Население птиц антропогенных местообитаний поймы Нижней Оби при разном уровне обводнённости

Автор: Пасхальный Сергей Петрович, Головатин Михаил Григорьевич
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 572 т.19, 2010 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140151793
IDR: 140151793
Bird population of disturbed habitats in flood-plain of Lower Ob under different levels of water regime
Текст статьи Население птиц антропогенных местообитаний поймы Нижней Оби при разном уровне обводнённости
-
2) Институт экологии растений и животных Уральского отделения Российской Академии наук, ул. 8 Марта, 202, Екатеринбург, 620144, Россия. E-mail: golovatin@ipae.uran.ru
Поступила в редакцию 16 апреля 2010
Антропогенные местообитания в пойме Оби в настоящее время занимают относительно небольшую площадь. Инженерные сложности размещения здесь каких-либо строений, большая вероятность их затопления во время паводка и другие технические проблемы пока сдерживают расширение техногенных площадей. Однако дальнейшее освоение региона, скорее всего, приведёт к расширению здесь человеческой деятельности. Поэтому изучение фауны нарушенной территории поймы Нижней Оби становится актуальным.
Хорошо известно, что в поймах население птиц в сильной степени зависит от уровня и продолжительности половодья. Обская пойма в этом отношении особо интересна, поскольку из-за небольших перепадов высот значительные площади поймы Оби в некоторые годы остаются затопленными практически всё лето. В маловодные же годы, когда весенний паводок не достигает максимальных уровней, уровень воды довольно быстро падает. Понятно, что в столь разных условиях состав, численность и пространственное распределение птиц могут существенно различаться. В настоящей работе мы анализируем эти аспекты населения птиц окрестностей города Лабытнанги – одного из самых измененных человеком участков поймы Оби.
Район работ
Исследования проводили в пойменной части на периферии города Лабытнанги (66 ° 40' с.ш., 66 ° 30' в.д.). Техногенная территория простирается здесь вдоль 6-км насыпной дамбы с железной и автомобильной дорогами от протоки Вылпосл, протекающей у левого коренного берега реки, до русла Большой Оби, где расположен речной порт. На большей своей части дамба проходит вдоль озёр и стариц. Наши работы были сосредоточены на двух участках, прилегающих к дамбе: возле протоки Вылпосл и у Лабытнангского порта на Оби. Площадь и конфигурация водоёмов (или затопленных территорий) в разных условиях обводнённости здесь существенно различаются.
На площадке № 1 на правом берегу протоки Вылпосл ранее находилось предприятие по трелёвке и разделке древесины из приводимых сюда плотов. Сейчас этот участок не используется и представляет собой пустырь, засыпанный древесными отходами и разным мусором. С противоположной стороны эта территория прилегает к заливным осоковым лугам и мелким озёрам, высокорослым древовидным и парковым ивнякам, а далее ограничена старицей. Местами заросли ивняков более густые, но низкорослые, местами обычного паркового типа. С противоположной стороны дамбы размещается действующая торговая база, пустыри и свалка мусора. С запада и востока эта площадка граничит с соровыми озёрами, с юга – с заливными лугами с разреженным низкорослым ивняком.
На площадке № 2 (у порта) проложено несколько насыпей с дорогами, имеются действующие и заброшенные производственные сооружения, участки складирования материалов. На территории порта и рядом с ним есть несколько озёр и стариц. Участки между водоёмами и собственно антропогенными территориями заняты либо заливными осоковыми лугами и болотами, чаще небольшими по площади, либо зарослями древовидных ивняков, местами очень густых, кое-где разреженных. На границах с соровыми озёрами высота и густота кустарниковых зарослей обычно ниже.
Материал и методика
В работе представлены материалы разных по водности лет – 2007 и 2008. В 2007 году наблюдения были начаты в мае и закончены в октябре. Основные учётные мероприятия пришлись на июль и август. В 2008 году работы также начались в мае, учёты выполняли в конце июня – начале июля.
Учеты птиц проводили двумя методами: абсолютный учёт гнездящихся пар (для домового воробья – особей) на площади и относительный – оценка числа особей каждого вида на 1 км маршрута. В 2007 году данные для абсолютного учёта использованы только за июль, для относительного – за июль и август, с тем, чтобы сравнить изменение численности птиц в течение двух месяцев лета. Для рано гнездящихся птиц (сорока, серая ворона, чечётка) и видов-посетителей абсолютную плотность не определяли. Размер и конфигурация площадок, а также прокладка маршрутов в значительной мере определялись тем, насколько возможно было передвижение при имеющемся в тот момент уровне воды. В 2007 году значительные участки поймы из-за затопления оказались просто непроходимыми. С другой стороны, они и не использовались для гнездования большинством видов птиц. В 2008 году удалось заложить маршруты несколько шире, но в пределах тех же площадок. Размер учётных площадок и протяжённость маршрутов на них представлены в таблице 1.
