Накопление тяжелых металлов и активность антиоксидантных ферментов в пшенице при воздействии

Автор: Мурзаева С.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биология и экология
Статья в выпуске: 2 т.4, 2002 года.
Бесплатный доступ
Исследовали накопление тяжёлых металлов (ТМ) в зерне пшеницы при воздействии многокомпонентных загрязнителей (промышленных стоков разной степени очистки). Показано, что степень очистки стоков определяет абсолютное содержание токсических металлов: Zn, Cd, Cu, Cr, Ni, Co, Pb и Mn. Увеличение загрязненности зерна ТМ сопровождалось повышением уровня активностей антоксидантных ферментов: супероксиддисмутазы, каталазы и пероксидазы в листьях и супероксиддисмутазы, пероксидазы в корнях проростков. Обнаружено высокое отношение мембранных форм ферментов к цитозольным. Обсуждается мембранотропное воздействие многокомпонентных загрязнителей, опосредованное ТМ (через образование активных форм кислорода), которые индуцировали антиоксидантную защиту в проростках нового поколения пшеницы.
Короткий адрес: https://sciup.org/148197698
IDR: 148197698 | УДК: 628.581.13.577.7.15
The aссumulation of heavy metals and the activity of antioxidative enzymes in wheat exercise influence of sewages
Investigated accumulation of heavy metals in the wheaten grain exercise influence of multicomponent pollutants (industrial sewages differ by degree of cleaning). It is significant, that the degree of clearing of sewages determines the absolute contents of toxic metals: Zn, Cd, Cu, Cr, Ni, Co, Pb and Mn. The increase polluted of a grain by heavy metals was accompanied by increase of a level activity of antioxidative enzymes: a superoxide dismutase, catalase and peroxidase in leaves and superoxide dismutase, peroxidase in the roots of young growth wheat.s. The high relation of the membranous to cytosoling forms of enzymes is revealed. The membranotropism effect of multicomponent pollutants, through action of heavy metals (through formation of the activity forms oxygen), which one induced the antioxidative protection in young growth wheat of a new generation is discussed.
Текст научной статьи Накопление тяжелых металлов и активность антиоксидантных ферментов в пшенице при воздействии
Открытые системы сброса промышленных и бытовых стоков являются многокомпонентными загрязнителями. Проблема оценки токсичности смесей загрязнителей заключается в малой изученности механизмов аддитивного, синергического и конкурентного взаимодействия составляющих вредных веществ [1]. Разнообразный состав, химическое взаимодействие и динамика изменений присутствующих компонентов усложняет проблему. Особо опасными токсичными веществами в стоках представляются тяжелые металлы (ТМ) [2]. ТМ способны метаболизировать в растениях и накапливаться в их биомассе [3]. По пищевым цепям, через растения, они попадают в организм животных и человека, вызывая терратогенное, мутагенное и канцерогенное действие [4].
В литературе обсуждаются вопросы инициирования тяжелыми металлами свободнорадикальных реакций при контакте с биологическими мембранами, приводящие к образованию активных форм кислорода (АФК) [5] и индуцированию окислительных стрессов [6]. В живых организмах существует многоуровневая система защиты от окислительного стресса, включающая специализированные антиоксидантные ферменты и низкомолекулярные антиоксиданты. Ферментные системы супероксиддисмутазы (СОД), каталазы (КАТ) и пероксидазы (ПО)
составляют первую линию защиты от окислительного стресса, регулируя уровень и устраняя излишки АФК [7]. Степень устойчивости и надежности живых систем, их старение в сильной степени зависят от активности антиоксидантных ферментов [8]. Существуют доказательства экспрессии генов СОД, КАТ и других антиоксидантных белков, индуцированных ТМ и АФК ‒ супероксидом кислорода (О-2) и перекисью водорода (Н2О2) [7, 9, 10]. С этой точки зрения, идентификация мишеней воздействия многокомпонентных загрязнителей по изменению активностей защитных антиоксидантных ферментов представляется актуальной задачей для выявления механизмов трансформации ТМ в растениях.
