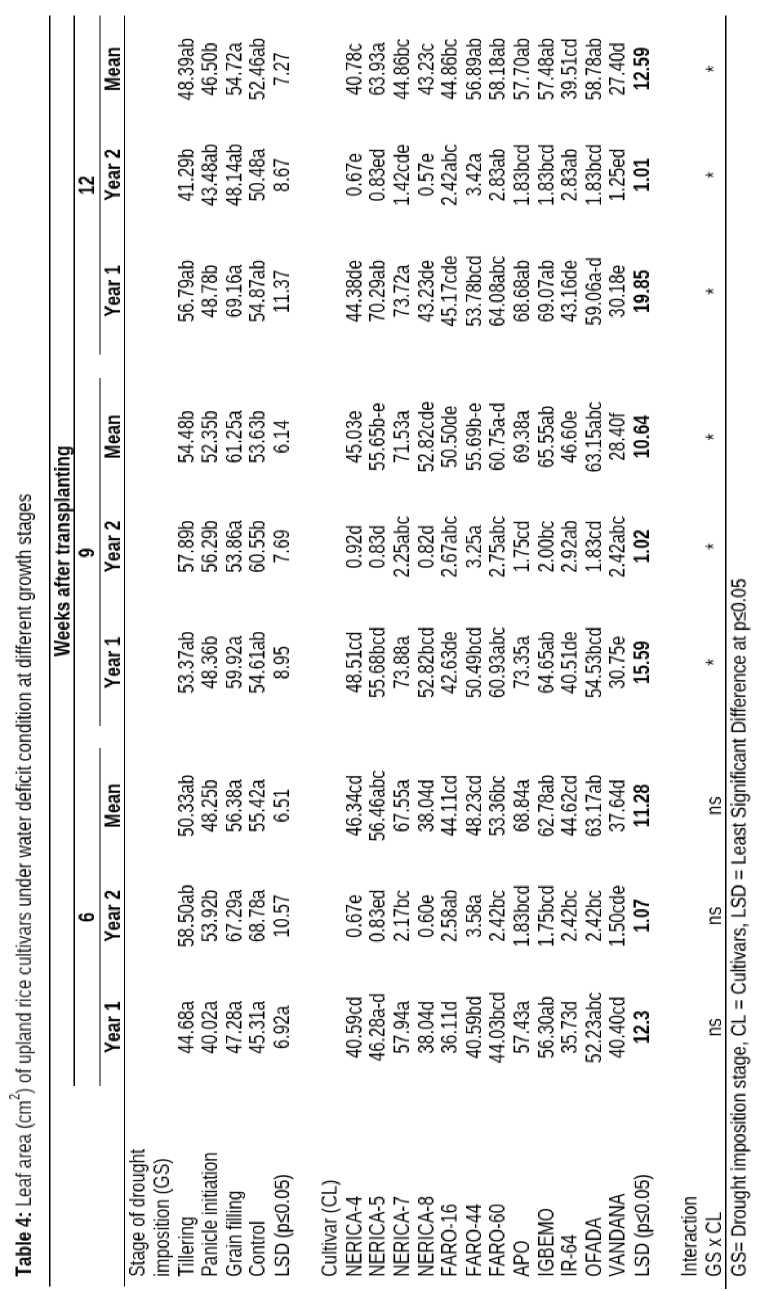
The growth stage at which drought was imposed had no significant effect on number of leaves produced at 6 WAT. However, number of leaves reduced significantly at 9 WAT, in plant water-stressed at grain filling, tillering and at panicle initiation stage while at 12 WAT plants water stressed at grain filling period had the highest number of leaves 11.42 which was significantly higher than plants water stressed at panicle initiation period (9.31) but was not significantly different from other growth stages (Table 3). The response of the different cultivars to water deficit condition was significant. IR-64 had the highest number of leaves (13.67, 17.38 and 17.54) at 6, 9 and 12 WAT but this was not significantly higher that number of leaves formed by FARO-44 and FARO-60.
Number of leaves produced was not significantly influenced by the interaction between growth stages when drought was imposed and different cultivars at 6 and 9 WAT except at 12 WAT when these factors influenced number of leaves significantly.
