Находки раковин пресноводных моллюсков в верхнепалеолитическом культурном слое пещеры Двойная (Северо-Западный Кавказ)
")
Автор: Успенская О.И., Леонова Е.В., Фролов П.Д.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Рубрика: Каменный век
Статья в выпуске: 259, 2020 года.
Бесплатный доступ
В статье рассматривается коллекция перфорированных раковин пресноводных моллюсков, происходящих из верхнепалеолитического культурного слоя пещеры Двойная (Северо-Западный Кавказ). Приведена характеристика видового состава моллюсков, описаны их морфологические особенности и реконструированы способы перфорирования. Трасологический анализ позволил выявить, что помимо перфораций, совершенных человеком, на раковинах есть следы природного сверления, связанные, предположительно, с деятельностью неизвестного хищника, а также следы «погрызов» неустановленного генезиса. Свидетельства сверления раковин пресноводных моллюсков хищниками редки и плохо изучены. Характер следов износа поверхности раковин позволяет с наибольшей долей вероятности реконструировать их использование в качестве бусин в составе ожерелья, браслета или пронизок в декоративном оформлении одежды.
Каменный век, верхний палеолит, северо-западный кавказ, пещера двойная, пресноводные моллюски theodoxus, пресноводные моллюски lithoglyphus naticoides, трасология, украшения, природная перфораци
Короткий адрес: https://sciup.org/143173105
IDR: 143173105
Finds of fresh-water mollusks shells from the upper paleolithic occupation layer in Dvoynaya cave (the Northwestern Caucasus)
The paper examines the collection of perforated shells of fresh-water mol-lusks found in the Upper Paleolithic occupation layer in Dvoynaya Cave (the northwestern Caucasus). It provides characteristics of mollusk species, describes their morphological traits and reconstructs methods of perforation. The tracewear analysis evidences that, in addition to perforations made by humans, the shells bear traces of biotic drilling, supposedly, left by an unknown predator as well as biting marks of unidentified origin. Evidence of drilling predation found on the shells of freshwater mollusks is not ample and well-studied. The nature of wear traces on the shell surface makes it possible to reconstruct their use as beads of a necklace, a bracelet or tubular beads used as elements of clothes decorative design.
Текст научной статьи Находки раковин пресноводных моллюсков в верхнепалеолитическом культурном слое пещеры Двойная (Северо-Западный Кавказ)
Находки перфорированных раковин на стоянках каменного века не редки, но представляют интерес при решении проблем мобильности древнего населения, а также являются источником для реконструкции неутилитарной деятельности первобытных сообществ. Исследованиями 2007–2014 гг. в пещере Двойная выделено три культурных слоя, материалы которых отражают три волны заселения Губского ущелья на Северо-Западном Кавказе на рубеже плейстоцена и голоцена (Леонова, 2015; Александрова, Леонова, 2017). В нижней части верхнепалеолитического культурного слоя (слой 7) – наиболее раннего этапа заселения пещеры, ориентировочно датированного по данным радиоуглеродного анализа около 14 тыс. лет, обнаружены раковины пресноводных брюхоногих моллюсков http://doi.org/10.25681/IARAS.0130-2620.259.100-113
с просверленными и пробитыми отверстиями, а также без следов перфорации, всего на стоянке найдено 30 раковин. Кроме этого, есть три неопределимых обломка устья раковин.
На основании морфологии и характера рисунка раковин удалось определить несколько видов моллюсков: Theodoxus fluviatilis (Linnaeus, 1758) (16), Theodoxus major Issel, 1865 (8), Theodoxus sp . (6) и одна раковина Lithoglyphus naticoides (C. Pfeiffer, 1828).
Theodoxus fluviatilis – очень изменчивый вид, что проявляется в различии форм, параметров, а также широкой вариабельности рисунка и окраски раковин ( Glöer, Pešić , 2015; Sands et al. , 2020). Этот вид сейчас широко распространен в реках и озерах Европы, Северной Африки, Передней Азии и Закавказья (Ibid . ). На территории России он встречается в бассейне Азовского, Черного, а также Балтийского морей ( Кантор, Сысоев , 2006). В рассматриваемой коллекции можно выделить три морфотипа, отнесенных к этому виду. T. fluviatilis – раковины преимущественно белесовато-бежевых и желтых оттенков без рисунка, который, по всей видимости, не сохранился (рис. 1: 17, 22, 25, 27, 31, 32 ). Одна светло-коричневая раковина с темно-коричневыми зигзагами соотносится с морфотипом T. euxinus (рис. 1: 14 ), характерного для пресных и солоноватых вод Западного Причерноморья ( Анистратенко и др. , 1999). И еще четыре раковины темно-серого цвета (вероятно, обожженные), похожи на морфотип T. cf. subthermalis (рис. 1: 12, 26, 28, 33 ), который обитает в реках, ручьях, родниках и проточных озерах восточного побережья Азовского и Черного морей и приурочен к Таганрогской зоогеографической провинции (Там же).