В настоящей работе исследовали накопление ТМ в пшенице под влиянием промышленных стоков разной степени очистки. Проведена оценка изменения генеративной и ферментативной антиоксидантной функций в проростках в зависимости от уровня загрязненности зерна токсичными ТМ.
Методика
Пшеницу (не сортовую) выращивали в вегетационных ящиках при естественном освещении на открытом воздухе. Поливали све-жеотобранными стоками 1-2 раза в неделю (1 л на 10 см2 при плотности посева семян 1
шт на см2). Контрольные варианты поливали отстоянной водопроводной водой. Использовали сточные воды завода производства минеральных удобрений разной степени очистки: А1 ‒ стоки после биологической очистки, А2 - неочищенные стоки и СМ - условно-чистые, содержащие промышленные и бытовые стоки города. Во избежание постороннего увлажнения, ночью и во время дождя, вегетационные сосуды с растениями накрывали пленкой. После созревания урожая, целые растения с корнем отмывали от почвы, высушивали и обмолачивали зерно. В отдельных частях - колосках без зерна, стеблях с листьями, корнях и молотом зерне определяли тяжелые металлы методом атомноадсорбционной спектроскопии [11].
Семена полученного урожая пшеницы проращивали в темноте, в чашках Петри на фильтровальной бумаге по 100 шт в 3-х повторностях . Увлажняли по мере подсыхания дистиллированной водой. Наклюнувшиеся семена выставляли на свет под люминостат-ные лампы (интенсивность 20 Вт м-2) и выращивали, с периодом освещения 12 ч, при температуре 20-270. Прорастание учитывали по количеству наклюнувшихся семян, биомассу - по весу зеленой массы проростков. В гомогенатах растертых листьев, определяли концентрацию хлорофилла по Вернону [12].
Цитозольные и мембранные белки выделяли из листьев и корней проростков. Навеску растительного материала, 2 г, растирали в ступке с жидким азотом в 20 мл среды, содержащей 50 мМ KH2PO4 и 0,5 мМ этилендиаминтетраацетат (ЭДТА), рН 7,8. Гомогенат листьев или корней фильтровали через четыре слоя капрона и центрифугировали 30 мин при 17000 g. Супернатант использовали для выделения цитозольных, а осадок - для выделения мембранных белков.
Супернатант пропускали через обеззоленный бумажный фильтр, растворимые белки из фильтрата осаждали сульфатом аммония до 80 % насыщения, выдерживая сутки в холодильнике. Высоленные белки отделяли центрифугированием 15 мин при 1300 g, растворяли в минимальном объеме среды, содержащей 0,01 М фосфатный буфер, рН 7,8 и
0,1 мМ ЭДТА, затем обессоливали на колонке с сефадексом g-25, уравновешенной 0,05 М Трис - ацетатным буфером, рН 8,0. В полученном элюате определяли активность растворимых цитозольных ферментов.
Мембранные белки получали из осадка ступенчатым экстрагированием, как описано в работе [13]. Первую экстракцию проводили 1 М NaCl, выдерживая раствор 1 ч при 40, непрерывно перемешивая. Экстракт отделяли центрифугированием и обессоливали на колонке с сефадексом g-25, как описано выше. Элюат содержал от 40 до 70 % мембранных ковалентно связанных белков. Оставшийся осадок обрабатывали охлажденным до -180 абсолютным ацетоном (объемное отношение осадка к ацетону 1:9), выдерживали, помешивая, 1 ч на ледяной бане, затем после осаждения на центрифуге (режим указан выше), промывали охлажденным 80 % ацетоном до полного удаления пигментов. Ацетоновые экстракты, содержащие растворимые пигменты и липиды, отбрасывали. Оставшийся осадок подсушивали под феном и из него экстрагировали липидорастворимые мембранные белки раствором, содержащим 10 мМ фосфатный буфер и 0,1 мМ ЭДТА, рН 7,8, непрерывно помешивая 1 ч при 40. Нерастворимую фракцию отделяли центрифугированием и заливали раствором 100 мМ NaOH на 1 ч при 40 для экстрагирования щелочнорастворимых мембранных белков. В результате последовательных трех экстракций удавалось наиболее полно (до 98 %) извлечь труднорастворимые мембранные белки из растений.