Ошибку при оценке плотности гнездования определяли по формуле: m = (√ N ) / S , где N – число гнездящихся пар, S – учётная площадь (Смирнов 1964, 1965). Значимость различий в плотности гнездования видов и таксономических групп в разные годы оценивали по t -критерию (Песенко 1982).
Таблица 1. Размер учётных площадок и протяжённость маршрутов
|
Год |
Учетная площадь, км2 |
Период |
Протяженность маршрутов, км |
|
|
Площадка № 1 |
Площадка № 2 |
|||
|
2007 |
1.7 |
1.8 |
Июль |
17.6 |
|
1.7 |
1.8 |
Август |
11.5 |
|
|
2008 |
1.7 |
1.8 |
Июнь-июль |
15.5 |
Особенности летних сезонов 2007-2008 годов
Лето 2007 года было тёплым, хотя весна, включая июнь, выдалась затяжной и холодной. Время от времени случались сильные ветры и обильные осадки. Ледоход на Оби отмечен 20 мая. В районе наблюдений уровень паводковых вод оказался высоким, позднее, во второй половине июля, он ещё повысился и примерно на таком уровне продержался до середины сентября. В результате большие площади в пойме оказались затоплены на длительное время. Этому способствовало и большое количество осадков летом. Участки высокорослых и парковых ивняков стояли в воде, она заливала даже насыпные дороги. Как следствие, резко сократилось число мест, пригодных для устройства гнёзд для наземногнездящихся видов, особенно мелких древесно-кустарниковых воробьиных, куликов и уток. Сократилась также площадь наземных мест кормёжки.
В 2008 году ледоход на основном русле Оби наблюдался 27-28 мая при низком уровне паводка. Из-за маловодности большинство пойменных озёр до конца мая так и не освободились ото льда. Максимальный подъём воды был примерно на 11.5 м ниже уровня 2007 года. Паводок оказался непродолжительным – уже к 6 июля затопленные участки поймы стали осушаться. Первая половина июня выдалась прохладной, хотя среднемесячная температура воздуха была на 0.7° выше средних многолетних значений, июля – выше на 1.4°. Экстремальных погодных явлений не отмечено. Дожди были слабыми и кратковременными.
Видовой состав населения птиц
Всего в летнее время на исследуемой территории отмечено 45 видов птиц (табл. 2). Из них достоверно гнездились в этом районе 35 видов. Возможно гнездование ещё 3 видов птиц (статус не ясен), остальные только посещают этот район поймы. Ряд воробьиных (луговой и краснозобый коньки, жёлтая трясогузка и некоторые другие) становятся обычными в пойме лишь с началом послегнездовых кочёвок. В период пролёта видовой состав птиц значительно расширяется как за счёт птиц, населяющих ненарушенные местообитания поймы, так и за счёт мигрантов, гнездящихся севернее или в этом же районе, но за пределами поймы. Важно отметить, что мы говорим о составе летней орнитофауны в том районе поймы, который подвержен существенному антропогенному влиянию.
Численность
Об общем уровне численности птиц в период наших работ можно сказать следующее. В 2007 году многочисленными (≥ 10 пар/км2) были 4 вида: озёрная и малая чайки, речная крачка и береговая ласточка, образовывавшие колонии на учётных площадках (табл. 3). В 2008 году в числе многочисленных оказались только малая чайка и береговая ласточка. Относительно низкие значения плотности связаны с большими площадями местообитаний, где гнездование птиц невозможно (водоёмы, дороги с интенсивным движением транспорта, места постоянного присутствия людей и т.п.).
