Население стратобионтных пауков (Araneae) лесов национального парка "Припышминские боры" (летний аспект)
 лесов национального парка \"Припышминские боры\" (летний аспект)")
Автор: Есюнин С.Л., Тураева А.С.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 2, 2016 года.
Бесплатный доступ
Изучены группировки пауков, обитающих в подстилке, в разнотипных и разновозрастных (15, 25 и 70-летние сосновые посадки, естественные березово-сосновые, сосново-еловые и еловые) лесах национального парка «Притпнминские боры» (Свердловская обл.). Материал получен в летние месяцы (июнь, июль) 1997 и 2002 гг. по единой методике. Пробы подстилки размером 25 на 25 см помещали в энтомологическое сито; пауков из мелкоразмерного «остатка» выбирали вручную. Группировки пауков сосновых посадок и естественных лесов отличаются друг от друга. В сосновых лесопосадках обнаружено больше видов (18-25, против 6-10 в лесах), соотношение таксонов здесь более выравненное (H' = 2.32.7, против H' = 1.3-2.1 в лесах), присутствует ряд видов (например, Abacoproeces saltuum (L. Koch, 1872), Pocadicnemis pumila (Blackwall, 1841), Tapinocyba insecta (L. Koch, 1869)), не отмеченных в лесах. Пробы пауков из естественных лесов отличаются присутствием обильного вида в доминантном комплексе (Diplocentria bidentata (Emerton, 1882)) и высокой долей пауков-линифиид (86-100% от всех особей). В посадках сосны плотность пауков варьирует от 35.1 до 95.6 экз./м2; в молодых лесопосадках (10-15 лет) многочисленны представители семейств Linyphiidae, Gnaphosidae, Theridiidae, Lycosidae и наиболее обильны неполовозрелые Haplodrassus sp. и Tapinocyboides pygmaeus (Menge, 1869); в прочих лесопосадках многочисленны Linyphiidae (2/3 всех особей) и Theridiidae, доминируют D. bidentata и Robertus lividus (Blackwall, 1836). В лесах плотность пауков увеличивается с возрастом от 45-53 экз./м2 в смешанных березово-сосновых лесах, до 168 экз./м2 в еловом лесу; в смешанных лесах доминирует D. bidentata; в еловом лесу - D. bidentata, Erigonella hiemalis (Blackwall, 1841) и Minyriolus pusillus (Wider, 1834).
Пауки, население, летний аспект, хвойные леса, южная тайга, западно-сибирская равнина
Короткий адрес: https://sciup.org/147204767
IDR: 147204767 | УДК: 595.44(571.1)
The stratobios spider populations (Araneae) of forests of the Pripyshminskie bory national park (summer aspect)
Assemblages of spiders, inhabiting a litter of different-type and uneven-age coniferous forests (15, 25 and 70-years pine tree-planting, natural birch-pine, pine-spruce and spruce forests), are studied in "Pripyshminskie Bory" National Park (Sverdlovsk Area). The material was sampled during the summer months (June, July) 1997 and 2002 using identical technique and methods. The samples of litter with size 25 x 25 cm. were passed through the entomology sieve for exclusion of a large size fractions. Spiders from the 'small size remnant' were captured manually. The spiders assemblages of the investigated pine tree-plantings and natural forests are differs. The samples collected in the pine tree-plantings contain more species (18-25) than those gathered in the forest (6-10), with more leveled ration parity of taxa (Shannon index (H') = 2.3-2.7, against H' = 1.3-2.1 in forests). Some species, which were not detected in the forests (for example, Abacoproeces saltuum (L. Koch, 1872), Pocadicnemis pumila (Blackwall, 1841), Tapinocyba insecta (L. Koch, 1869)), are present in pine tree-plantings. Controversially, the samples of spiders from natural forests are characterized by presence of a single abundant species (Diplocentria bidentata (Emerton, 1882)) together with a high ratio of linyphiid-spiders specimens (86-100%). The density of spiders varies from 35.1 up to 95.6 specimens/m2 in pine tree-plantings. Specimens belonging to four families (Linyphiidae, Gnaphosidae, Theridiidae, Lycosidae) are most numerous in the young pine tree-planting (10-15 years), with immature specimens of Haplodrassus sp. and Tapinocyboides pygmaeus (Menge, 1869) showing maximal ability. Only two families, Linyphiidae (2/3 of all individuals collected) and Theridiidae, are numerous in 20-25 and 70 years aged tree-plantings, with dominance of D. bidentata and Robertus lividus (Blackwall, 1836). The density of spiders increases with the age of the forests from 45-53 specimens/m2 in the mixed birch-pine forests, up to 168 specimens/m2 in spruce forest (climax commu nity). Whereas only one species D. bidentata has been noticed as a dominant in the mixed forests, the three co-dominating species, D. bidentata, Erigonella hiemalis (Blackwall, 1841) and Minyriolus pusillus (Wider, 1834), were detected in the spruce forest.