В экстрактах, содержащих цитозольные и мембранные белки, определяли содержание белка по методу Бредфорд [14] и активность антиоксидантных ферментов: суперок-сиддисмутазы (СОД), каталазы (КАТ), пероксидазы (ПО), как описано в работе [15]. За 1 относительную единицу активно сти СОД принято количество фермента, ингибирующее восстановление нитросинего тетразолия на 50 %, на мг белка в пробе. За единицу активности КАТ (или ПО) принято количество фермента, разрушающего 1 мкмоль Н2О2 (или ммоль) в мин на мг белка в пробе.
В работе использовали реактивы отече- ственного производства марки Х.Ч. и О.С.Ч., кроме: НАДН и ЭДТА фирмы "Reanal" (Венгрия), сефадекс g-25 средний и ФМС фирмы "Serva" (Швеция).
Результаты
В пшенице, выращенной при поливе сточными водами А1, А2 и СМ, определяли девять металлов - Fe, Mn, Cu, Zn, Cr, Co, Ni, Pb и Cd . Все эти элементы присутствуют в сточных водах, сбрасываемых в водохранилище [16] . В табл.1 представлено абсолютное содержание исследуемых элементов и их суммы ( ∑ ТМ) в четырех вариантах пшеницы (контроль, А1, А2, СМ) в целых растениях и в отдельных частях - корнях, зерне, соломе (листья + стебель и колос без зерна). Для удобства сравнения с контролем, увеличение содержания тяжелых металлов в опытных вариантах А1, А2 и СМ выделено в таблице затенением ячеек и жирным шрифтом.
Суммарное содержание элементов в целых растениях пшеницы ( ∑ ТМ) соответствует принятой классификации абсолютного содержания металлов в растениях на кг сухой массы [3, 17]: высокие концентрации (выше 100 мг) - Fe, Mn, Zn ; средние (1-100 мг) - Cu,Cr,Ni, Co, Pb и низкие (меньше 1 мг) - Сd, табл. 1. Характер распределения элементов по органам пшеницы акропетальный [3], однако, несмотря на концентрирование ТМ в корнях, они активно переходят в надземные части растений и накапливаются в зерне. Для сравнения показаны ПДК Cu, Zn, Cr, Co, Ni, Pb и Cd , допускаемые ГОСТом в зерне и кормовых [18]. Исходя из норм ПДК, можно сказать, что контрольный вариант зерна пшеницы загрязнен и неудовлетворителен по содержанию Co, Ni, Pb, а соломы - Zn, Cr . Возможно, это было связано с загрязнением почвы, которую использовали для выращивания пшеницы. По данным работы [19] в почвах нашего региона содержание этих металлов, превышает величины кларка.
Сравнивая с контролем, можно видеть, что сточные воды избирательно способствовали накоплению элементов в пшенице (целое растение). Воды А1 снижали на 27-47% содержание Cd, Pb, Co, незначительно уве- личивали - Fe, Mn, Cr, Ni. Общий уровень тяжелых металлов, ∑ТМ, под действием этих стоков был повышен только на 5 %.
Воды СМ увеличивали накопление Cd, Ni, Mn и Cu на 6-14 %, концентрации остальных металлов - уменьшали, особенно, Fe и Co (на 22 и 42 %), в итоге сумма элементов была уменьшена на 20 %.
Воды А2 способствовали накоплению всех металлов, кроме Co . Значительно увеличивали содержание Fe и Cr (на 71 и 101%). Общий уровень металлов в растениях возрастал на 67 %. Причиной тому, по-видимому, была большая загрязненность неочищенных стоков, по сравнению с двумя другими, которые частично очищаются [16 с.140].
По суммарному накоплению ТМ целыми растениями, исследуемые варианты пшеницы представляют ряд: А2 > А1 > К > СМ, что показывает зависимость действия стоков от очистки и положительное влияние условно-чистых стоков СМ. Такой же ряд соответствует загрязнению ТМ корней. Однако, при распределении металлов по другим органам пшеницы ряд накопления ∑ ТМ варьирует: в соломе - А2> А1> СМ > К, а в зерне - СМ > К > А2> А1.
