Накопление макроэлементов морской воды листьями галофитов супралиторали Японского моря

Автор: Воронкова Н.М., Бурковская Е.В., Тимофеева Я.О., Холина А.Б.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 11, 2013 года.
Бесплатный доступ
В статье рассматриваются различные уровни накопления Na, Mg, K, Ca в листьях эугалофитов (Suaeda heteroptera, Salicornia europaea, Salsola komarovii), криногалофитов (Glehnia littoralis) и гликогалофитов (Artemisia stelleriana). Обсуждается их связь с анатомией и мезоструктурой листьев.
Галофиты, накопление элементов, супралитораль
Короткий адрес: https://sciup.org/14082780
IDR: 14082780 | УДК: 581.526.535(571.63)
Accumulation of the sea water macroelements by the halophytes leaves of the Japan sea supralittoral
The various levels of Na, Mg, K, Ca accumulation in the euhalophyte (Suaeda heteroptera, Salicornia europaea, Salsola komarovii), krinohalophyte (Glehnia littoralis and glycohalophyte (Artemisia stelleriana) leaves are considered in the article. Their relationship with the leaf anatomy and mesostructure is discussed.
Текст научной статьи Накопление макроэлементов морской воды листьями галофитов супралиторали Японского моря
Введение . Способность растений морских побережий выживать и успешно размножаться в условиях засоления связана с наличием у них различных адаптаций к высокому уровню ионов Na. В ответ на солевой стресс у соленакапливающих, солевыводящих и солеисключающих галофитов вырабатывается определенная стратегия. Считается, что изучение ответных реакций галофитов как модельных систем способствует выяснению механизмов солеустойчивости [1, 2, 3, 4] и может помочь разработке стратегии улучшения культурных растений-гликогалофитов [5]. С этой точки зрения весьма интересно исследование адаптаций растений морских берегов, особенно супралиторали, где крайне переменчивы не только отдельные экологические факторы, но и весь их комплекс. Специфические условия обитания – засоление почв, импульверизация соленой водой, бедные скелетные почвы, малая мощность снежного покрова, глубокое промерзание почвы, сильные иссушающие ветры, более низкие температуры вегетационного периода, высокая влажность воздуха, периодические тайфуны, антропогенный пресс и другие – оказывают существенное влияние на прибрежную растительность. Высокая динамичность и напряженность природных экологических факторов, а также антропогенные воздействия, позволяют считать морские побережья зоной парадоксальных экологических ситуаций. При анализе флоры морских побережий отмечены черты ксероморфоза многих растений, несмотря на высокую постоянную влажность окружающей среды, обеспеченную муссонным климатом. Кроме того, среди видового разнообразия присутствуют представители семейств, типичных для аридных территорий. На морских берегах есть представители тех же групп галофитов (эу-, крино- и гликогалофиты), что и в аридных зонах [6], но в отличие от последних здесь в основном преобладают представители растений с С 3 -типом фотосинтеза. Растения с С 4 -типом фотосинтеза встречаются значительно реже [7].
В процессе адаптации растения супралиторали вырабатывают особые анатомические, морфологические и физиологические приспособления, позволяющие им выполнять свои жизненно важные функции в присутствии токсических концентраций солей натрия. При этом незаслуженно мало внимания уделяется изучению состава ионов у разных видов галофитов прибрежных районов. Установлено, что ионный состав растений равнинных берегов был стабильным признаком для различных видов галофитов атлантического побережья и не зависел от мозаичной солености [8]. При повышении солености субстрата факультативный галофит Atriplex prostrata резко сокращал фактический фотосинтез, происходило существенное накопление Na, а концентрация K, Ca и Mg уменьшалась [9].
Цель исследований. На данном этапе работы мы исследовали способность к накоплению макроэлементов морской воды представителями разных групп галофитов побережья Японского моря на уровне аккумуляции их в листьях в связи с различной характеристикой мезоструктуры листьев.