Таблица 2. Список видов птиц, отмеченных летом 2007-2008 годов на исследуемой территории поймы Оби
|
Вид |
Статус |
Численность |
|
|
Красношейная поганка |
Podiceps auritus |
Гнездящийся |
Редкий |
|
Чирок-свистунок |
Anas crecca |
Гнездящийся |
Редкий |
|
Свиязь |
Anas penelope |
Гнездящийся |
Немногочисленный |
|
или обычный |
|||
|
Шилохвость |
Anas acuta |
Гнездящийся |
Немногочисленный |
|
Широконоска |
Anas clypeata |
Гнездящийся |
Редкий |
|
Хохлатая чернеть |
Aythya fuligula |
Гнездящийся |
Обычный |
|
Синьга |
Melanitta nigra |
Не ясно |
Редкий |
|
Дербник |
Falco columbarius |
Посетитель |
Редкий |
|
Галстучник |
Charadrius hiaticula |
Гнездящийся |
Немногочисленный |
|
Малый зуёк |
Charadrius dubius |
Гнездящийся |
Немногочисленный |
|
Фифи |
Tringa glareola |
Гнездящийся |
Немногочисленный |
|
Большой улит |
Tringa nebularia |
Гнездящийся |
Редкий |
|
Мородунка |
Xenus cinereus |
Гнездящийся |
Обычный |
|
Турухтан |
Philomachus pugnax |
Не ясно |
Редкий |
|
Белохвостый песочник |
Calidris temminckii |
Гнездящийся |
Немногочисленный |
|
Бекас |
Gallinago gallinago |
Гнездящийся |
Обычный |
|
Средний кроншнеп |
Numenius phaeopus |
Посетитель |
Редкий |
|
Малый веретенник |
Limosa lapponica |
Посетитель |
Редкий |
|
Малая чайка |
Larus minutus |
Гнездящийся |
Многочисленный |
|
Озерная чайка |
Larus ridibundus |
Гнездящийся |
Многочисленный |
|
Восточная клуша |
Larus heuglini |
Не ясно |
Редкий |
|
Сизая чайка |
Larus canus |
Гнездящийся |
Немногочисленный |
|
Речная крачка |
Sterna hirundo |
Гнездящийся |
Многочисленный |
|
Береговая ласточка |
Riparia riparia |
Гнездящийся |
Обычный |
|
Луговой конёк |
Anthus pratensis |
Посетитель |
В августе обычный |
|
Краснозобый конёк |
Anthus cervinus |
Гнездящийся |
Редкий, в августе обычный |
|
Жёлтая трясогузка |
Motacilla flava |
Гнездящийся |
Редкий, в августе обычный |
|
Белая трясогузка |
Motacilla alba |
Гнездящийся |
Обычный |
|
Сорока |
Pica pica |
Гнездящийся |
Обычный |
|
Серая ворона |
Corvus cornix |
Гнездящийся |
Обычный |
|
Ворон |
Corvus corax |
Посетитель |
Редкий |
|
Свиристель |
Bombycilla garrulus |
Посетитель |
Редкий |
|
Камышевка-барсучок |
Acrocephalus schoenobaenus |
Гнездящийся |
Обычный |
|
Пеночка-весничка |
Phylloscopus trochilus |
Гнездящийся |
Обычный |
|
Пеночка-таловка |
Phylloscopus borealis |
Гнездящийся |
Обычный |
|
Пеночка-теньковка |
Phylloscopus collybita |
Гнездящийся |
Немногочисленный |
|
Каменка |
Oenanthe oenanthe |
Гнездящийся |
Редкий |
|
Варакушка |
Luscinia svecica |
Гнездящийся |
Обычный |
|
Рябинник |
Turdus pilaris |
Гнездящийся |
Обычный |
|
Белобровик |
Turdus iliacus |
Гнездящийся |
Немногочисленный |
|
Домовый воробей |
Passer domesticus |
Гнездящийся |
Обычный |
|
Юрок |
Fringilla montifringilla |
Гнездящийся |
Обычный |
|
Обыкновенная чечётка |
Acanthis flammea |
Гнездящийся |
Обычный |
|
Тростниковая овсянка |
Emberiza schoeniclus |
Гнездящийся |
Обычный |
|
Овсянка-крошка |
Emberiza pusilla |
Гнездящийся |
Обычный |
Таблица 3. Плотность гнездования (пар/км2) и относительная численность (особей/км) птиц антропогенной поймы Нижней Оби в 2007-2008 годах
|
Вид |
Пар |