Относительно накопления ТМ зерном, следует отметить, что все стоки способствовали активному транспорту из корней в зерно Zn и Mn . Стоки СМ оказывали заметное действие по продвижению Cr и Fe из корней в зерно. В данном случае превышение по железу в зерне составляло в 1,5 раза, а по хрому в 10 раз по сравнению с контролем , поэтому приоритетными загрязнителями зерна по ∑ ТМ были стоки СМ.
Оценивая фитотоксичность пшеницы в целом, отметим, что в основном, ТМ аккумулируется в корнях и, следовательно, остаются в почве. Определенная часть металлов трансформируется по растению и, в зависимости от метаболизма фотосинтезирующими органами, попадает в стебли, листья и зерно. В итоге, солома (кормовой продукт для скота) обогащена не менее чем 5-7, а зерно - 3-4 аккумулированными элементами. Среди них высоко токсичные - Zn, Cd и токсичные - Cu, Cr, Ni, Co, Pb, Mn (классификация токсично-
Таблица 1. Содержание металлов в пшенице, мг на кг сухой массы
|
Вариант пшеницы |
Органы пшеницы |
Ее |
Мп |
Си |
Zn |
Ст |
Со |
м |
РЬ |
Cd |
Ztm |
|
Контроль |
корни |
4550.8 |
117.3 |
10.4 |
44.8 |
6.1 |
2.1 |
9.9 |
2,63 |
0.13 |
4744.16 |
|
*солома |
322.3 |
76.9 |
7.2 |
52.9 |
0.74 |
1.7 |
2.5 |
0.95 |
0.23 |
465,42 |
|
|
зерно |
35.6 |
44,7 |
4,3 |
39.5 |
0,07 |
2 Л |
2.2 |
0.71 |
0Л2 |
129.3 |
|
|
целое растение |
4908.7 |
238.9 |
21,9 |
137,2 |
6.91 |
5.9 |
14.6 |
4.29 |
0,48 |
5338.88 |
|
|
Стоки А] |
корни |
4773,3 |
117.8 |
10.7 |
34.8 |
6,26 |
2.0 |
10,1 |
2,21 |
0.14 |
495731 |
|
солома |
415,8 |
82,3 |
7.0 |
46.9 |
1,08 |
1.1 |
3,6 |
0.69 |
0.1 |
558,57 |
|
|
зерно |
11,3 |
48,4 |
3,1 |
47,6 |
н/о |
н/о |
2,5 |
н/о |
0.11 |
113.01 |
|
|
целое растение |
5200,4 |
248,5 |
20.8 |
129.3 |
7,34 |
ЗД |
16,2 |
2.9 |
0.35 |
5628,99 |
|
|
Стоки А2 |
корни |
7800,4 |
139,7 |
13.2 |
42,2 |
13,02 |
3,4 |
13,7 |
2,95 |
0,14 |
8028,71 |
|
солома |
606,9 |
101,4 |
6.6 |
52.0 |
0,89 |
2,3 |
3,8 |
2,62 |
0.21 |
776,72 |
|
|
зерно |
9.7 |
54,0 |
4.4 |
49,5 |
н/о |
н/о |
2,5 |
0.54 |
0,27 |
120.91 |
|
|
целое растение |
8417,0 |
295,1 |
24,2 |
143,7 |
13,91 |
5,7 |
20,0 |
6,11 |
0,62 |
892634 |
|
|
Стоки СМ |
корни |
3433.0 |
132,8 |
11,8 |
33.9 |
5.6 |
1.0 |
9.1 |
1,69 |
0,21 |
3629,1 |
|
солома |
333,3 |
84,5 |
8,0 |
42,3 |
0,92 |
1.7 |
43 |
2,17 |
0.19 |
477,38 |
|
|
зерно |
53,0 |
55,2 |
3.6 |
52,5 |
0,2 |
0.7 |
2.2 |
н/о |
0.11 |
167,51 |
|
|
целое растение |
3819.3 |
272,5 |
23,4 |
128.7 |
6.72 |
3.4 |
15,6 |
3.86 |
0,51 |
4273.99 |
|
|
ГОСТ |
ПДК для зерна. |
- |
— |
10 |
50 |
0.35 |
0.06 |
0.7 |
0.5 |
0.1 |
|
|
1986 г |
|||||||||||
|
для кормовых |
- |
- |
30 |
50 |
0,5 |
- |
- |
3.0 |
0.5 |
Биология и экология
Таблица 2. Влияние стоков на генеративную функцию семян пшеницы
|
Вариант |
Проросло, % |
Зеленая часть |
|||
|
Рост, см |
Биомасса растений, 100 г |
Хлорофилл, мкг на мг с |
∗ Белок, ырой массы |
||
|
Контроль |
97 |
5 |
4,4 |
0,95 |
32 |
|
А 1 |
98 |
9 |
7,0 |
0,95 |
34 |
|
А 2 |
99 |
7 |
5,6 |
0,9 |
29 |
|
СМ |
98 |
8 |
5,4 |
1,0 |
37 |
Примечание: * - общий белок (мембранные + цитозольные белки). Возраст проростков 9 суток, освещение 20 Вт на м2. Приведены средние данные трех биологических опытов, хлорофилл и белок определяли в каждом опыте в двух аналитических повторностях - отклонение от среднего не превышало ± 5 %.