Вид Theodoxus major также обладает высокой изменчивостью ( Sands et al. , 2020), встречается в Каспийском и Азовском морях, на нижней Волге и нижнем Дону, ранее отмечался в Аральском море (но вымер во втор. пол. XX в.), кроме того, его находки зафиксированы в Армении, Азербайджане и Иране ( Анистратенко и др. , 1999; Plotnikov et al. , 2016; Sands et al. , 2020). В коллекции раковин из пещеры Двойная этот вид представлен только одним морфоти-пом – Theodoxus pallasi (= Theodoxus astrachanicus ) ( Anistratenko et al. , 2017) (рис. 1: 1, 9, 13, 15, 24 ), раковины бежевые или светло-коричневые, с окраской в виде красно-коричневых зигзагов или волнистых полос. В Черном море Theodoxus pallasi , вероятно, вымер в Неоэвксинское время (~ 24–11 тыс. л. н.) ( Янина , 2012).
Пресноводный брюхоногий моллюск Lithoglyphus naticoides (рис. 1: 6 ) на территории России сейчас распространен в реках бассейна Черного и Азовского морей ( Vinarski, Kantor , 2016).
Среди изучаемой выборки раковин Theodoxus sp. определяется несколько размерных групп: наиболее крупные раковины имеют высоту 9–9,8 мм, ширину 6–6,5 мм, толщину 4–5 мм. Самые маленькие раковины высотой 5 мм, в ширину 3,5–4 мм, толщиной 2,5 мм. В среднем преобладают раковины высотой 7–8 мм, шириной 5–6 мм и толщиной 3,5–4 мм (рис. 1). Поверхность трех раковин сильно корродирована, белесая (рис. 1: 17, 24, 25 ), еще несколько раковин с перфорациями частично окрашены красноватым веществом (предположительно, охра) (рис. 1: 11, 22, 29, 31 ) .
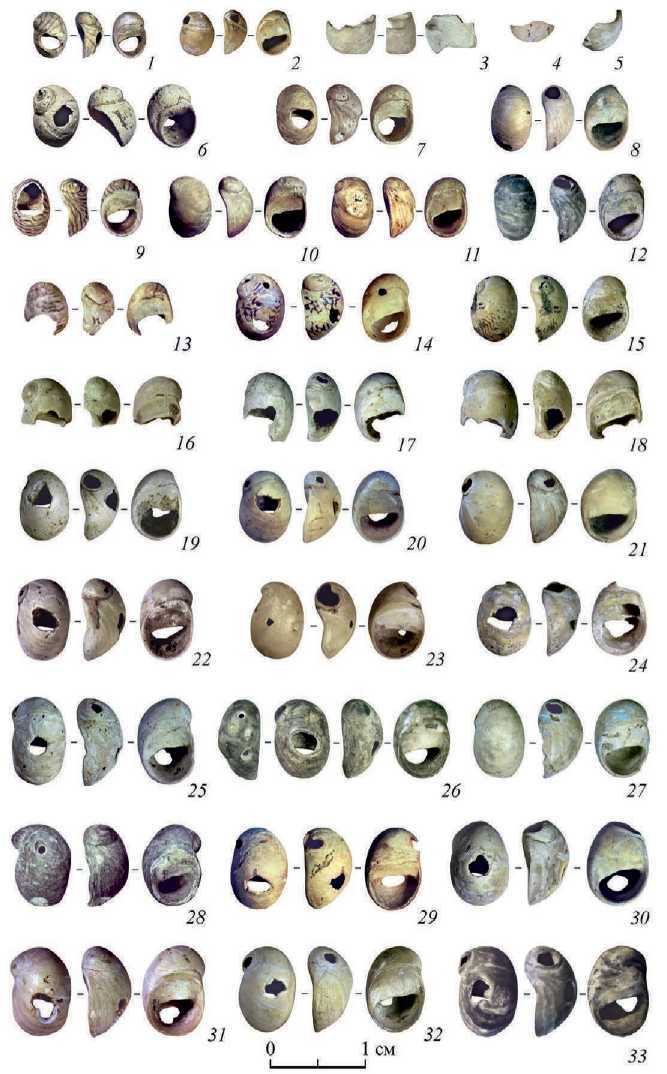
В коллекции 26 целых раковин, 4 – с поперечным сломом от края устья и примерно до середины последнего оборота (рис. 1: 13, 16–18 ). На одном из трех неопределимых обломков последнего оборота читается характерная угловатая кромка перфорированного отверстия (рис. 1: 3 ). Сломы проходят примерно по первой трети последнего оборота по границе отверстий. На сломанных раковинах видны лишь небольшие участки кромки отверстий.
Только две раковины не имеют отверстий (рис. 1: 10, 11 ), на остальных – от одного до четырех, которые можно распределить по двум группам: искусственные и естественные (рис. 2). К естественным отнесены отверстия через макушку раковины, которые могли образоваться в постседиментационный период, при носке раковин в составе украшения, либо антропогенное воздействие невозможно установить (рис. 2: 2 ). Подобные частичные или полные выломы макушки наблюдаются на 18 раковинах: Theodoxus sp. (17) и Lithoglyphus naticoides (1). Форма этих отверстий преимущественно овальная или округлая, частично повторяет очертания границ первых оборотов, размеры отверстий сильно варьируют от 0,5 до 3 мм в поперечнике.
У 16 целых раковин на расстоянии 3,5–5 мм от края устья пробиты отверстия ( Theodoxus sp. (15) и Lithoglyphus naticoides (1)), которые имеют различную форму: каплевидную, многогранника, подтреугольную. Их размеры в среднем 2,5–3,5 мм в поперечнике (рис. 2: 1 ).