Effect of drought at different growth stages on leaf area of upland rice cultivars
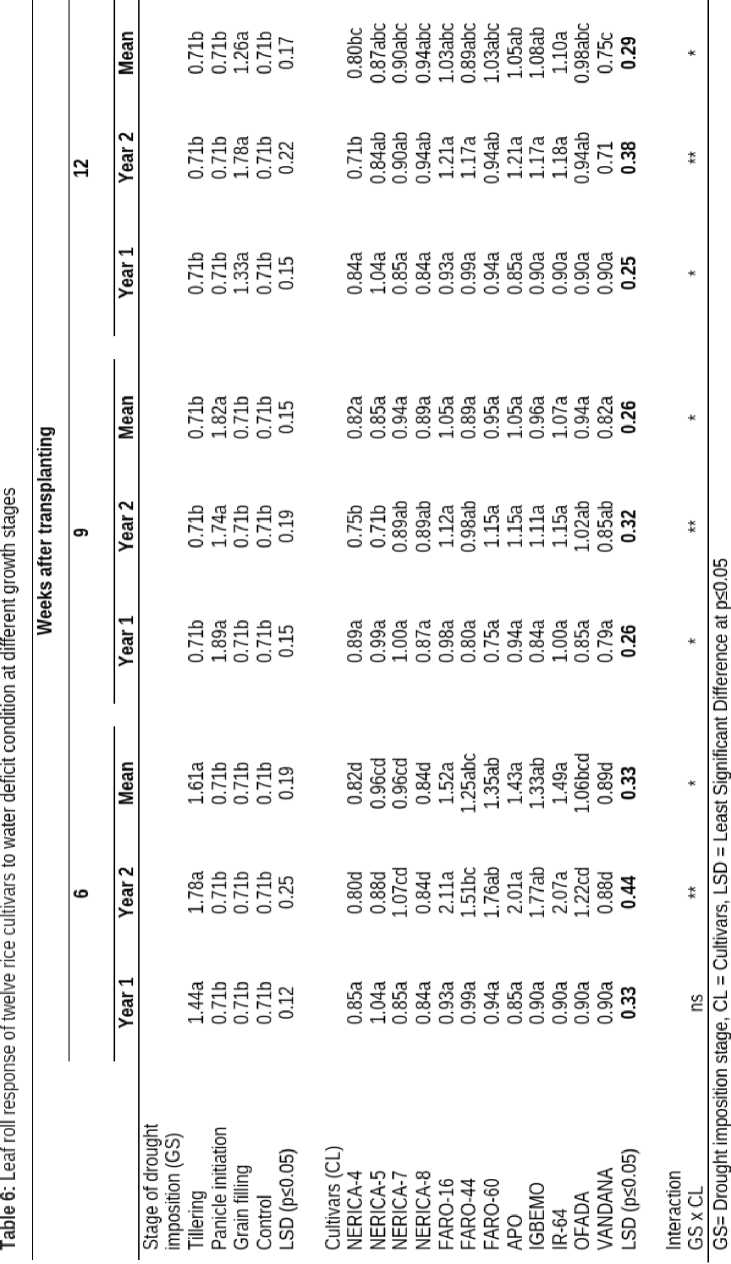
Water stress imposition across the growth phases had significant effect on the leaf area of upland rice cultivars. Plants stressed at tillering and panicle initiation had reduced leaf area which was significantly lower than leaf area obtained in plants water stressed at grain filling stage (Table 4).
Different upland rice cultivars responded differently to imposition of drought. Leaf area of APO was significantly higher (68.84 cm2) than other cultivars, but statistically similar to NERICA-7, IGBEMO, OFADA and NERICA-5 at 6 WAT. Also, at 9 WAT, NERICA-7 had highest leaf area (71.53 cm2) but this was comparable to leaf area of APO, OFADA and FARO-60 (Table 46). However, at 12 WAT, NERICA-5 had highest leaf area (63.93 cm2) which was not significantly higher than OFADA, FARO-60, APO, IGBEMO and FARO-44 at 12 WAT.
The interaction between varieties and growth stages had no significant influence on leaf area of upland rice cultivars at 6 WAT. However, these factors affected the leaf area significantly at 9 and 12 WAT (Table 4).
Number of tillers of twelve rice cultivars at different growth stages under water deficit condition
Water stress imposition had no significant effect on tiller formation at the phenological stages when drought was imposed. However, drought had significant effect on numbers of tillers produced by different upland rice cultivars. IR-64 cultivar formed higher number of tillers than other cultivars. Nevertheless, the number of tiller produced by FARO-44 and FARO-60 were similar to that of IR-64. NERICA-4 formed the lowest number of tillers across the growth phases (Table 5).
The interaction showed that growth period and cultivars had no significant influence on number of tillers at 6 and 12 WAT except at 9 WAT when these factors significantly influenced number of tillers formed (Table 5).