сти элементов по ГОСТу 1983 [20]). Качественный состав стоков и степень их очистки определяли абсолютное содержание токсикантов в органах пшеницы. Так, стоки А 1 , прошедшие биологическую очистку аккумулировали в зерне Mn, Zn и Ni , стоки А2 - Mn, Zn, Ni и Cd , а стоки СМ - Mn, Zn и Cr . Содержание Ni в зерне соответствующих вариантов превышало ПДК в 3,6 , Cd - в 2,7 , а Zn -1,05 раза. В то же время стоки А 1 и СМ ограничивали поступление в репродуктивные органы пшеницы Cd, Pb и Cu . В итоге ряд загрязнения зерна токсичными ТМ соответствует степени очистки стоков: А 2 > СМ > А 1 > К и отличается от такового по суммарному накоплению ТМ зерном, где на первое место выходили стоки СМ и контроль, а затем уже А 2 и А
Пролонгирующее действие ТМ на генеративную функцию полученных семян показано в табл.2. Оказалось, что семена всех четырех вариантов пшеницы при проращивании имели одинаковую энергию прорастания. По скорости роста и наращиванию зеленой биомассы все опытные варианты проростков опережали контроль. Наибольшие показатели по биомассе имели проростки А Впрочем, такие же результаты были получены нами при проращивании "родительских" с емян в присутствии соответствующих стоков [21]. Следовательно, те изменения, которые произошли на уровне родительских форм под воздействием стоков проявились при генерации нового поколения, но каких-либо новых морфологических отклонений обнаружено не было.
Проростки практически не различались по биохимическим показателям - содержанию хлорофилла и белка. Незначительные изменения, которые наблюдали, скорее можно расценивать как тенденцию негативного или положительного влияния соответствующих стоков на биохимические синтезы белков и пигментов в пшенице.
Главные и заметные изменения на биохимическом уровне обнаружены в антиоксидантной защите проро стков. Они проявлялись в изменении активностей ферментов СОД, КАТ и ПО. Эти ферменты присутствуют в растительной клетке в мембранах и цитозоле [13, 15,22]. На рис.1,2, 3 представлены активности мембранных и цитозольных форм ферментов в листьях и корнях исследу-
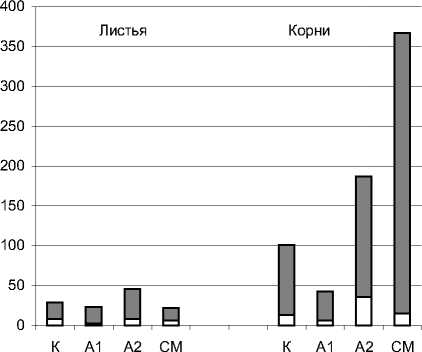
цитозоль мембраны
Рис. 1. Активность супероксиддисмутазы в проростках пшеницы
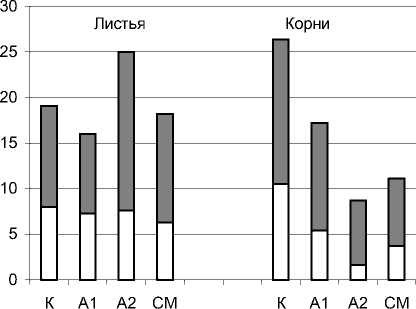
цитозоль мембраны
Рис. 2. Активность каталазы в проростках пшеницы
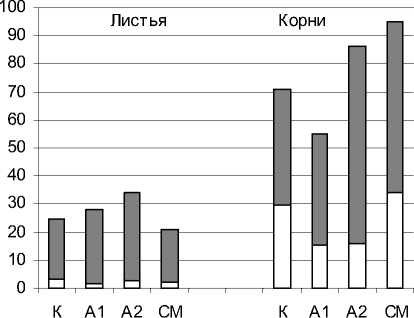
цитозоль мембраны
Рис. 3. Активность пероксидазы в проростках пшеницы
емых вариантов проростков пшеницы.