На четырех раковинах обнаружены сквозные округлые отверстия, диаметр которых около 1 мм: на двух раковинах по два отверстия (рис. 1: 14, 26 ) и еще на двух – по одному (рис. 1: 28, 31 ). Стенки трех отверстий представляют собой абсолютно гладкую «оплавленную» поверхность без характерных линейных следов сверления (рис. 2: 3 ; 4: 6 ). На одной раковине мелкая «сверлина» располагается на последнем обороте со стороны, обратной устью, поверхность канала шершавая, видна структура стенки раковины, представляющая собой карбонатные пластины перекрещенно-пластинчатого слоя, напоминающие линейные следы от сверления (рис. 2: 3 ; 4: 7 ). Сохранившийся участок аналогичной «сверлины», только в несколько раз крупнее, зафиксирован на одной раковине, пробитой человеком непосредственно по этому отверстию (рис. 1: 31 ; 2: 5 ).
На трех раковинах были зафиксированы деформации, напоминающие «лунки» подокруглой формы, с неровным изъеденным дном, которые, предположительно, также связаны с деятельностью хищных сверлильщиков. На одной раковине две лунки, диаметром 1 мм, располагающиеся по бокам от макушки
Рис. 1 (с. 102). Пещера Двойная.
Находки раковин пресноводных брюхоногих моллюсков из верхнепалеолитического слоя
1, 9, 13, 15, 24 – Theodoxus major Issel, 1865 (морфотип – T. pallasi ); 2, 10, 19, 21, 30 – Theodoxus sp.; 3–5 – неопределимые обломки раковин; 6 – Lithoglyphus naticoides (C. Pfeiffer, 1828); 7, 20, 29 – Theodoxus cf. major Issel, 1865 (морфотип – T. pallasi ); 8, 11, 16, 18, 23 – Theodoxus cf. fluviatilis (Linnaeus, 1758); 12, 26, 28, 33 – Theodoxus fluviatilis (морфотип – T. cf. subthermalis ); 14 – Theodoxus fluviatilis (Linnaeus, 1758) (морфотип – T. euxinus ); 17, 22, 25, 27, 31, 32 – Theodoxus fluviatilis (Linnaeus, 1758)
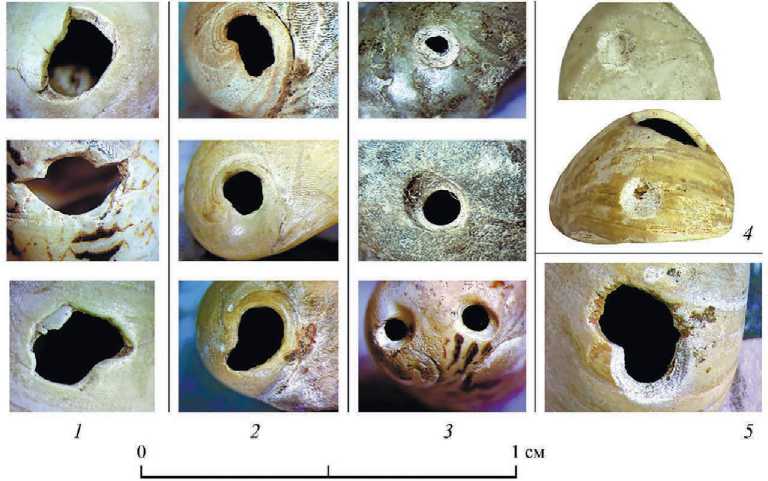
Рис. 2. Пещера Двойная.
Виды отверстий на раковинах пресноводных моллюсков Theodoxus
1 – намеренно пробитые человеком по центру раковины с внешней стороны; 2 – выломы вершины первого завитка раковины; 3 – «сверлины», связанные с деятельностью неизвестного хищника; 4 – «выгрызы» в виде неглубоких округлых лунок; 5 – отверстие, пробитое человеком по предположительно хищнической «сверлине». Все фотографии поверхности раковин приведены к единому масштабу
(рис. 1: 28 ). Еще на одной раковине подовальная (диаметром около 1 мм) «лунка» располагается на границе колумелярной площадки и последнего оборота (рис. 1: 8 ; 2: 4 ), на третьей – на последнем обороте ближе к устью, чуть менее 1 мм в диаметре (рис. 1: 7; 2: 4 ; 4: 8 ). Глубина всех лунок составляет доли миллиметра.
Большинство раковин в коллекции были обнаружены in situ в нижней части верхнепалеолитического культурного слоя, а некоторые извлечены из промывки культурного слоя. Двенадцать раковин (8 целых и 4 обломка) обнаружены у очага у западного борта на кв. l,m-3, 4 (рис. 3). Второе скопление из 13 раковин (11 целых и 2 обломка) зафиксировано ближе к СВ борту раскопа на квадратах k,l-6,7. Две раковины происходят из кв. o-5,7 привходовой части пещеры, где отложения частично подвержены эрозии, поэтому изначальная стратиграфическая позиция находок неизвестна. Остальные 6 раковин найдены в верхнепалеолитическом слое, но скоплений не образуют. Следует отметить, что раковины без отверстий залегают в том же контексте, что и намеренно пробитые человеком или сломанные по пробитому отверстию, каких-либо закономерностей в их расположении не определяется.