Плотность M ± m |
Пар |
Плотность M ± m |
Относительная численность, M ± m |
||
|
2007 |
2008 |
||||||
|
2007 |
2008 |
VII |
VIII |
VI-VII |
|||
|
Красношейная поганка |
1 |
0.3±0.3 |
0.17±0.12 |
||||
|
Чирок-свистунок |
1 |
0.3±0.3 |
2 |
0.6±0.4 |
0.06±0.06 |
0.17±0.12 |
|
|
Свиязь |
2 |
0.6±0.4 |
15 |
4.3±1.1 |
0.17±0.10 |
0.17±0.12 |
1.97±0.41 |
|
Шилохвость |
1 |
0.3±0.3 |
3 |
0.9±0.5 |
0.11±0.08 |
0.34±0.17 |
|
|
Широконоска |
1 |
0.3±0.3 |
0.17±0.12 |
||||
|
Хохлатая чернеть |
15 |
4.3±1.1 |
20 |
5.7±1.3 |
0.97±0.23 |
1.30±0.34 |
3.25±0.53 |
|
Синьга Утки, ближе не опред. |
0.11±0.08 |
0.43±0.19 |
0.09±0.09 |
||||
|
Дербник |
0.06±0.06 |
||||||
|
Галстучник |
2 |
0.6±0.4 |
0.68±0.20 |
0.09±0.09 |
|||
|
Малый зуек |
3 |
0.9±0.5 |
1 |
0.3±0.3 |
0.57±0.18 |
0.35±0.17 |
0.17±0.12 |
|
Фифи |
2 |
0.6±0.4 |
3 |
0.9±0.5 |
0.11±0.08 |
0.35±0.17 |
0.43±0.19 |
|
Большой улит |
1 |
0.3±0.3 |
0.08 |
||||
|
Мородунка |
9 |
2.6±0.9 |
7 |
2.0±0.8 |
1.25±0.27 |
0.17±0.12 |
2.14±0.43 |
|
Турухтан |
0.09±0.09 |
||||||
|
Белохвостый песочник |
3 |
0.9±0.5 |
1 |
0.3±0.3 |
0.45±0.16 |
0.26±0.15 |
0.17±0.12 |
|
Бекас |
2 |
0.6±0.4 |
5 |
1.4±0.6 |
0.11±0.08 |
0.17±0.12 |
0.85±0.27 |
|
Средний кроншнеп Малый веретенник |
0.06±0.06 |
0.09±0.09 |
|||||
|
Малая чайка |
100 |
28.6±2.9 |
147 |
42.0±3.5 |
11.53±0.81 |
24.44±1.45 |
|
|
Озерная чайка |
55 |
15.7±2.1 |
31 |
8.9±1.6 |
10.97±0.79 |
3.57±0.56 |
3.68±0.56 |
|
Восточная клуша |
0.11±0.08 |
0.09±0.09 |
|||||
|
Сизая чайка |
1 |
0.3±0.3 |
0.91±0.23 |
1.30±0.34 |
0.43±0.19 |
||
|
Чайки, ближе не опред. |
0.11±0.08 |
0.17±0.12 |
0.17±0.12 |
||||
|
Речная крачка |
65 |
18.6±2.3 |
22 |
6.3±1.3 |
8.86±0.71 |
26.09±1.51 |
4.53±0.62 |
|
Береговая ласточка |
39 |
11.1±1.8 |
61 |
17.4±2.2 |
0.85±0.22 |
0.09±0.09 |
1.54±0.36 |
|
Луговой конек |
2.00±0.42 |
||||||
|
Краснозобый конек |
2 |
0.6±0.4 |
5.83±0.71 |
0.34±0.17 |
|||
|
Коньки, ближе не опред. |
4.35±0.61 |
||||||
|
Желтая трясогузка |
1 |
0.3±0.3 |
0.40±0.15 |
4.52±0.63 |
0.17±0.12 |
||
|
Белая трясогузка |
28 |
8.0±1.5 |
16 |
4.6±1.1 |
4.32±0.50 |
4.96±0.66 |
3.25±0.53 |
|
Сорока |
0.68±0.20 |
0.78±0.26 |
0.85±0.27 |
||||
|
Серая ворона |
12.16±0.83 |
2.61±0.48 |
5.13±0.66 |
||||
|
Ворон |
0.17±0.12 |
||||||
|
Свиристель Камышевка-барсучок |
10 |
2.9±0.9 |
33 |
9.4±1.6 |
1.59±0.30 |
0.26±0.15 |
7.69±0.81 |
|
Пеночка-весничка |
5 |
1.4±0.6 |
13 |
3.7±1.0 |
0.68±0.20 |
2.43±0.46 |
2.91±0.50 |
|
Пеночка-таловка |
3 |
0.9±0.5 |
12 |
3.4±1.0 |
0.23±0.11 |
0.26±0.15 |
2.74±0.48 |
|
Пеночка-теньковка Обыкновенная каменка |
1 |
0.3±0.3 |
9 |
2.6±0.9 |
0.17±0.12 |
1.71±0.38 |
|
|
Варакушка |
2 |
0.6±0.4 |
2 |
0.6±0.4 |
1.13±0.31 |
0.34±0.17 |
|
|
Рябинник |
4 |
1.1±0.6 |
7 |
2.0±0.8 |
0.45±0.16 |
0.43±0.19 |
2.14±0.43 |
|
Белобровик |
1 |
0.3±0.3 |
0.17±0.12 |
||||
|
Дрозды, ближе не опред. Домовый воробей |
68 |
19.4±2.4 |
15 |
4.3±1.1 |
0.17±0.10 |
7.04±0.78 |
0.09±0.09 |
|
Вьюрок |
3 |
0.9±0.5 |
17 |
4.9±1.2 |
0.34±0.14 |
0.52±0.21 |
6.32±0.74 |
|
Обыкновенная чечётка |
6 |
1.7±0.7 |
1.02±0.24 |
0.26±0.15 |
4.27±0.60 |
||
Продолжение таблицы 3
|
Вид |
Пар |
Плотность M ± m |
Пар |
Плотность M ± m |