На рис.1. показаны изменения активности СОД. Характерным признаком была буль-шая активность СОД в корнях, по сравнению с листьями (в 2-16 раз) и преобладание активности мембранной формы фермента над цитозольной. В листьях отношение мембранная/цитозольная СОД варьировало в пределе 2,5-9,2 раза, в корнях - 4,2-22,8. Сравнивая с контролем, видно, что стоки А 1 повлияли на уменьшение активности фермента в листьях и корнях в 1,3 и 2,3 раза, а А2 - на увеличение - в 1,6 и 1,9 раза, соответственно. Воды СМ уменьшали активность СОД в листьях в 1,3, но увеличивали - в корнях в 3 раза. Увеличения активности связаны, в основном, с мембранной формой фермента.
В отличие от СОД, каталазная активность была выше в листьях, а не в корнях, за исключением контроля, и отмечалось наличие большей доли цитозольной формы фермента, рис.2. Отношение мембранная/цито-зольная КАТ было меньше чем для СОД (варьировало в пределе 1,2-2,3), но преобладала также мембранная форма. Увеличение активности КАТ в 1,3 раза происходило только в листьях проростков А 2 и оно было связано, в основном, с мембранной формой. В о стальных случаях наблюдали уменьшение активности КАТ - незначительное в листьях А 1 (в 1,2 раз) и более сильное в корнях
А1, А2 и СМ (в 1,5 - 3 раза).
Общая активность ПО в проростках, также как и СОД, была выше в корнях (в 2-5 раз), по сравнению с листьями, рис.3. Подобно СОД в листьях присутствовала, в основном, мембранная форма фермента и следы цитозольной. В корнях обнаруживали обе формы пероксидазы с преобладанием в 1,4 - 2,6 раза мембранной над цитозольной. Стоки А1 незначительно повышали активность ПО в листьях (в 1,1), А2 - в листьях и корнях (в 1,4 и 1,2, соответственно), а СМ - только в корнях (в 1,3 раза). Стоки СМ способствовали снижению активности фермента в листьях (в 1,2), а стоки А 1 - в корнях (в 1,3 раза).
Динамика изменения активностей ферментов в зависимости от стоков, в процентном отношении к контролю, показана в табл.3. Видно, что колебания активности СОД в про-ро стках происходили в пределе от -58 до +265, КАТ - от -67 до +30, а ПО - от -23 до +40 %. Самые большие изменения в активности под влиянием стоков происходили с СОД, особенно в корнях. Ферменты КАТ и ПО испытывали меньшие изменения. Следует также отметить соответствие увеличения или уменьшения активностей всех трех ферментов СОД, КАТ, ПО в листьях и пары СОД, ПО в корнях в зависимости от уровня, присутствующих в зерне токсичных ТМ: А2 > СМ > А 1 .
Таблица 3. Динамика изменений активностей антиоксидантных ферментов в проростках пшеницы в зависимости от стоков, %
|
Вариант |
Листья |
Корни |
||||
|
СОД |
КАТ |
ПО |
СОД |
КАТ |
ПО |
|
|
А 1 |
-23 |
-16 |
+10 |
-58 |
-35 |
-23 |
|
А 2 |
+60 |
+30 |
+40 |
+87 |
-67 |
+20 |
|
СМ |
-23 |
-5 |
-15 |
+265 |
-58 |
+30 |
± - увеличение или уменьшение по отношению к контролю.