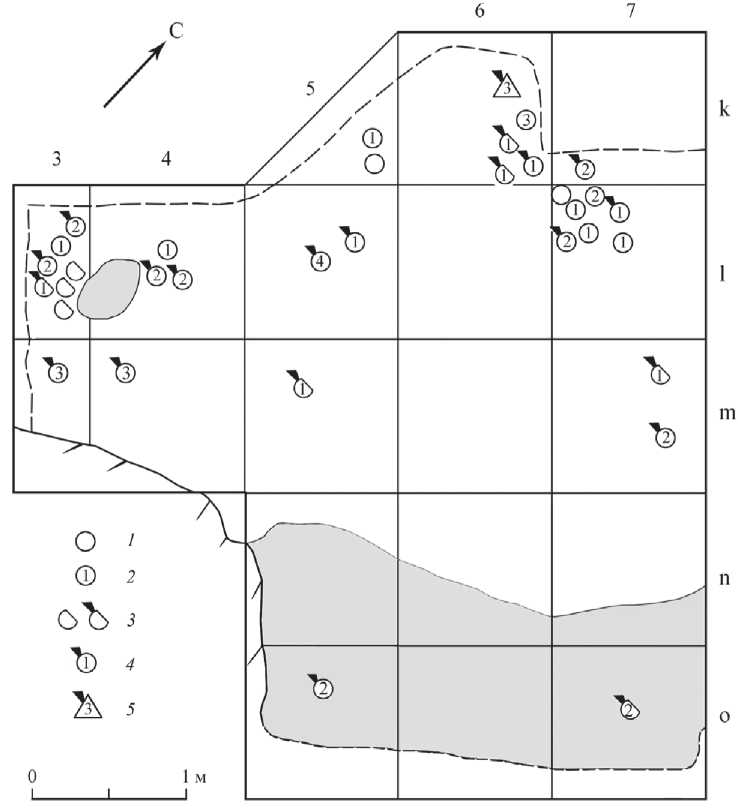
Рис. 3. Пещера Двойная. Планиграфическое распределение раковин Theodoxus и Lithoglyphus naticoides в верхнепалеолитическом культурном слое
1 – Theodoxus без отверстий; 2 – Theodoxus с отверстиями (цифрой внутри символа обозначено число отверстий); 3 – Theodoxus с пробитым человеком отверстием; 4 – обломки Theodoxus ; 5 – Lithoglyphus naticoides с пробитым человеком отверстием
На основании проведенного трасологического анализа коллекции удалось определить две области со следами износа. Первая формируется на участке между стенкой пробитого отверстия и естественным краем устья в результате трения волокна/ремешка, на который нанизана бусина. На этих участках бусин видны макроскопические следы износа, представленные истиранием естественной поверхности раковины, утончением перемычки между отверстием и краем
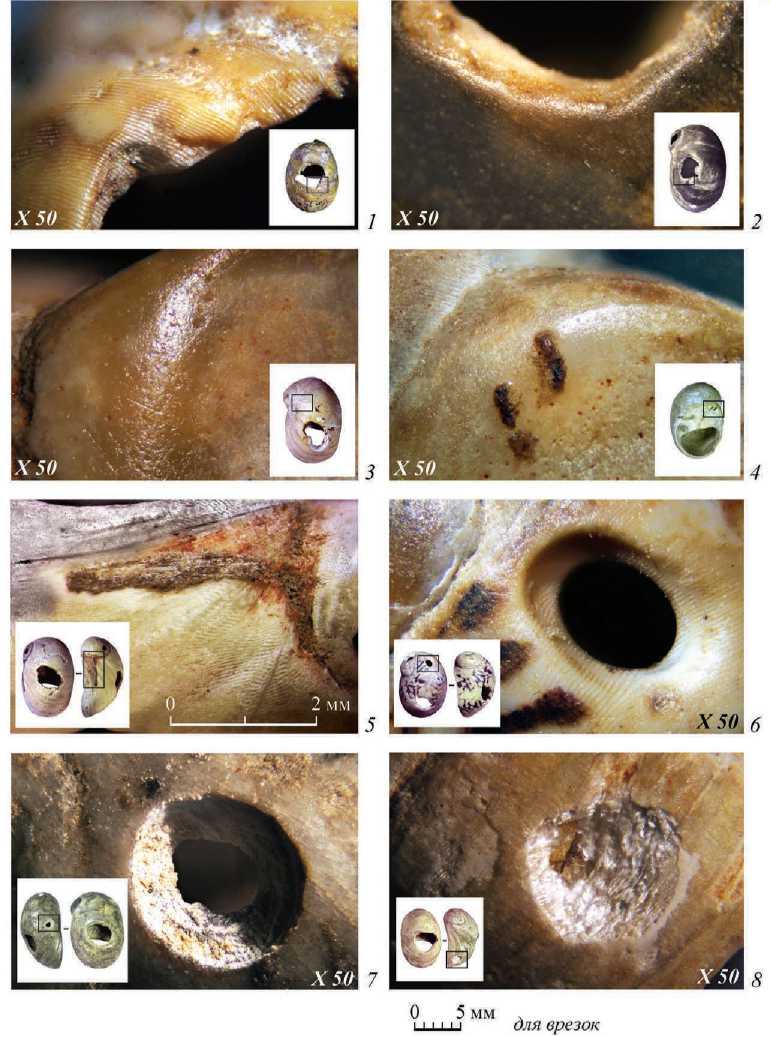
Рис. 4. Пещера Двойная. Следы износа на раковинах Theodoxus
1, 2 – следы заглаженности от нитей на краях пробитых отверстий; 3, 4 – заполировка поверхности; 5 – остатки предположительно органического волокна и прокрашенность поверхности охрой; 6, 7 – «сверлины», оставленные неизвестным хищником; 8 – повреждения в виде округлых «выгрызов»
устья, развитием заметной вогнутости и сглаживания определенного участка стенки отверстия (рис. 4: 1, 2 ). Расположение этих следов на участке ближе к краю устья раковины свидетельствует о том, что самая тяжелая часть корпуса раковины (у макушки) была обращена вниз. Наиболее интенсивная степень износа для бусин, подвешенных подобным образом, связана с разрушением раковины, идентичным тем, что наблюдаются на сломанных фрагментах из пещеры Двойной (рис. 1: 13, 16–18 ).