Относительная численность, M ± m |
||
|
2007 |
2008 |
||||||
|
2007 |
2008 |
VII |
VIII |
VI-VII |
|||
|
Тростниковая овсянка 5 1.4±0.6 13 3.7±1.0 0.74±0.20 0.52±0.21 2.22±0.44 Овсянка-крошка 3 0.9±0.5 7 2.0±0.8 0.23±0.11 3.13±0.52 3.08±0.51 Гусеобразные 19 5.4±1.3 41 11.7±1.8 1.42±0.28 1.91±0.41 4.52±0.54 Кулики 22 6.3±1.3 17 4.9±1.2 3.24±0.43 1.57±0.37 2.84±0.43 Чайковые 221 63.1±4.3 200 57.1±4.0 32.50±1.36 31.22±1.65 25.10±1.27 Воробьиные* 103 29.4±2.9 200 57.1±4.0 23.86±1.16 41.48±1.90 33.94±1.48 Прочие 1 0.3±0.3 0.06±0.06 0.17±0.12 Всего* 366 104.6±5.5 458 130.9±6.1 61.08±1.86 75.35±2.58 66.39±2.07 |
|||||||
Примечание: нулевые значения не показаны. Для рано гнездящихся видов (сорока, серая ворона, в 2007 – чечётка) и видов-посетителей плотность гнездования не определяли. Плотность домового воробья выражена в особях на 1 км2.
* - число пар и плотность птиц без домового воробья.
К числу обычных видов с плотностью гнездования от 2 до 9.9 пар на 1 км2 в 2007 году относились 5 видов: хохлатая чернеть, мородунка, белая трясогузка, камышевка-барсучок, домовый воробей, в 2008 – 15: свиязь, хохлатая чернеть, мородунка, озёрная чайка, речная крачка, белая трясогузка, камышевка-барсучок, весничка, таловка, теньковка, рябинник, домовый воробей, юрок, тростниковая овсянка и овсянка-крошка. Остальные виды с плотностью гнездования менее 2 пар/км2 были немногочисленными или редкими – за все время учётов отмечены единичные пары, выводки или небольшое число особей. Некоторые малочисленные виды гнездились в пределах учётных площадок не ежегодно.
Наиболее многочисленными птицами были чайковые – малая и озёрная чайки, речная крачка. Вся эта группа составляла порядка 5060% от общей численности птиц. Второе место занимали воробьиные, численность уток и куликов была в 5-10 раз ниже.
Обилие птиц, в том числе обычных и многочисленных видов, сильно менялось по годам. Особенно резкие изменения плотности отмечены у свиязи (7-кратные), речной крачки, камышевки-барсучка, пеночек, юрка. Различия в плотности гнездования отдельных групп видов и птиц в целом в оба сезона были не столь значительными.
Внутрисезонная динамика состава и численности птиц
В августе 2007 года было зарегистрировано несколько «новых» видов, отсутствовавших в июле: синьга, турухтан, луговой и краснозобый коньки, жёлтая трясогузка, ворон, свиристель (табл. 2). Жёлтых трясогузок стали регистрировать с 26 июля, в августе их численность вы- росла в 10 раз. Коньков встречали с 4 августа. В это же время увеличилась численность фифи, речной крачки, веснички, домового воробья, юрка, овсянки-крошки. Появление и рост численности большинства названных видов связано с началом масштабных послегнездовых кочёвок и миграции, во время которых большинство птиц, закончивших размножение, скатывается с водоразделов в пойму. Речные крачки после вылета молодых продолжали держаться в месте рождения. Из-за особенностей поведения учёт варакушек после начала насиживания становится затруднительным, поэтому в начале июля 2007 две пары не были учтены – их удалось обнаружить только в августе, когда они стали активно беспокоиться. В это же время мы стали встречать и кочующих варакушек.