Leaf rolling scores of twelve rice cultivars at different growth stages under water deficit condition
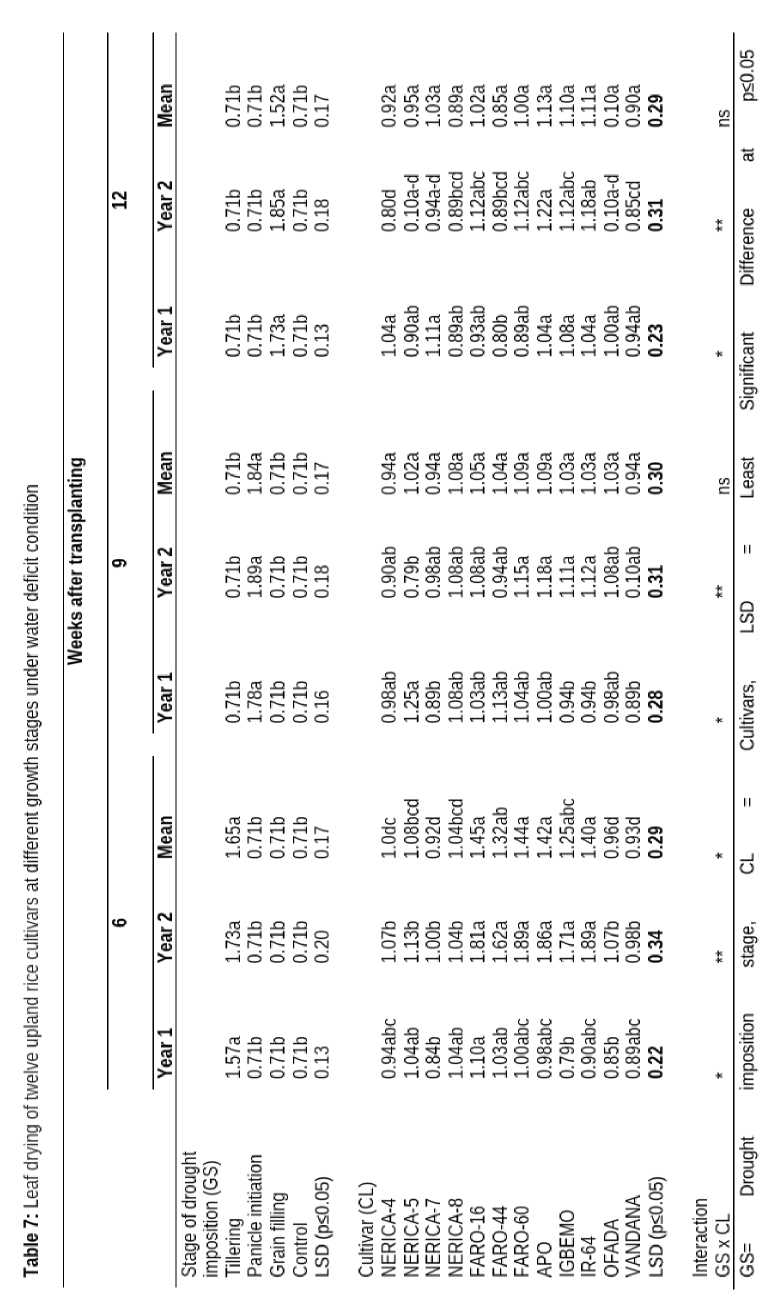
Imposition of drought caused the leaves of upland rice cultivars to roll significantly at all the phenological stages (Table 6). Leaf of upland rice cultivar rolled up (1.73) in response to drought mostly when drought was imposed at tillering at 6 WAT. At 9 WAT, leaf roll score was highest when drought was imposed at panicle initiation (1.84), while imposition of drought at grain filling resulted in higher leaf roll (1.52) at 12 WAT.
The interaction between growth stages and varieties caused significant leaf roll at 6 and 12 WAT (Table 6).
Leaf drying as an indicator of response to drought by rice cultivars at different growth stages
The response of different upland rice cultivars to drought as indicated by leaf drying differed significantly. At 6 WAT, FARO-16 had the highest leaf roll score (1.52) which was not significantly higher than the leaf roll score observed in IR-64, FARO-44, FARO-60 and APO cultivars. At 9 and 12 WAT there was no significant difference among the cultivars regarding leaf roll score (Table 7).
The interaction between growth stages and cultivars had no significant influence on leaf drying of rice cultivars at 9 and 12 WAT (Table 7).
Effect of Water-stress Imposed at Different Growth Stages on Yield Components of Upland Rice Cultivars
Imposition of drought at different growth stages reduced number of panicles, weight of panicles and number of spikelets per panicle formed significantly, relative to the yield components recorded in well-watered plants. Highest number of panicles (2.34), weight of panicle (2.40 g) as well as number of spikelets (11.75) was recorded in irrigated plants relative to plants water-stressed at various stages of growth (Table 4.10) Generally, yield components such as number of panicles (1.63), weight of panicle (1.78 g) and number of spikelets per panicle were lowest when the plants were water-stressed at panicle initiation (Table 6).