Вторая область со следами износа обнаруживается на участке поверхности между пробитым отверстием и вершиной первого завитка раковины. Поверхность раковины гладкая, с макроскопическим блеском (рис. 4: 3, 4 ). Тесный контакт бусин между собой, а также вероятные соударения раковин вершинами могли приводить к их систематической поломке, что и фиксируется для раковин из пещеры Двойной в виде естественных выломов первого завитка.
На основании экспериментальных данных ( Álvarez Fernández , 2006; Benghiat et al. , 2009; Stiner , 2010; Stiner et al. , 2013; Tátá et al. , 2014; André, Bicho , 2016) можно заключить, что отверстия на раковинах Theodoxus sp. и Lithoglyphus naticoides из пещеры Двойная с наибольшей долей вероятности созданы как при помощи удара с посредником (отверстия с неровными очертаниями), так и посредством прямого давления жалом перфоратора на внутреннюю стенку (отверстия округлые и более мелкие). Фиксируются характерные для этих техник сломы различных конфигураций ( Rigaud , 2013. Р. 220). Разрушение раковин, которое наблюдается на нескольких обломках, могло произойти как при изготовлении отверстия, так и при последующем использовании раковины в процессе носки. На одной раковине зафиксированы признаки сверления и пролома корпуса по сверлине с внешней стороны, однако сверлину нельзя однозначно определить как созданную человеком (рис. 1: 31 ; 2: 5 ). Интересно, что экспериментальные данные относительно продолжительности носки раковин в составе ожерелья и браслетов говорят о том, что износ, наблюдаемый на археологических образцах, мог сформироваться только в результате многолетнего использования украшения ( Mărgărit et al. , 2018. P. 2105). В археологических материалах прослеживается различная продолжительность использования некоторых раковин, обнаруженных в единых комплексах, что может свидетельствовать в пользу постепенного изменения набора бусин в ожерелье или ином украшении со временем ( Rigaud , 2013. Р. 218). Характерная заполировка краев и поверхности от постоянного трения со шнурком, телом или одеждой позволяет говорить о достаточно длительном использовании некоторых из рассматриваемых бусин пещеры Двойная (рис. 1: 26, 33 ; 4: 1–4 ). На нескольких бусинах удалось обнаружить красно-коричневые пятна и полосы, предположительно, обрывки волокон или нитей (рис. 4: 4, 5 ).
За время работы Губской экспедиции ни одного экземпляра речного моллюска Theodoxus в р. Губс встречено не было, но специальных поисков не предпринималось. В соседних речных бассейнах они встречаются. Наличие в культурном слое пещеры раковин без отверстий и на начальном этапе изготовления позволяет предположить их местное происхождение. Трасологически не удалось проследить, собирали ли древние насельники живых моллюсков и затем высушивали или подбирали уже пустые раковины.
Следы перфорирования на раковинах моллюсков и других скелетных организмов известны с кембрия, основной причиной возникновения перфораций считается хищническая деятельность. Существует множество организмов, способных к сверлению и декальцинированию раковины, таких как водоросли, грибы, лишайники, губки, различные членистоногие, а также моллюски ( Boekschoten , 1966). Одной из основных групп ископаемых и современных хищных сверлильщиков помимо осьминогов, круглых и плоских червей являются морские брюхоногие моллюски, среди которых способностью к сверлению обладает ряд видов, объединенных предпочтением к условиям нормальной солености (32–35 ‰), в то время как соленость современного Черного моря составляет 18 ‰, а Каспийского – 12–13 ‰ ( Kowalewski , 2002).
Существуют редкие свидетельства наличия перфораций на раковинах гастропод, собранных из миоцена, плиоцена, плейстоцена, а также из субфоссильных отложений в основном юга СССР ( Ильина , 1987; 2001). Отверстия с гладкими оплавленными краями, наблюдаемые на раковинах Theodoxus из рассматриваемой коллекции, наиболее близки к перфорациям, производимым хищными морскими брюхоногими моллюсками (рис. 4: 6, 7 ). Похожая сглаженность поверхности канала, как бы оплавленная, получается в результате протравливания кислотой хищного моллюска, однако для пресноводных водоемов вопрос о деятельности хищных моллюсков остается открытым ввиду слабой изученности этого явления. Еще одним возможным объяснением наличия хищнической деятельности на раковинах является их возможное принесение из солоноватых ареалов обитания, например, из акватории черноморских лиманов, где свидетельства хищнической деятельности зафиксированы на раковинах Theodoxus fluviatilis , однако вне археологического контекста ( Бондарев , 2016. С. 19. Рис. 1). Происхождение части не-досверленных лунок с неровной, как бы выскобленной поверхностью, вероятнее всего, также связано с деятельностью неустановленного хищника (рис. 4: 8 ).