Некоторые виды отсутствовали в августовских учётах, у других численность заметно снизилась. В частности, раннее исчезновение малой чайки может быть вызвано гибелью колонии из-за подтопления гнёзд при резком подъёме уровня воды в период гнездования. Этот вид устраивает гнёзда на небольших кочках или у их основания очень невысоко над водой, поэтому гнёзда весьма чувствительны к подъёму уровня воды. Расчётные сроки размножения показывают, что вылупление птенцов у малой чайки в 2007 году происходило около 20 июня, к 15-20 июля молодые должны были подняться на крыло (птенцы становятся лётными на 23-25-й день). Некоторое время они держатся вблизи места гнездования и, следовательно, хотя бы часть птиц должна была попасть в поле зрения. Однако малые чайки полностью отсутствовали в учётах. В то же время озёрных чаек мы наблюдали, хотя их численность снизилась почти втрое по сравнению с июлем. Это объясняется перемещением части птиц на магистраль Оби к местам скопления молоди рыб. В отличие от малых, у озёрных чаек гнездо представляет собой громоздкое конусообразные сооружения из травы, способное даже плавать. Поэтому повышение уровня воды на них сказалось в меньшей степени.
Уменьшение численности ряда куликов: галстучника, малого зуйка, мородунки, белохвостого песочника,– а также ласточки-береговушки мы связываем с началом их отлёта. Камышевка-барсучок не попала в учёты из-за того, что этот скрытный вид плохо регистрируется при отсутствии поющих самцов. Более высокая численность серой вороны в июле связана со случайной встречей одного крупного скопления птиц на свалке на площадке № 1.
Таким образом, внутрисезонные изменения состава и численности птиц в обследованной антропогенной пойме вполне соответствуют хорошо известной естественной динамике их населения в пойме Нижней Оби, типичной для второй половины лета. Высокая обводнённость местообитаний не оказала заметного влияния на её течение, за ис- ключением одного вида – малой чайки, которая раньше обычного покинула район гнездования из-за неудачного размножения.
Состав и численность птиц при разном уровне обводнённости
При низком уровне и сокращенной продолжительности затопления пойменных местообитаний в 2008 году ряд видов увеличил численность по сравнению с 2007 годом (табл. 3). Достоверно выросла численность свиязи, малой чайки, береговушки, камышевки-барсучка, юрка, воробьиных и всех птиц в целом ( P < 0.01), а также пеночек: та-ловки ( P < 0.05) и веснички ( P < 0.10). На гнездовании были отмечены широконоска, краснозобый конёк, жёлтая трясогузка, теньковка и белобровик, которых в 2007 году не обнаружили. В то же время снизилась численность почти у всех куликов (за исключением массовых – фифи и бекаса) и некоторых других видов. Значимые изменения зафиксированы у озёрной чайки, речной крачки и домового воробья ( P < 0.01), а также белой трясогузки ( P < 0.90). Для остальных видов, в основном по причине их малочисленности, изменения выглядят как случайные.
Увеличение плотности гнездования гусеобразных, малой чайки и многих воробьиных связано, очевидно, с освобождением от затопления их типичных гнездовых и кормовых биотопов. С другой стороны, снижение уровня водности привело к сокращению площади удобных мест гнездования озёрной чайки, избегающей строить гнезда на слишком маловодных или полностью осушенных местах. Уменьшение численности других видов (крачки, белой трясогузки и домового воробья), судя по всему, с изменением режима обводненности не связаны.
Следует также отметить, что относительные показатели численности менее точно характеризуют её изменения – повторный учёт одних и тех же птиц на сравнительно коротких маршрутах в пределах ограниченных территорий приводит к завышенной их оценке.
Изменение пространственного распределения птиц в зависимости от уровня и продолжительности затопления поймы
В 2008 году, по сравнению с 2007, заметные изменения произошли в пространственном распределении птиц, что мы также связываем с разным характером затопления поймы в эти годы.
Число колоний и групповых поселений чаек и речной крачки на учётных площадках в 2008 году, по сравнению с 2007 годом, существенно не изменилось (рис. 1, табл. 4): в 2007 на площадке № 1 обнаружена 1 колония, в 2008 – 2, на площадке № 2, соответственно, по 5. Но в 2008 году за пределами учётных площадок обнаружены ещё 3
колонии, тогда как в 2007 году – только одна. Колонии 1а, 1б и 4 в 2008 году возникли на полуобсохших луговинах, которые в 2007-м полностью находились под водой. Колония 3 сместилась: в 2007 она была на сравнительно высоком мысу озера, в 2008 – на пойменном лугу у берега этого же озера.