Drought imposition had significant effect on yield components of upland rice cultivars. VANDANA produced the highest number of panicles (2.88 g) which was significantly similar to number of panicles recorded in IR-64 and FARO-44 but statistically higher than number of panicles obtained in the other cultivars. Likewise, FARO-60 had highest weight of panicle (2.84 g) that was significantly similar to weight of panicle observed in IR-64, NERICA-4 and NERICA-5. imilarly, IR-64 had highest number of spikelets (13.30) per panicle and the lowest (6.58) was recorded in OFADA.

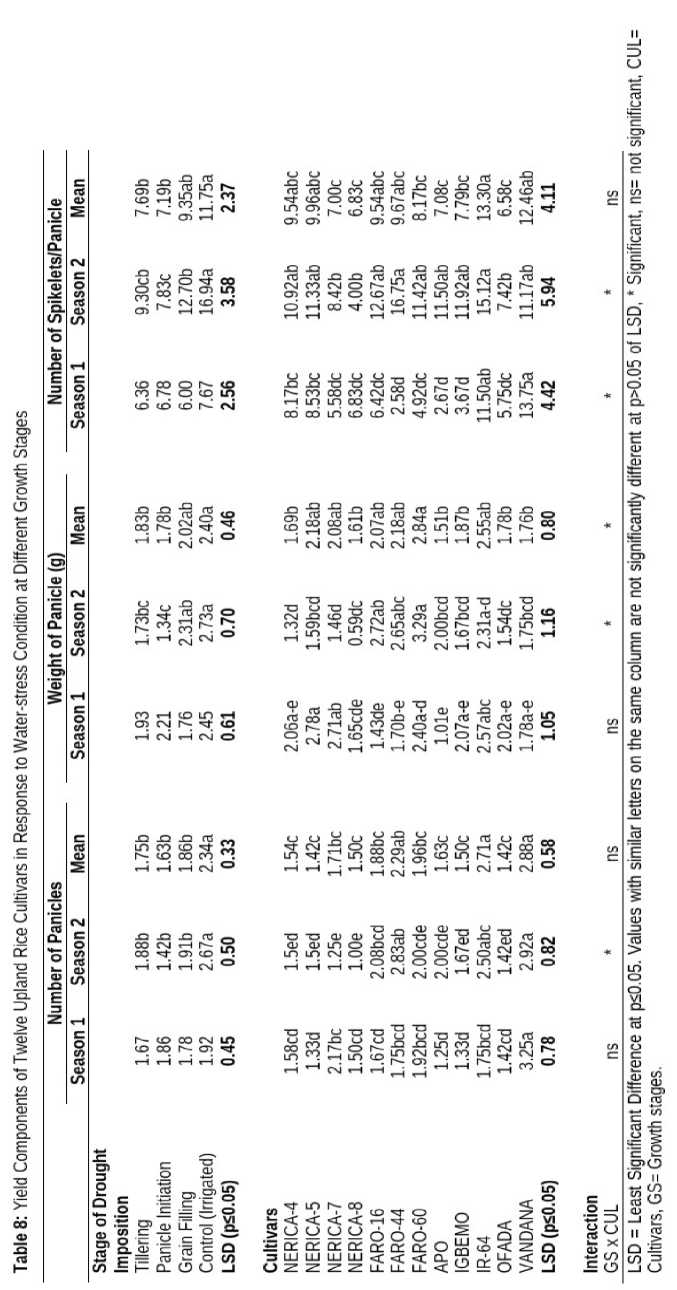
The number of panicles
and spikelets per panicle were not significantly influenced by interaction between growth stages when drought was imposed and upland rice cultivars. However, weight of panicle of different upland rice cultivars was significantly influenced by growth stage at which drought was imposed (Table 8).