Текст научной статьи Население стратобионтных пауков (Araneae) лесов национального парка "Припышминские боры" (летний аспект)
Фауна пауков Западной Сибири на сегодняшней момент изучена достаточно полно [Mikhailov, 2013]. Для подзоны южной тайги Западно-Сибирской равнины, где располагается национальный парк «Припышминские боры», отмечено 350 видов пауков [Есюнин, Стёпина, 2014].
Гораздо меньше информации о населении пауков Западной Сибири. Наиболее хорошо изучена структура группировок герпетобионтных пауков лесотундры Южного Ямала [Коробейников, Есюнин, 1995; Есюнин, 1999] и лесостепной зоны [Есюнин, Ширпужева, 2006; Пахоруков и др., 1983; Пахоруков, 1985; Esyunin, 2008]. Первые, и на данный момент единственные, данные о плотности и видовом составе пауков различных лесных и производных от них биоценозов тайги ЗападноСибирской равнины приведены в работе Б.Р. Стригановой и Н.М. Порядиной [2005]. Кроме того, А.Е. Волков [1987], основываясь на ручных сборах, описывает состав «фоновых» видов пауков некоторых биоценозов южной тайги Тюменской области.
Ранее для национального парка «Припышминские боры», в фауне которого насчитывается 141 вид пауков [Есюнин, Стёпина, 2014; Тураева, Есюнин, 2015], был описан осенний аспект населения герпето- и хортобионтных пауков [Тураева, Есюнин, 2015].
Цель данной работы - описание летнего аспекта населения пауков, населяющих подстилку лесных биоценозов национального парка «Припышминские боры».
Материал и методы
Национальный парк «Припышминские боры» располагается на юго-востоке Свердловской обл. (Тугулымский и Талицкий р-ны). Леса занимают 88% территории национального парка. Преобладают сосновые леса (43% покрытой лесом площади). На долю ельников, произрастающих здесь вблизи южной границы своего ареала, и сохранившихся преимущественно в наиболее пониженных и увлажненных местообитаниях, приходится всего 2-4% площади парка, покрытой лесом [Национальный парк ..., 2016].
Население пауков подстилки было изучено в семи вариантах лесонасаждений:
1) посадки сосны, возраст 10-15 лет, проективное покрытие травостоя (ПП) - 5%, преобладающие растения - сныть, злаки, чина;
-
2) посадки сосны с березой и осиной, возраст 20-25 лет, ПП - 10%, преобладающие растения -злаки, костяника, земляника;
-
3) посадки сосны, возраст около 70 лет, 1111 -80%, преобладающие растения - папоротник, злаки, земляника;
-
4) лес березово-сосновый с травостоем из папоротника, злаков, черники, крапивы;
-
5) лес березово-сосновый с травостоем из злаков, сныти, плауна, крапивы;
-
6) лес сосново-еловый с травостоем из кислицы, земляники, черники, крапивы;
-
7) лес еловый, в травостое сныть, кислица, плаун, крапива.
Исследования осуществлены по единой методике, но разными авторами и в разные годы. Биотопы 1-3 обследованы 20-23 июня 2002 г. Л. Шу-миловских, а биотопы 4-7 - 1-23 июля 1997 г. Д. Казанцевым. Пробы подстилки размером 25 на 25 см (1/16 м2) помещали в энтомологическое сито (сифтер) [Голуб и др., 2012] для отделения крупного опада, ветвей и т.п. Пауков из отсифтованного материала выбирали вручную в лабораторных условиях.
Сходство группировок пауков оценивалось при помощи индекса Чекановского-Соренсена (/cs); для оценки разнообразия населения использован индекс Шеннона ф*У Расчеты и кластерные построения выполнены с применением программы BIODIV [Baev, Penev, 1993]. Оценка обилия видов пауков проводилась с использованием пятибалльной ограниченной сверху логарифмической шкалы относительного обилия видов, предложенной Ю.А. Песенко [1982].