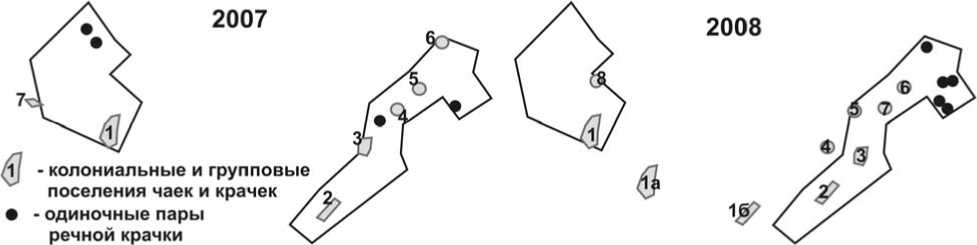
Рис. 1. Размещение мест гнездования чаек и речной крачки в районе работ в 2007 и 2008 годах (сплошная линия – границы участков; площадка № 1 на рисунках слева;
1-8 – номера колоний и групповых поселений).
Таблица 4. Состав колоний и групповых поселений чайковых в 2007 и 2008 годах
|
Вид |
№ колонии |
|||||||||||||
|
2007 год |
2008 год |
|||||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
1 |
2 |
3 |
5 |
6 |
7 |
8 |
|
|
Малая чайка 100 — — — — — — 70 15 40 20 2 — — Озёрная чайка 15 5 — — — — 35 20 1 1 5 — — 5 Сизая чайка — 1 — — — — — — — — — — — — Речная крачка 40 3 14 3 3 2 — 15 2 2 — 2 3 — |
||||||||||||||
Образование малой и озёрной чайками крупных колоний, насчитывающих более сотни гнезд,– достаточно обычное явление в регионе (Пасхальный 2001; Головатин и др. 2007). Но в 2008 году эти чайки более равномерно распределялись по территории, если оценивать величину отдельных поселений (табл. 4). Это свидетельствует о том, что у птиц появились новые возможности загнездиться на менее затопленных участках поймы.
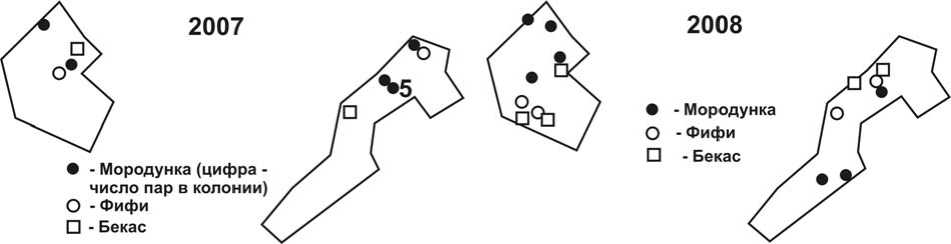
Рис. 2. Размещение гнездовых территорий некоторых видов куликов на учётных площадках в 2007 и 2008 годах. Обозначения – как на рис. 1.
Среди куликов наиболее заметные различия в локализации территорий в 2008 по сравнению с 2007 наблюдались у фифи и бекаса (рис. 2). Они заключались в том, что птицы занимали более низкие уровни поймы. Расширилась область гнездования и у мородунки.
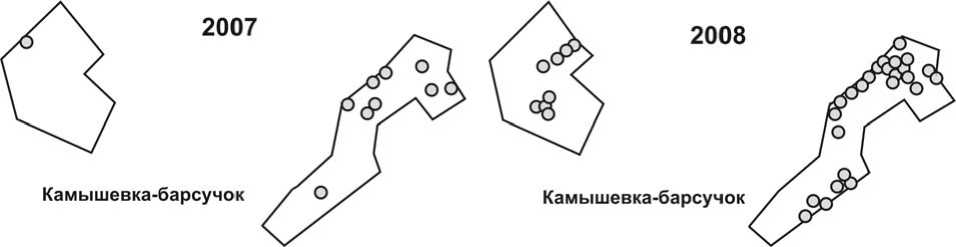
Рис. 3. Размещение гнездовых территорий камышевки-барсучка на учётных площадках в 2007 и 2008 годах. Обозначения – как на рис. 1.
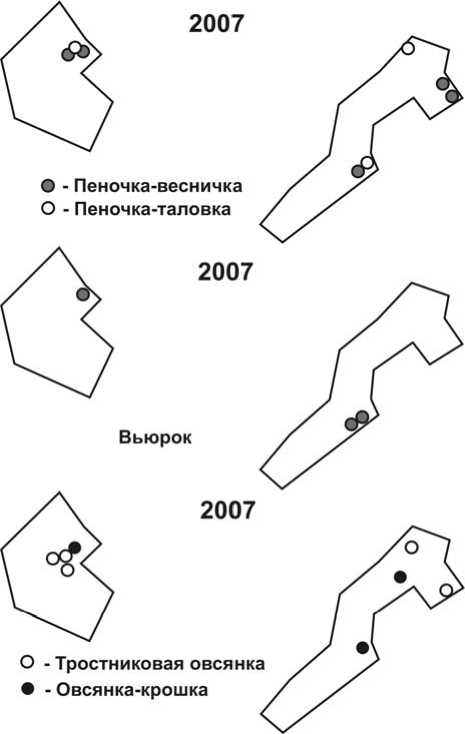
Рис. 4. Размещение гнездовых территорий пеночек, вьюрка и овсянок на учётных площадках в 2007 и 2008 годах. Обозначения – как на рис. 1.