Материалы и методы исследований. Сбор растительного материала был проведен в 2010 г. в Южном Приморье в эстуарных зонах на песчано-галечных и заболоченных участках талассосолей залива Угловой и бухты Суходол, входящих в состав зал. Петра Великого Японского моря. Объекты исследований – виды супралиторали эугалофиты Suaeda heteroptera Kitag. , Salicornia europaea L. , Salsola komarovii Iljin, крино-галофит Glehnia littoralis Fr. Schmidt ex Miq. , гликогалофит Artemisia stelleriana Bess. S. Komarovii, которые относятся к растениям с С 4 -типом фотосинтеза, остальные с С 3 -типом. Растения находились в приливноотливных зонах или подвергались в природных условиях импульверизации морской водой. Наружную отмывку солей не проводили.
Для определения содержания катионов основных солей растительный материал предварительно высушивали и измельчали до порошкообразного состояния. Экстракцию элементов из высушенных растительных образцов листьев проводили методом мокрого озоления [10]. Определение элементного состава растений и морской воды выполнено на атомно-эмиссионном спектрометре с индуктивно-связной плазмой Optima 2100 DV (“Perkin Elmer”, США) (РД 52.10.243-92) в центральной лаборатории ОАО “Приморгеология”. Результаты опытов по накоплению катионов в листьях представлены как средние арифметические значения и их стандартные ошибки, полученные в 2 повторностях при анализе 2 усредненных биологических проб, каждая из которых состояла из листьев 10 растений.
Исследование количественных параметров фототрофных тканей листа Salsola komarovii проводили по той же методике, которую использовали при изучении остальных видов, представленных в данной статье, – Suaeda heteroptera, Salicornia europaea, Glehnia littoralis, Artemisia stelleriana [6]. Некоторые мезоструктур-ные характеристики, относящиеся к последним 4 видам, опубликованы ранее [6]. Для удобства изложения материала фотосинтезирующие органы (стилоиды) S. europaea , представляющие сросшийся со стеблем лист, условно именуются листом.
Результаты исследований и их обсуждение. Содержание макроэлементов Na, Mg, K, Ca. В районе исследований морская вода является основным источником макро- и микроэлементов, накапливающихся в растениях как в результате поступления из прибрежных маршевых почв, содержащих те же соли, так и в результате импульверизации и непосредственного контакта во время приливно-отливных явлений. Результат определения указанных макроэлементов в морской воде в районе исследований (залив Угловой) представлен в порядке убывания (мг/л):
Na – 7796 > Mg – 922,5 > Ca – 305,5 > K – 214.
В листьях галофитов преобладающее содержание Na отмечено только для 3 видов ( S. heteroptera , S. komarovii , S. europaea), но с различной интенсивностью накопления (рис.).
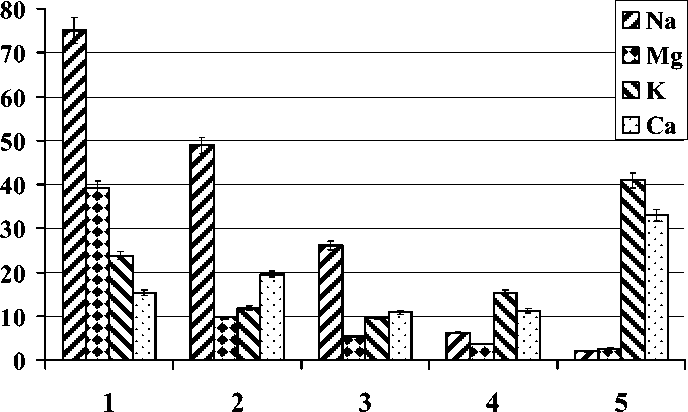
Содержание Na, Mg, K, Ca в листьях галофитов (мг/г сухого вещества):
1 – Suaeda heteroptera; 2 – Salsola komarovii; 3 – Salicornia europaea; 4 – Artemisia stelleriana;
5 – Glehnia littoralis
Индивидуальное содержание остальных макроэлементов у этих растений ниже в 2–5 раз. В отличие от этого, уровень Na и Mg в листьях G. littoralis и A. stelleriana намного ниже в сравнении с уровнем K и Ca, особенно у G. littoralis (рис.). Ранжирование видов по их способности к накоплению отдельных макроэлементов представляет следующие ряды:
для Na – S. heteroptera > S. komarovii > S. europaea > A. stelleriana > G. littoralis ;
Mg – S. heteroptera > S. komarovii > S. europaea > A. stelleriana > G. littoralis ;
К – G. littoralis > S. heteroptera > A. stelleriana > S. komarovii > S. europaea ;
Ca – G. littoralis > S. komarovii > S. heteroptera > A. stelleriana > S. europaea .