Обсуждение результатов
Данные, представленные в настоящей работе, показывают, что в присутствии сточных вод пшеница накапливает ТМ. Характер количественных изменений распределения отдельных элементов по органам пшеницы, свидетельствуют о сложных механизмах трансформации загрязнителей в растениях. Возможно, действующим началом поглощения растениями элементов являются неорганические и органические комплексы металлов, содержащиеся в стоках и образующиеся при поступлении в органы растений. Они могли конкурировать между собой и составлять конкуренцию подвижным формам элементов используемой почвы [23-25], что можно проследить по Cd .
Подвижный элемент Cd, проявляющий повышенную мобильность в растениях [4], обнаруживался в значительных количествах во всех органах контрольного варианта пшеницы. Биологически очищенные стоки А1 снижали, а неочищенные стоки А2 и условно-чистые СМ способствовали увеличению этого элемента в целых растениях пшеницы, по сравнению с контролем. Причем, стоки СМ задерживали кадмий в корнях, а стоки А2, напротив, способствовали трансформации токсиканта по растению и накоплению в семенах. По-видимому, в стоках СМ содержались компоненты, способные связывать кадмий и ограничивать его поступление в фотосинтезирующие, а затем в репродуктивные органы растений. Это могли быть основания, сдвигающие рН в щелочную сторону и способствующие осаждению Ме [3, 18, 23]. В действительности, измерение рН стоков во время наших экспериментов показывали более щелочной показатель у СМ (рН 8-9), по сравнению со стоками А1 и А2 (рН 6-8).
Не следует исключать и конкуренцию между Ме или, образуемыми органическими комплексами с катионами металлов, стабильность которых определяется природой металла [23, 25]. Стоки СМ и А1 задерживали в корнях и листьях не только Cd , но и Cu, Pb , Mn , что уменьшало их накопление в зерне. Следовательно, репродуктивные органы пшеницы были защищены от наиболее активных ТМ, легко реагирующих с физиологически важными веществами. Cd и Pb - вызывают мутагенез [3, 4], а Cu индуцирует оксидазные системы в живых организмах [25]. В итоге, фитотоксиность зерна по загрязнению токсичными ТМ была снижена, по сравнению со стоками А2.
В работе [21] нами было показано, что во время вегетации пшеница в присутствии стоков А2 быстрее проходила все стадии развития, но отставала в росте и имела более ломкий стебель. Кроме того, эти стоки увеличивали долю уродливых растений. Вероятно, это было следствие канцерогенного действия Cd [3,4], которое, как показали данные в настоящей работе, не проявилось при проращивании новых семян. Увеличение в два раза количества этого элемента в зерне варианта А2, не вызывало каких-либо внешних морфологических нарушений в эмбриогенезе. Энергия прорастания и биологическая продуктивнсть (биомасса) проростков увеличивались (см. табл.2) и были сходны тем, что наблюдали у родительских форм [21]. Данные показывают, что при явном отсутствии внешних морфологических нарушений, в пшенице нового поколения на ранней стадии развития происходили изменения на биохимическом уровне - изменялись активнос- ти антиоксидантных ферментов: СОД, КАТ, ПО. Под действием стоков отмечается увеличение доли мембранных форм ферментов по отношению к цитозольным. Этот факт указывает на мембранотропное последействие стоков.
Увеличение загрязненности зерна токсичными ТМ приводит к повышению уровня антиоксидантной защиты в проростках. Это прослеживается по возрастанию активности трех ферментов СОД-КАТ-ПО в листьях у варианта А2 или пары - СОД-ПО в корнях вариантов А2 и СМ, см. рис.1-3, родительские растения которых имели больший уровень накопления ТМ в соответствующих органах и зерне см табл.1. Напротив, у пшеницы подверженной действию менее загрязненных сточных вод А1 (которые, в дополнение, уменьшали накопление токсичных металлов в зерне), в проростках обнаруживали уменьшение активностей СОД-КАТ-ПО в корнях и СОД-КАТ в листьях.