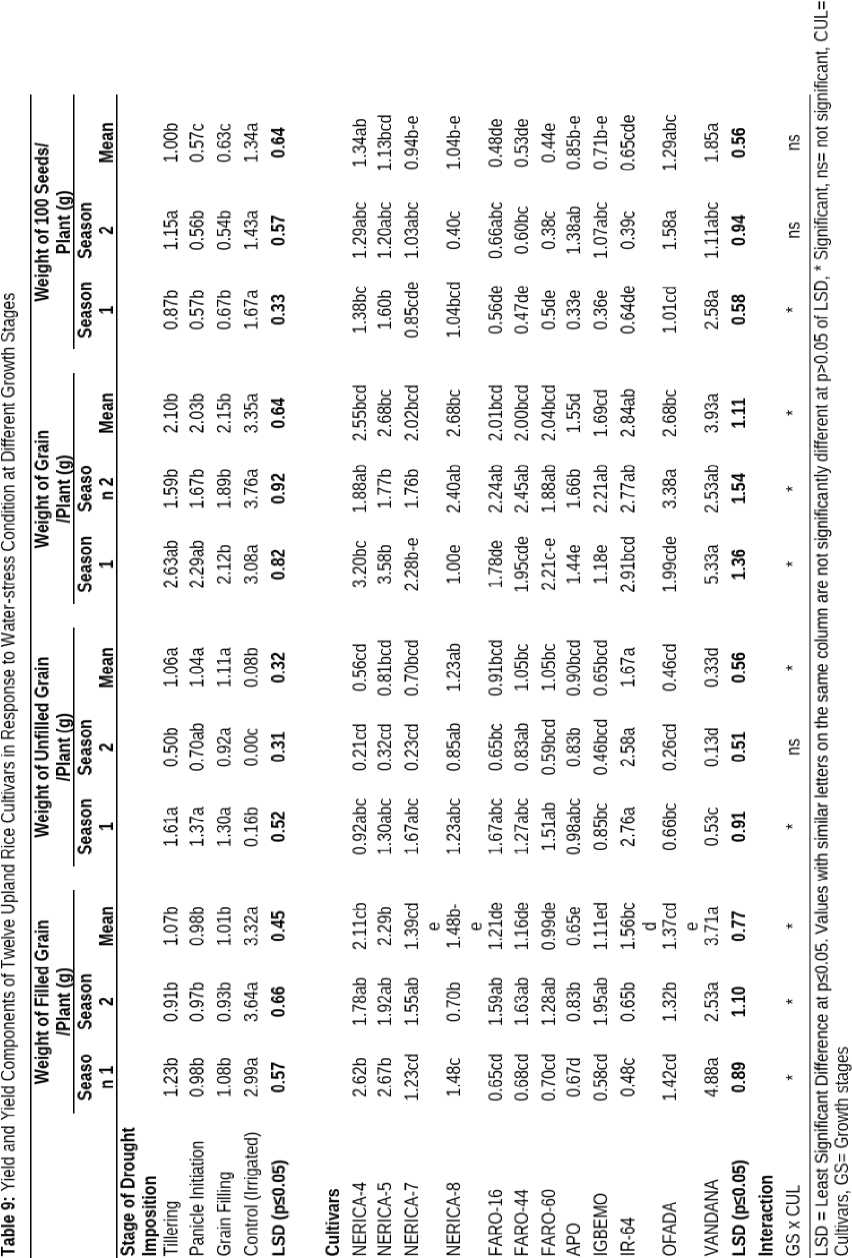
Filled grain, unfilled grain and grain weight of the different rice cultivars differed significantly in response to water limiting conditions (Table 9). Plants that were well watered (Control) had highest weight of filled grains (3.32 g) which was significantly higher than weight of filled grains obtained from plants subjected to drought at tillering, panicle initiation and grain filling stages. imilarly, well watered plants had lowest weight of unfilled grains (0.08 g) relative to plants subjected to drought at different growth stages. Highest weight of grains and 100 seeds was recorded in plants that were well watered compared to plants subjected to waterstress at different growth stages.
VANDANA had the highest weight of filled grain (3.71 g) which was statistically higher than other cultivars. Cultivar, IR-64 cultivar had highest weight of unfilled grain (1.67 g) which was significantly different from other cultivars but not significantly different from NERICA-8. VANDANA had the highest weight of grain (3.93 g) which was significantly similar to weight of grain recorded in IR-64. imilarly, weight of 100 seeds (1.85 g) was significantly higher in VANDANA than in any other cultivar.
The interaction between different upland rice cultivars and growth stages when drought was imposed had significant effect on all the yield components such as weight of filled grain, unfilled grain weight. However water-stress had no significant effect on weight of 100 seeds as indicated in Table 9.