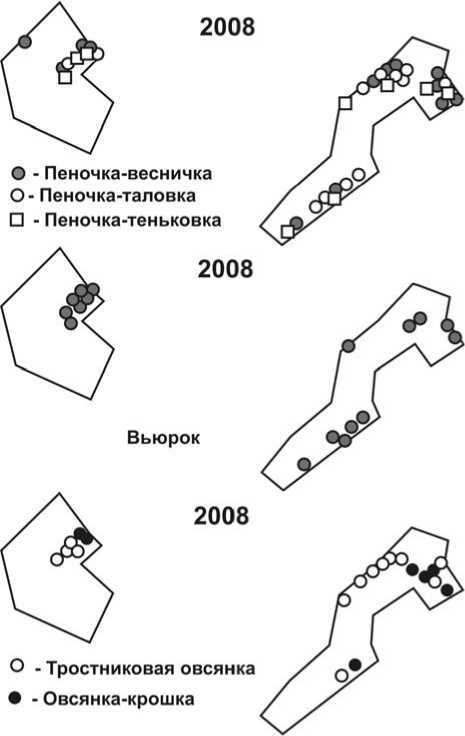
В 2008 году значительно увеличилась площадь, на которой гнездились камышевки-барсучки (рис. 3), особенно за счёт занятия этими птицами участков низкорослых ивняков и луговин, расположенных на низких высотных уровнях поймы, которые в 2007 году были затоплены.
Сформировались очень плотные поселения в оптимальных для вида местообитаниях.
Сходная картина распределения гнездовых территорий отмечена у пеночек – веснички и таловки (рис. 4). Кроме того, в 2008 году на гнездовании зарегистрированы 9 пар пеночек-теньковок, которых в предшествующем году вообще не отмечали.
Аналогичным образом на умеренное затопление территории отреагировали юрок и тростниковая овсянка, в меньшей степени – овсянка-крошка (рис. 4). При этом юрки, придерживаясь в основном высокорослых ивняков на гривах, встречались даже в зарослях заболоченного кустарника высотой до 2 м. В последнем местообитании появилось и значительное число гнездовых пар тростниковой овсянки, что особенно наглядно было видно на площадке № 2.
Заключение
Влияние режима обводнённости отражается на пространственном распределении птиц поймы. При высоком уровне воды, как в 2007 году, площадь удобных для гнездования мест для видов, гнездящихся или собирающих корм на земле, сокращается. Это вызывает снижение численности птиц на данной территории. При более низком, соответствующем среднему уровне водности (2008 год) расширение площади, пригодной для гнездования, ведёт к более широкому распределению птиц по пойме, что, в свою очередь, приводит к увеличению их численности. Это характерно для речных уток, малой чайки, массовых видов куликов (фифи и бекаса) и воробьиных.
Высокий уровень воды практически не отражается на хохлатой чернети, озёрной чайке и речной крачке. Хохлатая чернеть при высокой воде может устраивать гнёзда на затопленных луговинах, размещая их на вершинах выступающих над водой кочек, сплавинах, плавающих брёвнах и т.п. местах. Озёрная чайка строит достаточно массивные гнёзда из растительных остатков и прошлогодней травы, которые она достраивает при увеличении уровня воды и которые, в крайнем случае, могут плавать. Более того, высокий уровень обводнённости улучшает защитные свойства гнёзд, т.к. снижается их доступность для хищников. При более низкой воде защитные свойства территории уменьшаются и птицы перестают гнездиться на обсыхающих участках. Этим можно объяснить падение численности озёрной чайки в 2008 году. Речные крачки гнездятся в основном на насыпных островках, по окраинам дамб, т.е. на сравнительно высоких участках, редко заливаемых водой.
Внутрисезонная динамика населения птиц при высокой водности соответствует обычному для данного района движению населения птиц во второй половине лета.
В целом можно заключить, что режим обводнённости в пойме Нижней Оби оказывает существенное и дифференцированное влияние на состав и численность птиц, гнездящихся здесь. Многоводные годы, судя по имеющимся наблюдениям, для большинства видов оказываются менее благоприятными из-за сокращения мест гнездования и кормёжки.