Находки раковин моллюсков на стоянках и в погребениях каменного века нередки и служат источником информации о палеоэкологической обстановке в древности, о возможных путях миграции населения или обмена.
История использования раковин пресноводных моллюсков в качестве украшений первобытными людьми на территории Евразии начинается в верхнем палеолите и продолжается до неолита включительно ( Harzhauser et al. , 2007; Hutterer et al. , 2014).
В Европе первые свидетельства изготовления украшений из раковин моллюсков, в том числе Theodoxus и Lithoglyphus naticoides , относятся к началу верхнего палеолита (см., например: Kuhn et al. , 2001. Р. 7642; Stiner et al. , 2013. P. 387; Синицын , 2016. С. 324, 328; Borić, Cristiani , 2019. P. 223; и т. д.). Бусины из раковин моллюсков рода Theodoxus встречаются в целом ряде ближневосточных и европейских памятников конца плейстоцена – начала голоцена ( Яковлева , 2012; Kandel et al. , 2018; Mărgărit et al. , 2018; Street et al. , 2002; Yanevich et аl. , 2009; и т. д.).
На Северо-Западном Кавказе находки бусин из раковин речных моллюсков, часть которых, судя по морфологии, принадлежит к роду Theodoxus известны в коллекции слоя 1–3 Мезмайской пещеры ( Голованова, Дороничев , 2012. С. 64. Рис. 5: 2 ). Похожая раковина с пробитым отверстием найдена в слое 7 навеса Сосруко в Баксанском ущелье ( Голованова и др. , 2019. С. 13, 15. Рис. 4: III-2 ).
В этих публикациях авторами приводятся определения раковин как Helicidae gen., Succineidae gen. ( Succinea sp.) и Pupillidae gen. (определение А. Л. Чепа-лыги для находок из Мезмайской пещеры), а раковина из Сосруко идентифицирована как Succinea sp., но судя по морфологии экземпляров, представленных на иллюстрациях, можно предположить их принадлежность к роду Theodoxus .
В Губском ущелье кроме пещеры Двойной три бусины происходят из верхнепалеолитического слоя Касожской пещеры ( Голованова, Дороничев , 2012. С. 66. Рис. 7: 1 ).
Находки перфорированных раковин в контексте погребений позволяют достоверно реконструировать их применение в качестве нашивок для декорирования одежды, а также использование в виде бус, подвесок, браслетов и т. д. ( Grünberg , 2013. Р. 247; Street et al. , 2002).
Несмотря на значительную по количеству выборку, а также большой географический и хронологический разброс находок бусин, прослеживается определенное сходство в способах перфорирования раковин рода Theodoxus. Морфологические характеристики отверстий аналогичны для бусин, происходящих со стоянок, расположенных в Западной и Восточной Европе, Средиземноморье, Северном Причерноморье, на Кавказе и Ближнем Востоке ( Cristiani et al. , 2014; Pickard et al. , 2017). Важным представляется отсутствие упоминаний о перфорациях, связанных с хищнической деятельностью, аналогичных тем, что зафиксированы на раковинах из пещеры Двойная. По всей видимости, подобные свидетельства крайне редки и/или могут указывать на определенные специфические условия обитания моллюсков, связанные с возможным их сосуществованием с хищными видами. Наличие на нескольких раковинах из пещеры Двойная следов хищнической деятельности неустановленного генезиса раскрывает дальнейшие перспективы для изучения этой категории находок специалистами-малакологами. Остатки предположительно растительных волокон и красящих пигментов на бусинах из раковин требуют более детальных исследований их состава и происхождения ( Александрова и др. , 2014).
Как было показано выше, изготовление бусин из раковин рода Theodoxus было широко распространено на протяжении всего верхнего палеолита, мезолита и неолита Евразии и не может являться каким-либо специфическим культурным или хронологическим маркером. Видимо, яркая окраска и доступность этих моллюсков в ареалах их обитания привлекали внимание первобытных людей, и раковины использовали в качестве украшений. Также в археологических материалах иногда удавалось проследить, что эти ракушки или украшения из них были предметом обмена ( Нужный, Ступак , 2001; Komšo, Vukosavljević , 2011).
Однако для памятников Губского ущелья свидетельства использования раковин моллюсков рода Theodoxus зафиксированы всего в двух случаях – в верхнепалеолитических слоях пещер Двойная и Касожская, материалы которых схожи по облику каменного инвентаря и, вероятно, в археологическом отношении могут считаться синхронными. Для археологических коллекций других, близких по времени, памятников Губского ущелья подобных изделий пока не известно. Отчасти это может быть связано с методикой раскопок прошлых лет, когда не проводились просеивание и промывка культурных отложений и мелкие предметы могли быть не замечены. Но результаты раскопок пещеры Двойная и соседнего навеса Чыгай, где полностью проводилась промывка всех культурных отложений, а также находки в Мезмайской и Касожской пещерах и в навесе Сосруко на данном этапе исследования позволяют предположить, что наличие бусин из раковин речных моллюсков родов Theodoxus и Lithoglyphus в материалах памятников Северо-Западного и Центрального Кавказа является специфической культурно-хронологической чертой населения, обитавшего в этих регионах в конце верхнего палеолита.