Effect of Water-stress on Root Parameters of Upland Rice Cultivars at Different Growth Stages
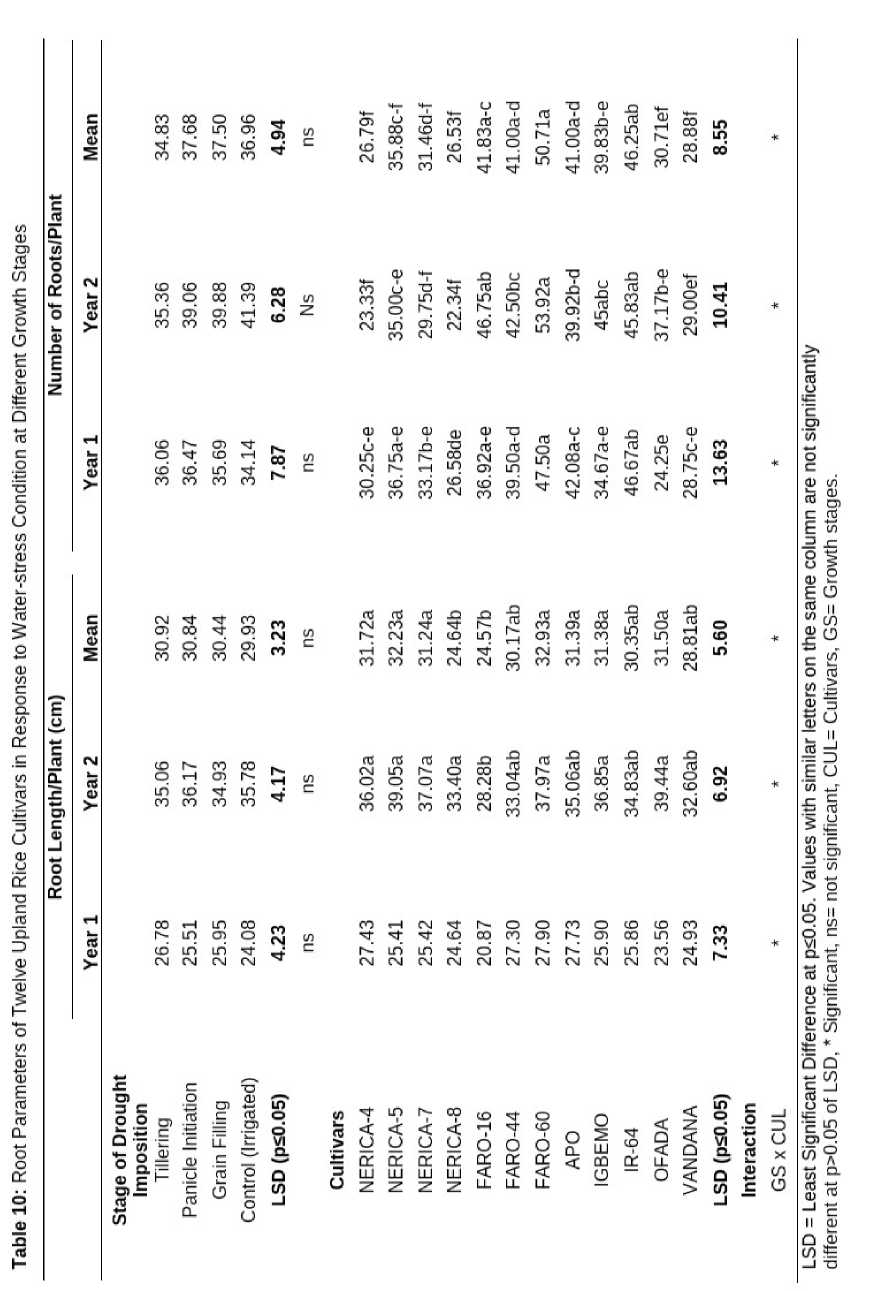
The length and number of roots of upland rice cultivars were not significantly influenced by drought imposed at different growth stages. However, among the cultivars, FARO-60 had the highest number of secondary roots (32.93) and main root length (50.71 cm), while FARO-16 recorded the lowest roots length (24.57 cm) and lowest number of roots was recorded in NERICA- 8 cultivar (26.53).
The interaction between growth stages when drought was imposed and different cultivars had significant influence on length and numbers of roots of upland rice cultivars (Table 10).
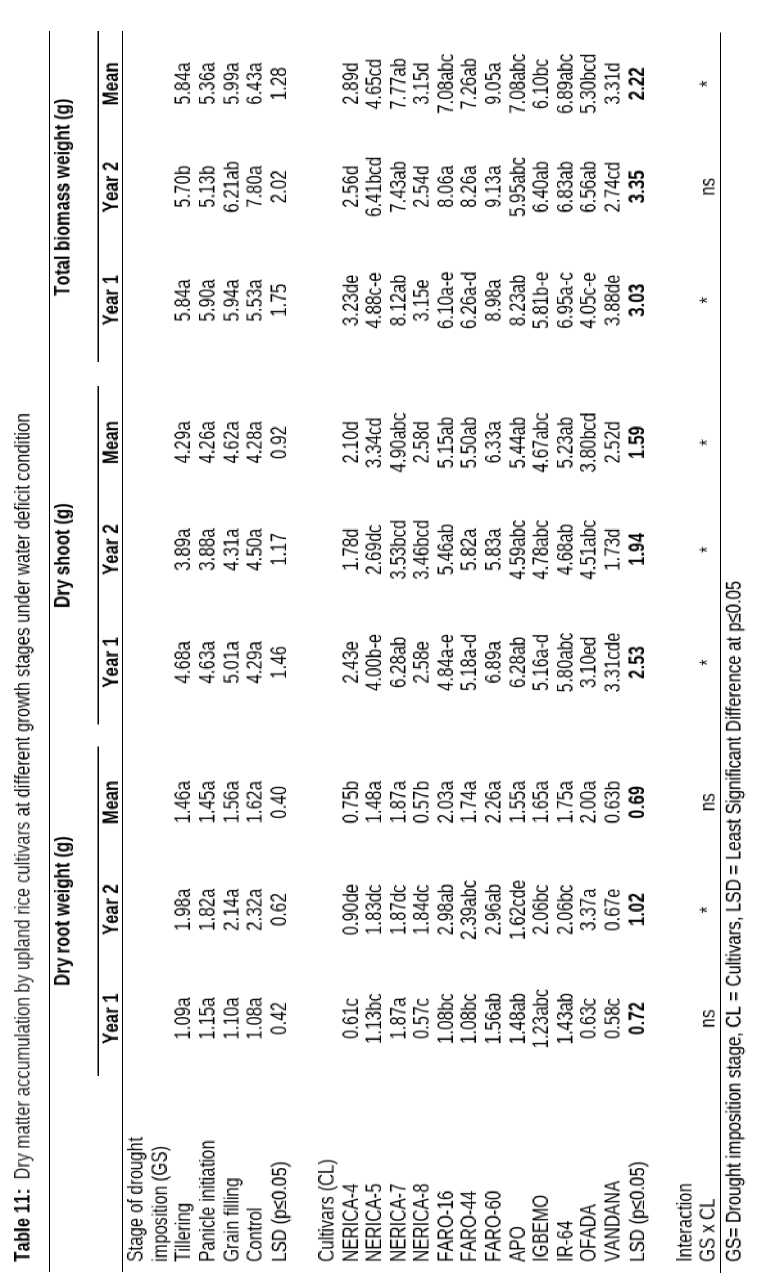
Dry Matter Accumulation by Upland Rice Cultivars Water-stress at Different Growth Stages
There was no variation in dry matter partitioning into shoot or root of upland rice cultivars exposed to water deficit condition at different growth stages. However, accumulation and partitioning of dry matter into shoot and root of different upland rice cultivars were significantly influenced by drought condition. NERICA-8, NERICA-4 and VANDANA had comparable weight of dry roots and shoot but these were significantly lower than dry matter into accumulated by shoot and root of the other cultivars (Table 11). Weight of shoot (6.33 g) and total biomass (9.05 g) of FARO-60 were significantly higher than that of NERICA-8, NERICA-4, OFADA, NERICA-5 and VANDANA.
The interaction between growth stages when drought was imposed and different upland cultivars had significant influence on dry matter accumulation into shoot and total biomass except weight of dry root which was not significantly influenced by interaction as presented in (Table 11).
DISCUSSION
Obviously, occurrence of water deficit condition at various growth stages adversely affected morphological development and yield of rice plant. However, the duration of stress, the stage of plant growth and development at which the plant was exposed to drought and severity contributed to plant response to abiotic stress. imilar reports by alam et al. (2001) showed that rice is highly susceptible to drought across growth stages. In our study, majority of the cultivars were susceptible to drought as implicated by poor growth performance at tillering and panicle initiation as well as reduced yield at grain filling. This is in consonance with the report of ingh et al . (2012) on wheat and Adejumo et al . (2018) on okra.