Население пауков
Так как пробы брались в различное время, анализ данных проводится раздельно для лесопосадок и естественных лесонасаждений.
Население пауков лесопосадок сосны
Плотность пауков в посадках сосны варьирует от 35.1 до 95.6 экз./м2 (табл. 1). В молодых посадках (10-15 лет) население пауков характеризуется относительно высоким разнообразием семейств. Здесь многочисленны пауки-линифииды (табл. 1) и гнафозиды (32% от общей плотности) и обычны представители семейств Theridiidae (9%) и Lycosi-dae (5%). В более возрастных посадках таксономическая структура упрощается. Подавляющая часть населения пауков (не менее 90%) представлена особями из двух семейств: пауки-линифииды со-
ставляют более 2/3 населения, а пауки-теридииды - 17 и 12% во 2- и 3-м биотопах, соответственно.
Linyphiidae (табл. 1). Из видов, представленных в этом биотопе половозрелыми особями, обычен Tapinocyboides pygmaeus.
Количество видов пауков, обнаруженных в пробах, варьирует от 18 до 27. Индекс разнообразия Шеннона имеет значения от 2.3 до 2.7, что свидетельствует о высоком разнообразии изученных группировок пауков.
В возрастных посадках таксономическую структуру отражает состав доминантных видов (табл. 2). Здесь многочисленны два вида: Diplocen-tria bidentata (Linyphiidae) и Robertus lividus (Ther-idiidae). В молодых посадках летом преобладают неполовозрелые особи из рода Haplodrassus и сем.
Таблица 1
Видовой состав, плотность (экз./м2) и показатели разнообразия населения стратобинтных пауков лесных биотопов национального парка «Припышминские боры», летний аспект
|
Таксон |
Номер биотопа* |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
|
Araneidae |
|||||||
|
Araneus sp. (неполовозрелые) |
0.6 |
0.5 |
|||||
|
Clubionidae |
|||||||
|
Clubiona sp. (неполовозрелые) |
1.1 |
||||||
|
Dictynidae |
|||||||
|
Emblyna burjatica (Danilov, 1994) |
4.0 |
||||||
|
Gnaphosidae |
|||||||
|
Haplodrassus silvestris (Blackwall, 1833) |
0.6 |
||||||
|
H. soerenseni (Strand, 1900) |
0.6 |
3.2 |
|||||
|
H. umbratilis (L. Koch, 1866) |
1.7 |
||||||
|
Haplodrassus sp. (неполовозрелые) |
12.6 |
2.3 |
|||||
|
Zelotes clivicola (L. Koch, 1870) |
0.6 |
||||||
|
Zelotes sp. (неполовозрелые) |
2.9 |
||||||
|
Liocranidae |
|||||||
|
Agroeca sp. (неполовозрелые) |
0.6 |
||||||
|
Linyphiidae |
|||||||
|
Abacoproeces saltuum (L. Koch, 1872) |
2.9 |
0.5 |
1.1 |
||||
|
Agyneta olivacea (Emerton, 1882) |
0.6 |
2.1 |
0.6 |
||||
|
A. ramosa Jackson, 1912 |
1.6 |
5.1 |
|||||
|
Anguliphantes angulipalpis (Westring, 1851) |
1.7 |
3.2 |
|||||
|
Bathyphantes nigrinus (Westring, 1851) |
4.0 |
||||||
|
Centromerus arcanus (O.P.-Cambridge, 1873) |
20.0 |
||||||
|
Diplocentria bidentata (Emerton, 1882) |
3.2 |
13.7 |
26.7 |
9.6 |
36.0 |
72.0 |
|
|
Diplocephalus picinus (Blackwall, 1841) |
5.3 |
||||||
|
Erigonella hiemalis (Blackwall, 1841) |
0.5 |
5.3 |
3.2 |
28.0 |
|||
|
Helophora insignis (Blackwall, 1841) |
5.3 |
||||||
|
Hypselistes jacksoni (O.P.-Cambridge, 1902) |
0.6 |
2.3 |
2.3 |
4.0 |
|||
|
Macrargus rufus (Wider, 1834) |
1.1 |
0.6 |
6.4 |
||||
|
Maro pansibiricus Tanasevitch, 2006 |
1.7 |
2.1 |
0.6 |
||||
|
Maro Tsublestus Falconer, 1915 |