Список литературы Находки раковин пресноводных моллюсков в верхнепалеолитическом культурном слое пещеры Двойная (Северо-Западный Кавказ)
- Александрова О. И., Киреева В. Н., Леонова Е. В., 2014. Опыт исследования остатков веществ органического и неорганического происхождения на поверхности каменных орудий из мезолитического слоя пещеры Двойная (Северо-Западный Кавказ) // АЭАЕ. № 4 (60). С. 2-12.
- Александрова О. И., Леонова Е. В., 2017. Реконструкция охотничьего вооружения культур поздней поры верхнего палеолита и мезолита Северного Кавказа (по материалам пещеры Двойная) // SP. No. 1. С. 255-270.
- Анистратенко О. Ю., Старобогатов Я. И., Анистратенко В. В., 1999. Моллюски рода Theodoxus (Gastropoda, Pectinibranchia, Neritidae) Азово-Черноморского бассейна // Вестник зоологии. Т. 33. № 3. С. 11-19.
- Бондарев И. П., 2016. Theodoxus fluviatilis (Gastropoda, Neritidae) как экологический маркер // Морской биологический журнал. Т. 1. № 2. С. 18-26.
- Голованова Л. В., Дороничев В. Б., 2012. Имеретинская культура в верхнем палеолите Кавказа: прошлое и настоящее // Первобытные древности Евразии. К 60-летию А. Н. Сорокина. М.: ИА РАН. С. 59-102.
- Голованова Л. В., Дороничев В. Б., Дороничева Е. В., 2019. Новые данные по палеолиту Приэльбрусья // РА. № 2. С. 7-18.
- Ильина Л. Б., 1987. О следах сверления в раковинах солоноватоводных гастропод // Палеонтологический журнал. № 3. С. 25-33.
- Ильина Л. Б., 2001. О возможном хищничестве неморских гастропод // Палеонтологический журнал. № 6. С. 25-27.
- Кантор Ю. И., Сысоев А. В., 2006. Морские и солоноватоводные брюхоногие моллюски России и сопредельных стран: иллюстрированные каталог. М.: Т-во научных изданий КМК. 371 с., 140 табл.
- Леонова Е. В., 2015. К проблеме хронологии и культурной вариабельности каменных индустрий конца верхнего палеолита и мезолита Северо-Западного Кавказа (по материалам навеса Чыгай и пещеры Двойная) // Традиции и инновации в истории и культуре: программа фундаментальных исследований Президиума РАН. М.: Отделение историко-филологических наук РАН, Ин-т этнологии и антропологии РАН. С. 77-87.
- Нужный Д. Ю., Ступак Д. В., 2001. Поселенческие структуры весенне-летнего периода и сезонная адаптация эпиграветских охотников на мамонта северной Украины // Каменный век европейских равнин: объекты из органических материалов и структура поселений как отражение человеческой культуры: материалы Междунар. конф. (Сергиев Посад, 1-5 июля 1997 г.) / Ред. Т. Н. Манушина. Сергиев Посад: Подкова. С. 51-75.
- Синицын А. А., 2016. Ранний верхний палеолит Восточной Европы: украшения и вопросы эстетики // Верхний палеолит: образы, символы, знаки. Каталог предметов искусства малых форм и уникальных находок верхнего палеолита из археологического собрания МАЭ РАН / Отв. ред. Г. А. Хлопачев. СПб.: Экстрапринт. С. 320-337.
- Яковлева Л. А., 2012. Прикраси з мушельверхнього палеолiту України // Археологiя. Київ: Iнститут археологiї НАН України. № 1. С. 3-12.
- Янина Т. А., 2012. Неоплейстоцен Понто-Каспия: биостратиграфия, палеогеография, корреляция. М.: Географический факультет МГУ. 264 с.
- Álvarez Fernández E., 2006. Los objectos de adorno-colgantes del Paleolítico superior y del Mesolíticoen la Сornisa Cantábrica y en el Valle del Ebro: una vision europea. Salamanca: Universidad de Salamanca. 1333 р. (Сoleccion Vitor; no. 195.)
- André L., Bicho N., 2016. Perforation techniques and traces of use on the Mesolithic adornments of the Trench Area at Cabeço da Amoreira Shellmidden (Muge, central Portugal) // Comptes Rendus Palevol. Vol. 15. Iss. 5. P. 569-580.
- Anistratenko V. V., Zettler M. L., Anistratenko O. Yu., 2017. On the taxonomic relationship between Theodoxus pallasi and T. astrachanicus (Gastropoda: Neritidae) from the Ponto-Caspian region // Archiv für Molluskenkunde. 146. 2. P. 213-226.
- Benghiat S., Komso D., Miracle P. T., 2009. An experimental analysis of perforated shells from the site of Sebrn Abri, Istria, Croatia // Mesolithic Horizons / Eds.: S. McCartan, R. Schulting, G. Warren, P. Woodman. Oxford: Oxbow books.Р. 730-736.
- Boekschoten G. L., 1966. Shell borings of sessile epibiontic organisms as palaeoecological guides (with examples from the Dutch coast) // Palaeogeography, palaeoclimatology, palaeoecology. Vol. 2. P. 333-379.