The better growth performance displayed by the following cultivars APO, OFADA, NERICA-7, FARO-44, FARO-60 and IR-64 in terms of plant height, number of leaves, leaf area and tiller formation suggests these cultivars may have the potential to tolerate drought irrespective of the growth stages. These cultivars should therefore be considered for further breeding program. Bouman and Toung (2001) reported that different cultivars might have different responses to the same drought stress, timing and intensity. It is therefore pertinent to consider water economy and its use efficiency by different rice cultivars in other to improve rice production in rice growing ecologies. imilar view had been shared by Datta et al. (2017) on the need to integrate rice biology with water economy, while considering rice designing program.
The stage of plant growth and development at which rice plant was exposed to drought contributed greatly to the observed effects. Drought affected morphological development at tillering, while grain formation was adversely affected when moisture stress was imposed at panicle initiation. Yield was greatly reduced when drought occurred at grain filling. The variation in response attested to the fact that water plays different roles in plant at varying stages of growth. Hence water deficit at any of the stage, will spell doom for the metabolic pathways responsible for the expected physiological characteristics. This could be attributed to plant metabolic activities, at the stage of active reproductive development, that are highly sensitive to changes in environmental factors more than at the vegetative stage ( aikumar et al ., 2016).Water deficit at these stages might have interfered with metabolic activities involved in these key processes necessary for plant development ( amarah, 2005; Adejumo et al ., 2018). Our view agrees with that of ah et al. (2020) on impact of drought at pre-flowering and grain filling stages in maize growth and yield. Moisture deficit at panicle initiation was more critical with damaging effects on yield of the crops. It was observed that rice showed appreciable level of tolerance to water shortage at grain filling stage than when imposed at tillering and panicle initiation stages. This observation contradicts the report of ah et al. (2020) who reported that water deficit affected maize similarly at all phenological stages.