4.0 |
12.0 |
|||||
|
Maso sundevalli (Westring, 1851) |
0.6 |
||||||
|
Micrargus herbigradus (Blackwall, 1854) |
0.6 |
1.7 |
5.3 |
8.0 |
|||
|
Microneta viaria (Blackwall, 1841) |
0.5 |
6.4 |
|||||
|
Minyriolus pusillus (Wider, 1834) |
1.1 |
20.0 |
|||||
|
Neriene sp. (неполовозрелые) |
0.6 |
||||||
|
Pocadicnemis pumila (Blackwall, 1841) |
0.6 |
1.1 |
|||||
|
Tapinocyba insecta (L. Koch, 1869) |
0.5 |
0.6 |
|||||
|
Tapinocyboides pygmaeus (Menge, 1869) |
6.3 |
4.0 |
|||||
|
Tapinopa longidens (Wider, 1834) |
4.0 |
||||||
|
Tibioplus diversus (L. Koch, 1879) |
0.6 |
4.0 |
|||||
|
Walckenaeria cucullata (C.L. Koch, 1836) |
0.6 |
||||||
|
W. atrotibialis (O. Pickard-Cambridge, 1878) |
5.3 |
4.0 |
|||||
|
W. karpinskii (O. Pickard-Cambridge, 1873) |
0.6 |
4.0 |
|||||
|
W. nudipalpis (Westring, 1851) |
0.6 |
||||||
|
W. picetorum (Palmgren, 1976) |
4.0 |
||||||
|
Linyphiidae sp. (неполовозрелые) |
10.9 |
12.3 |
38.3 |
8.0 |
|||
Окончание табл. 1
|
Таксон |
Номер биотопа* |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
|
Lycosidae |
|||||||
|
Pardosa lugubris (Walckenaer, 1802) |
0.6 |
||||||
|
P. palustris (Linnaeus, 1758) |
0.6 |
||||||
|
Pardosa sp. (неполовозрелые) |
0.6 |
||||||
|
Trochosa terricola Thorell, 1856 |
0.6 |
||||||
|
Trochosa sp. (неполовозрелые) |
0.6 |
0.5 |
|||||
|
Miturgidae |
|||||||
|
Zora nemoralis (Blackwall, 1861) |
0.6 |
||||||
|
Z. spinimana (Sundevall, 1833) |
1.1 |
||||||
|
Philodromidae |
|||||||
|
Thanatus sabulosus (Menge, 1875) |
0.6 |
||||||
|
Salticidae |
|||||||
|
Neon reticulatus (Blackwall, 1853) |
0.5 |
1.7 |
8.0 |
||||
|
Pseudeuophrys erratica (Walckenaer, 1826) |
0.6 |
||||||
|
Tetragnathidae |
|||||||
|
Metellina sp. (неполовозрелые) |
1.1 |
0.5 |
|||||
|
Tetragnatha pinicola L. Koch, 1870 |
1.1 |
||||||
|
Theridiidae |
|||||||
|
Crustulina guttata (Wider, 1834) |
0.5 |
||||||
|
Enoplognatha ovata (Clerck, 1758) |
0.6 |
3.2 |
|||||
|
Parasteatoda sp. (неполовозрелые) |
0.6 |
||||||
|
Robertus lividus (Blackwall, 1836) |
4.0 |
4.8 |
10.5 |
9.6 |
16.0 |
||
|
Theridion sp. (неполовозрелые) |
1.1 |
0.5 |
|||||
|
Thomisidae |
|||||||
|
Ozyptilapraticola (C.L. Koch, 1837) |
0.6 |
0.6 |
4.0 |
||||
|
Xysticus sp. (неполовозрелые) |
2.3 |
1.1 |
|||||
|
Общая плотность |
58.8 |
35.1 |
95.6 |
53.2 |
44.8 |
100.0 |
168.0 |
|
Доля (% особей) пауков сем. Linyphiidae |
44 |
77 |
77 |
100 |
64 |
92 |
86 |
|
Количество видов |
25 |
18 |
27 |
6 |
8 |
9 |
10 |
|
Индекс Шеннона |
2.72 |
2.35 |
2.32 |
1.30 |
2.09 |
2.08 |
1.79 |
Примечание. * Номера изученных биотопов приведены в тексте.
Население пауков лесов
Плотность пауков в изученных лесах значительно различается (почти в 4 раза, табл. 1), увеличиваясь с 45-53 экз./м2 в смешанных березовососновых лесах, до 168 экз./м2 - в еловом лесу. Таксономическая структура группировок пауков лесов проста. Здесь резко преобладают пауки из сем. Linyphiidae (табл. 1), составляющие до 100% всех отловленных особей. Реже (биотопы 5 и 7), кроме пауков-линифиид относительно многочисленны представители сем. Theridiidae. Обращает на себя внимание почти полное отсутствие в этих биоценозах незрелых особей пауков-линифиид.