Функция ферментов СОД, КАТ и ПО связана с утилизацией активных форм кислорода [5]. СОД осуществляет дисмутацию супероксидного радикала (О2-.) до менее токсичной перекиси водорода [5], а Н2О2 удаляется каталазой и пероксидазой разными путями [22, 26]. КАТ разрушает избыток образуемой перекиси водорода, а ПО использует Н2О2 для окисления широкого спектра веществ, в том числе при детоксикации чужеродных соединений [26, 27]. При оксидазном механизме пероксидазы при определённых условиях могут продуцироваться О2-. и, более токсичный гидроксил радикал (ОН.) [28]. Эти формы АФК инактивируют СОД и, особенно, КАТ [29]. Есть сообщения, что и СОД может продуцировать ОН. [25, 29, 30]. Таким образом, СОД и ПО, кроме антиоксидантного, проявляют еще и прооксидативное свойство, что может способствовать развитию окислительных реакций и инициированию окислительного стресса.
Экспериментально, в работе [15], нами было показано, что развитие окислительного стресса в растениях при токсических дозах ксенобиотика сопровождается гиперусилением пероксидазной активности, в 10 и более раз, на фоне увеличения СОД в 2-3 раза. Растения при этом погибали. В настоящей работе морфологических нарушений в проростках не наблюдали, следовательно окислительный стресс на ранней стадии развития растения не испытывали. Однако индуцирование пары ферментов СОД-ПО и уменьшение каталазной активности свидетельствуют об активно протекающих окислительных реакциях в корнях проростков. Известно, что ферменты СОД и ПО участвуют в детоксикации ксенобиотиков [26, 30]. Возможно, увеличение окислительной активности связано с продолжающимися реакциями инактивирования ТМ, осажденных в виде каких либо хелатов и комплексов в мембранах клеток корней, через окислительные реакции и образование неактивных лигандов [25]. В любом случае это следует расценивать как индукцию прооксидативного действия ферментов СОД-ПО в растительных клетках тяжелыми металлами и возможность усиления окислительной активности в растениях при дальнейшем развитии.
Итак, заключая полученные данные, отметим следующее.
-
- Многокомпонентные загрязнители -сточные воды, способствуют накоплению ТМ растениями пшеницы, адсорбируя их в зерне.
-
- В проростках пшеницы семян 1-го поколения индуцируются активности антиоксидантных ферментов СОД, КАТ и ПО с высоким отношением мебранные/цитозольные формы.
-
- Уровень и качественный состав ТМ в зерне определяют мембранотропный уровень антиоксидантной защиты - СОД-КАТ-ПО в новом поколении на ранней стадии развития проростков.
-
- Увеличение активностей СОД ‒ ПО, на фоне уменьшения каталазной активности, показывают возможность прооксидативного действия антиоксидантных ферментов при токсическом воздействии ТМ.
Остается вопрос о механизме индукции антиоксидантной защиты. Был ли это синтез ферментов de novo, вызванный экспрессией генов в родительских формах растений и пе- реданный новому поколению, или это активация ферментов, связанная с защитой мембран от ТМ, накопленных зерном и трансформируемых в проростках? Оба эти механизма могут иметь место. Из литературы известно, что индукторами экспрессии генов белков, защищающих от окислительного и других стрессов, являются факторы, вызывающие соответствующий стресс, в том числе АФК (О-.2, Н2О2) и ТМ [7, 9, 10]. На наш взгляд, некоторым подтверждением синтеза ферментов de novo, по крайней мере СОД, служит сильная активация этого фермента по сравнению с КАТ и ПО. Если такое происходило у родительских форм, то продуцируемая СОД перекись водорода, не успевая разрушаться, свободно проникала в цитозоль к ядерному матриксу и, как сигнальная молекула, могла индуцировать соответствующие участки в генах [10]. Вызывает интерес закрепления этих признаков - индуцирования антиоксидантной защиты, далее в поколениях пшеницы, что безусловно способствует формированию устойчивости растений к загрязнению окружающей среды ТМ. Эти вопросы является предметом наших дальнейших исследований.