Number of tillers reduced greatly, especially at tillering and panicle initiation stages relative to the well-watered plants and drought imposition at grain filling stage. Drought occurrence at the vegetative phase of rice plant may lead to reduced tiller ( ikuku et al., 2010)
as well as the number of leaves which is less detrimental to assimilate partitioning compared to water shortage at both anthesis and post anthesis (Hanum, 2017). imilarly the high number of tillers that was more in plants subjected to water deficit condition at grain filling phase than the control plants suggests that rice plant was more tolerant to water deficit at grain filling stage than other growth phases. Rahman et al. (2002) reported similar experience that tiller numbers of rice plant was reduced tremendously under moisture deficit condition at different growth stages. When water shortage occurs at the tillering stage, the cell division and expansion processes are impaired resulting in poor growth and development according to Dada et al . (2018) and Wicaksono et al. (2022). The high number of tillers and leaves displayed by IR-64 cultivars, a susceptible cultivar shows that the cultivar has potential for early growth or may display those attribute in response to moisture stress as an escape mechanism. Mattioli et al. (2020) and Wicaksono et al. (2022) reported that plant may accelerate completion of their life cycle under water stress situation. The ability of the plant under drought to maintain internal water content is crucial for plant survival. These help in maintaining cell turgor by conserving the internal water content and scavenging reactive oxygen species (RO ) produced in response to environmental challenge as opined by Mattioli et al. (2020).
Main root number and length were higher in plants under water stress at tillering and panicle initiation phases than those subjected to water deficit condition at grain filling. This implies that moisture stress was more severe at these phases as proliferation of root mass indicates massive production of roots necessary for scavenging for moisture within the rhizosphere. The massive root production may encourage quick recovery after the drought imposition. imilarly, production of long and numerous roots by FARO-60 and IR-64 suggests that the cultivars were highly susceptible to water deficit condition and hence develop numerous roots as a drought adaptation strategy to survive the harsh environmental condition. This is similar to the findings of alam (2001) on rice genotypes and Morales et al. (2013) on raspberry. High root numbers under water deficit condition has been reported to enhance nutrient uptake better than plants with poorly developed roots (Blum, 1996; Barnabas et al., 2008).
Dry matter accumulation was reduced by moisture stress mostly at tillering and panicle initiation stage possibly due to impaired nutrient uptake, which limits photosynthate formation and partitioning into biological and economic yield. The observed increased partitioning of dry matter into root is an indication of drought avoidance in a bid to maximize uptake of available water (Barnabas et al., 2008). Also water stress at tillering effectively reduced total biomass; this may be due to decrease in photosynthesis rate and dry matter accumulation. However, high dry matter was partitioned in FARO’ cultivars than any other cultivars.
Among the cultivars, VANDANA was more prolific with huge grain yield. This suggests that the cultivar is likely to be tolerant to moisture stress, while IR-64 and APO had lowest grain yield suggests the likelihood of these cultivars susceptibility to drought. The differences in grain yield may be directly linked to the recovery rate after stress imposition at different stages. Recovery from water stress is a function of growth stage and severity of the stress (Kato et al. 2007). However, panicle number and grain yield was reduced drastically at panicle initiation stage compare to other growth phases. Drought has been found to reduce yield in upland rice field up to 50% according to Dada et al . (2018). Cereal crops showed similar response to drought at all phases but became more intense or severe at grain filling stage (Barnabas et al., 2008).
Plants respond to drought at physiological stages and this is indicated by alteration in morphological responses such as leaf rolling, dry tips and dead heart. This could be the possible reasons why upland rice cultivars that were subjected to drought stress at panicle initiation period had the highest leaf roll and dry score all through the growth period. Alteration of leaf morphology under drought included the direct effect on photosynthetically active radiation and net photosynthetic rate ( yuhada and Jahan, 2016; Khairi et al., 2016). imilarly among the various cultivars used for the study, cultivar IR-64 had highest leaf rolling and drying score which suggests that the cultivar was highly susceptible to drought stress and this study is in line with previous research carried out by Bouman and Toung (2001) who reported that different cultivars might have different responses to the same drought stress, timing and intensity.
CONCLUSION
Oryza species is more sensitive to water deficit condition at tillering and panicle initiation growth stages than at grain filling phase. Drought at panicle initiation stage reduced grain yield than at tillering phase in rice cultivars. Also VANDANA cultivar had the yield performance compared to other cultivars. The studies conclude that moisture deficit condition should be avoided at the critical growing period like panicle initiation as irrigation schedule should be incorporated into rice growing system.
CONFLICTS OF INTEREST
The authors declare that they have no potential conflicts of interest.