В смешанных лесах имеется один обильный вид - D. bidentata (табл. 2). В еловом лесу комплекс доминантных видов разнообразнее; кроме упомянутого вида, здесь обычны Erigonella hie-malis и Minyriolus pusillus.
Таблица 2
Состав доминантного комплекса населения стратобионтных пауков лесных биотопов национального парка «Припышминские боры», летний аспект
|
Вид |
Номер биотопа* |
||||||
|
Посадки сосны |
Хвойные леса |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
|
Tapinocyboides pygmaeus |
Ш |
- |
- |
- |
- |
- |
- |
|
Robertus lividus |
- |
ш |
ш |
- |
- |
- |
- |
|
Agyneta ramosa |
- |
- |
ш |
- |
- |
- |
- |
|
Diplocentria bidentata |
- |
ш |
IV |
IV |
ш |
IV |
IV |
|
Centromerus arcanus |
- |
- |
- |
- |
- |
ш |
- |
|
Erigonella hiemalis |
- |
- |
- |
- |
- |
- |
ш |
|
Minyriolus pusillus |
- |
- |
- |
- |
- |
- |
ш |
Примечание. Ш - обычный, IV - обильный вид [Песенко, 1982]. * Номера изученных биотопов как в табл. 1.
Количество видов пауков, обнаруженных в пробах, варьирует от 6 до 10, несколько возрастая в ряду от березово-сосновых лесов к еловому лесу (табл. 1). Индекс разнообразия Шеннона имеет значения от 1.3 до 2.1, что свидетельствует о более низком, в сравнении с лесопосадками, разнообразии изученных группировок пауков лесных биоценозов.
Обсуждение результатов
Пробы пауков из лесопосадок и естественных лесов отличаются друг от друга (рисунок). В сосновых лесопосадках обнаружено больше видов, соотношение таксонов здесь более выравненное, присутствует ряд обильных видов, не отмеченных в лесах. Пробы пауков из естественных лесов, в свою очередь, отличаются присутствием обильного вида в доминантном комплексе, высокой долей пауков-линифиид и отсутствием неполовозрелых особей из сем. Linyphiidae. Наряду с различиями, имеются и общие черты, а именно, наличие общего доминанта (/L hideniaia). значительная представленность в преобладающем количестве проб вместе с пауками-линифиидами пауков из сем. Theridiidae, несколько общих для всех биотопов видов (например, Erigonella hiemalis, Hypselistes jacksoni, Micrargus herbigradus, Ozyptila praticola). Обращает на себя внимание тот факт, что общие черты касаются в значительной степени качественных показателей, а различия - количественных.
По-видимому, большое количество незрелых пауков из сем. Linyphiidae в июньских пробах из лесопосадок, и их отсутствие в июльских пробах из лесов связаны, в первую очередь, с различиями в календарных сроках взятия проб, т.к. многие бореальные виды пауков-линифиид вылупляются из яиц летом и достигают половой зрелости к осени. Различия погодных условий 1997 и 2002 гг., предположительно, объясняют такие «странности», как присутствие в пробах из лесопосадок и, наоборот, отсутствие в лесных пробах таких типичных лесных видов, как Agyneta olivacea, Maro pansibiricus, Tibioplus diversus. Суммируя высказанные замечания, приходим к выводу, что обсуждаемые различия могут быть обусловлены целым комплексом факторов. По этой причине, мы остановимся на анализе специфики структуры населения пауков только молодых сосновых посадок.
По своей структуре население пауков сосновых посадок 10-15-летнего возраста имеет переходный характер. Это уже не луговые группировки, но еще и не лесные. От лесных их отличает низкая доля пауков-линифиид, значительная представленность в населении пауков из семейств Gnaphosidae (32%) и Lycosidae (5%), отсутствие типичного для лесов доминантного вида (Z). bideniaia) и высокая вы- равненность населения (Н’ = 2.7). При этом в биотопе уже обычны такие лесные виды, как АЬасо-proeces saltuum, A. olivacea, М. herbigradus, R. lividus, T. diversus и т.д.