Сумма исследуемых элементов в листьях в конце вегетации (мг/г сухого вещества) распределяется в следующем порядке: S. heteroptera (153,3) > S. komarovii (89,9) > G. littoralis (78,5) > S. europaea (52,0) > A. stelleriana (36,3).
Мезоструктура листьев галофитов. Основные показатели мезоструктуры представлены в таблице.
Мезоструктура листа галофитов супралиторали Японского моря
|
Показатель |
Вид |
||||
|
Salsola komarovii |
Suaeda heteroptera |
Salicornia europaea |
Glehnia littoralis |
Artemisia stelleriana |
|
|
Площадь листа, см2 |
0,96±0,39 |
0,66±0,03 |
Нет данных |
28,7±7,5 |
3,45±0,54 |
|
Толщина, мкм: |
|||||
|
мезофилла |
60±6 |
354±34 |
377±33 |
501±15 |
254±14 |
|
водоносной ткани |
1667±67 |
947±45 |
2035±98 |
- |
- |
|
обкладки |
44±6 |
- |
- |
- |
- |
|
Число клеток, тыс/см2: |
|||||
|
столбчатых |
139,5±2,9 |
73,9±4,7 |
262,9±13,8 |
282,2±12 |
148,5±7,3 |
|
губчатых |
- |
40,2±2,8 |
- |
114,1±7,2 |
143,7±7,1 |
|
водоносных |
60,2±2,8 |
30,8±1,9 |
109,1±4,9 |
- |
- |
|
обкладки |
64,3±1,3 |
- |
- |
- |
- |
|
Объем клетки ткани, тыс. мкм: |
|||||
|
столбчатой |
13,3±1,3 |
177,1±17,5* |
25,2±2,3* |
10,5±1,7* |
2,4±0,2* |
|
губчатой |
- |
124,8±9,1* |
- |
3,4±0,4* |
2,3±0,2* |
|
водоносной |
8232,9±613,4 |
2558,3±121,7* |
225,9±12,3* |
- |
- |
|
обкладки |
25,4±2,2 |
- |
- |
- |
- |
|
Индекс поверхности наружных мембран клеток (ИМК) |
|||||
|
ИМК 1 |
120,6 \ |
40,4* 1 |
33,5* |
10,0* |
2,7* |
*Данные опубликованы нами ранее [6] и приведены для сравнительного анализа.
Примечание. Прочерк означает отсутствие этого типа ткани у данного растения.
Виды S. heteroptera , S. komarovii , S. europaea характеризуются наличием водоносной ткани, причем в значительно большем объеме по сравнению с мезофиллом. Те же виды в сравнении с A. stelleriana и G. littoralis имеют меньшую площадь листа, более толстые листья и более крупные клетки. Соответственно индекс поверхности мембран клеток первых трех видов значительно выше.
Удаление ионов и баланс между различными элементами может быть одним из важных механизмов солеустойчивости у галофитов [11, 12]. Опубликованные нами ранее данные по анатомическому строению листьев представителей различных групп приморских галофитов [6] и данные по мезоструктуре листа (табл.) указывают на существование различий в строении листа и накоплении элементов у исследуемых видов (рис.). Так, листья приморских эугалофитов ( S. heteroptera , S. komarovii , S. europaea ) со значительным объемом водоносной ткани (табл.) характеризуются приоритетным накоплением Na по сравнению с другими элементами (рис.). Для эугалофитов супралиторали нет существенной разницы в соотношении накопленных макроэлементов между видами с широким ареалом, растущими как в аридных, так и в гумидных территориях ( S. europaea ) , и видами, растущими только на морском побережье ( S. heteroptera , S. komarovii ).
У всех трех видов преобладает накопление Na. Та же закономерность обнаружена у представителей С3 (S. heteroptera, S. europaea) и С4 (S. komarovii) видов. Однако интенсивность накопления отдельных элементов у них разная (рис.). Больше всего Na обнаружено в эугалофитах, адаптированных к условиям побе- режья (S. heteroptera, S. komarovii), обладающих наибольшим объемом водоносных клеток и характеризующихся наибольшими значениями ИМК, несмотря на меньшее по сравнению с S. europaea число клеток и меньшую толщину водоносной ткани листа (табл.).