- Borić D., Cristiani E., 2019. Taking Beads Seriously: Prehistoric Forager Ornamental Traditions in Southeastern Europe // PaleoAnthropology. Special Issue: Personal Ornaments in Early Prehistory / Eds.: D. E. Bar-Yosef Mayer, M. D. Bosch. P. 208-239.
- Cristiani E., Živaljević I., Borić D., 2014. Residue analysis and ornament suspension techniques in prehistory: cyprinid pharyngeal teeth beads from Late Mesolithic burials at Vlasac (Serbia) // JAS. Vol. 46. P. 292-310.
- Glöer P., Pešić V., 2015. The morphological plasticity of Theodoxus fluviatilis (Linnaeus, 1758) (Mollusca: Gastropoda: Neritidae) // Ecologica Montenegrina. Vol. 2. No. 2. P. 88-92.
- Grünberg J., 2013. Animals in Mesolithic burials in Europe // Anthropozoologica. Vol. 48 (2). P. 231-253.
- Harzhauser M., Lenneis E., Neugebauer-Maresch C., 2007. Freshwater gastropods as Neolithic adornment: size selectiveness and perforation morphology as a result of grinding techniques // Annalen des Naturhistorischen Museums in Wien. Bd. 109А. Р. 73-85.
- Hutterer R., Linstädter J., Mikdad A., Mikdad A., 2014. Human manipulation of terrestrial gastropods in Neolithic culture groups of NE Morocco // QI. Vol. 320. P. 83-91.
- Kandel A. W., Bretzke B., Conard N. J., 2018. Epipaleolithic shell beads from Damascus Province, Syria // QI. Vol. 464. Part A. Р. 126-140.
- Komšo D., Vukosavljević N., 2011. Connecting coast and inland: perforated marine and freshwater snail shells in the Croatian Mesolithic // QI. Vol. 244. Iss. 1. Р. 117-125.
- Kowalewski M., 2002. The Fossil Record of Predation: An Overview of Analytical Methods // The Paleontological Society Papers. Vol. 8. P. 3-42.
- Kuhn S. L., Stiner M., Reese D. S., Güleç E., 2001. Ornaments of the earliest Upper Paleolithic: New insights from the Levant // Proceeding of the National Academy of Sciences. Vol. 98. No. 13. Р. 7641-7646.
- Mărgărit M., Radu V., Boroneanț A., Bonsall C., 2018. Experimental studies of personal ornaments from the Iron Gates Mesolithic // Archaeological and Anthropological Sciences. Vol. 10. Iss. 8. P. 2095-2122.
- Pickard C., Boronean A., Bonsall C., 2017. Molluscan remains from early to midden Holocene sites in the Iron Gates reach of the Danube, southeast Europe // Molluscs in Archaeology: Methods, Approaches and Applications (Studying Scientific Archaeology) / Еd. M. J. Allen. Oxford: Oxbow Books. P. 179-194.
- Plotnikov I. S., Ermakhanov Z. K., Aladin N. V., Micklin P., 2016. Modern state of the Small (Northern) Aral Sea fauna // Lakes and Reservoirs: Research and Management. Vol. 21. Iss. 4. P. 315-328.
- Rigaud S., 2013. Les objets de parure associés au dépôtfunérairemésolithique de Große Ofnet: implications pour la compréhension de l'organisation sociale des dernières sociétés de chasseurs-cueilleurs du Jura Souabe // Anthropozoologica. Vol. 48 (2). P. 207-230.
- Sands A. F., Glöer P., Gürlek M. E., Albrecht C., Neubauer T. A., 2020. A revision of the extant species of Theodoxus (Gastropoda, Neritidae) in Asia, with the description of three new species // Zoosystematics and Evolution. Vol. 96 (1). P. 25-66.
- Stiner M. C., 2010. Shell ornaments from the Upper Paleolithic and Mesolithic layers of Klissoura Cave 1 by Prosymnia, Greece // Eurasian Prehistory. Vol. 7 (2). P. 287-308.
- Stiner M. C., Kuhn S., Gulec E., 2013. Early Upper Paleolithic shell beads at Üçağızlı Cave I (Turkey): Technology and the socioeconomic context of ornament life-histories // Journal of Human Evolution. Vol. 64. Iss. 5. P. 380-398.
- Street M., Baales M., Cziesla E., Hartz S., Heinen M., Jöris O., Koch I., Pasda C., Terberger Th., Vollbrecht J., 2002. Final Paleolithic and Mesolithic Research in Reunified Germany // Journal of World Prehistory. Vol. 15. No. 4. Р. 365-453.
- Tátá F., Cascalheira J., Marreiros J., Pereira T., Bicho N., 2014. Shell bead production in the Upper Paleolithic of Vale Boi (SW Portugal): an experimental perspective. // JAS. Vol. 42. P. 29-41.
- Vinarski M. V., Kantor Y. I., 2016. Analytical Catalogue of Fresh and Brackish Water Molluscs of Russia and Adjacent Countries. Moscow: A.N. Severtsov Institute of Ecology and Evolution of RAS. 544 p.
- Yanevich A., Pean S., Crepin L., Laznickova-Galetova M., Prat S., Prysyajnuk V., 2009. Upper Palaeolithic settlements in Buran-Kaya 3 (Crimea, Ukraine): new interdisciplinary researches of the layers 5-2, 6-1 and 6-2 // Археологический альманах. № 20. С. 187-202.