Компартментация Na в вакуолях освобождает цитоплазму от токсических ионов, а также обеспечивает осморегуляцию, что считается необходимым для поддержания тургора и роста клеток растяжением [5, 12]. По-видимому, у эугалофитов такой механизм функционирует активно. Предполагают также, что перенос ионов из апопласта в вакуоль в листьях соленакапливающих галофитов, в том числе и S. europaea, может осуществляться, минуя цитоплазму путем пиноцитоза [13]. Представители крино- ( G. littoralis ) и гликогалофитов ( A. stelleriana ), наоборот, накапливают больше K и Ca в сравнении с Na (рис.). Существует мнение, что особенно важным для растений, произрастающих в условиях засоления, является избирательность поглощения K по сравнению с Na [9]. Различное соотношение Na и K в листьях отдельных видов галофитов, вероятно, можно объяснить их конкурентными взаимоотношениями и ингибированием поглощения K при высоких уровнях Na, как было обнаружено в опытах с градиентной соленостью [15], хотя ряд авторов высказывают сомнения в отношении конкуренции [12]). В любом случае избирательность поглощения K перед Na может быть важным адаптивным признаком для представителей крино- и гликогалофитов, характеризующихся отсутствием водоносной ткани и не способных накапливать Na в значительных количествах (рис.).
Для галофитов характерен малый размер листьев (микрофилия), однако механизм, подавляющий рост листьев в условиях солевого стресса, точно неизвестен. Высокая соленость подавляет процессы роста, но виды достаточно близкие могут обладать видоспецифическими экофизиологическими адаптивными особенностями [14]. Накопление K в условиях засоления происходит параллельно скорости роста листа, поэтому замедление роста может быть связано с прогрессирующим снижением уровня K в растущей ткани [15]. Площадь листа G. littoralis значительно выше, чем у эугалофитов (табл.). Ранее мы сообщали о наличии солевыделительных трихом у этого вида , благодаря чему вид обладает активной солевыделительной функцией, что позволило нам отнести его к криногалофитам [6]. Вероятно, повышенный уровень накопления K по сравнению с Na (рис.) и активная функция солевыделения позволяют растениям G. littoralis сохранять сложные пластинчатые листья нормальных размеров и обитать на морских побережьях наряду с соленакопителями (эугалофитами) – растениями высочайшей специализации. В листе гликогалофита A. stelleriana (при наименьшем общем содержании изученных макроэлементов в сравнении с другими видами), как и у криногалофита G. littoralis , K и Ca накапливается больше, чем Na, но уровень K и Ca ниже, чем у последнего (рис.). Кроме солености, ростовые процессы могут регулироваться уровнем азота и аэрации, как было показано в многовариантных экспериментах [16]. Прибрежные маршевые почвы характеризуются высокой субстратной мозаичностью, где уровень азота (песчаные и галечные участки) нередко является лимитирующим фактором.
Анализируя экспериментальные данные, а также полученные ранее результаты [6,17], можно сказать, что адаптивные признаки (анатомические, мезоструктурные) и специфическое соотношение накопления основных элементов, вызывающих засоление (Na, Mg, K и Ca), позволили изученным видам галофитов завершить онтогенез и сформировать жизнеспособные семена, несмотря на высокую соленость среды.
Выводы
-
1. Галофиты супралиторали морских берегов различаются по способности к накоплению в листьях элементов морской воды – Na, Mg, K и Ca.
-
2. Выявлена закономерная зависимость накопления основного засоляющего элемента морской воды Na от принадлежности вида к эу-, глико- и криногалофитам.
-
3. Приоритетное накопление Na свойственно представителям эугалофитов, причем как с С 3 -типом фотосинтеза ( S. heteroptera , S. europaea ), так и с С 4 -типом ( S. komarovii ).
-
4. Представители криногалофитов ( G. littoralis ) и гликогалофитов ( A. stelleriana ) накапливали значительно больше K и Ca, чем Na.