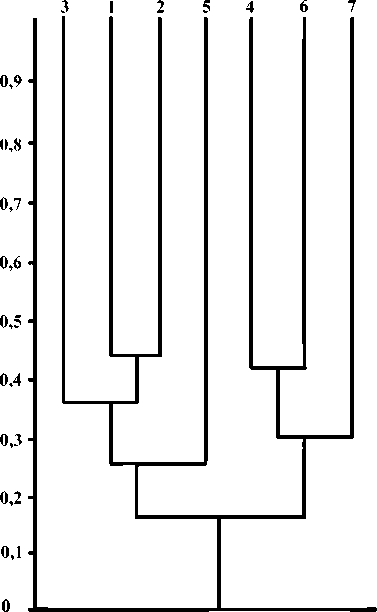
Кладограмма сходства (/cs) проб пауков из лесных биоценозов национального парка «Припышминские боры» (номера биотопов как в табл. 1)
Сравнение полученных нами характеристик населения пауков с данными других авторов затруднено двумя моментами. Во-первых, значительное количество сведений о плотности пауков хвойных лесов, полученных почвенными зоологами с использованием стандартных почвенных раскопок [Количественные ..., 1987], явно занижены. Как правило, приводимая плотность измеряется единицами экз./м2. Например, для подзоны южной тайги Западно-Сибирской равнины Б.Р. Стриганова и Н.М. Порядина [2005] приводят следующие данные. Плотность пауков в ельнике зеленомошном составляла 21.6, а в сосняке кладониевом - 24.0 экз./м2. Во-вторых, не все работы содержат информацию о фенологическом периоде, когда были взяты пробы.
Летний аспект структуры населения пауков изучала Т.А. Гаркуша [1975] в елово-широколиственных лесах Волжско-Камского заповедника. Исследуя особенности парцеллярной структуры, автор отмечает, что в различных парцеллах плотность пауков варьировала от 16 до 92 экз./м2 в
1972 г. и от 27 до 42 экз./м2 - в 1973 г., что существенно ниже полученных нами результатов для лесов с участием ели, но сопоставимо с показателями сосновых лесопосадок и смешанных березово-сосновых лесов. Сопоставимые с нашими результаты по летнему аспекту населения пауков были получены Н.Л. Уховой [1999] для пихтовоелового высокотравно-папоротникового леса в Ви-симском заповеднике (Свердловская обл., Кировоградский р-н), где плотность пауков составляла 112.9 экз./м2. Наконец, в работе М.Т. Штернбергса [1984, с. 191], посвященной описанию структуры группировок пауков ельника зеленомошного (Латвия), отмечается, что «наиболее низка она (плотность пауков - С.Е.) в июле (до 75 особей на 1 м2)».
Как правило, летние аспекты населения пауков характеризуются более низкими значениями плотности. Так, в пихтово-еловых лесах Висимского заповедника плотность пауков осенью хоть и незначительно, но возрастает по сравнению с летним аспектом [Ухова, 1999], колеблясь в различные годы от 113 до 162 экз./м2 [Ухова, 1999; Есюнин и др., 2000]. Близкие значения плотности пауков (108 экз./м2) получены для сосновых лесов Ильменского заповедника (Челябинская обл.) [Мазура, 1995]. В латвийских ельниках осенью плотность пауков увеличивается до 180 экз./м2 [Штернберге, 1984].
Таким образом, можно констатировать следующее. Плотность пауков в летних группировках в национальном парке «Припышминские боры» в целом соизмерима со значениями плотности в других регионах в этот же фенологический период. Летние значения плотности пауков в хвойных лесах ниже таковых в осенний период.
Набор доминирующих видов пауков в лесах национального парка «Припышминские боры» весьма специфичен. Только два вида из нашего списка (табл. 2): D. bidentata и R. lividus, упоминаются в качестве доминантов в группировках пауков хвойных лесов других регионов. Например, первый вид, является третьим по обилию в списке доминирующих видов в еловых лесах Латвии [Штернберге, 1984], где он менее обилен, чем Minyriolus pusillus и Tapinocyba pallens (О. Pickard-Cambridge. 1872). Но его нет в составе доминантов группировок пауков из латвийских сосняков [Штернберге, 1985] и хвойных лесов Онежского стационара (Архангельская обл.) [Кудряшева, 1987]. Второй вид из нашего списка обнаружен в составе доминантного комплекса, описанного для среднеуральских горных темнохвойных лесов (летний аспект) [Ухова и др., 2000], где, кроме него, многочисленны Bathyphantes nigrinus, Macrar-gus rufus, Micrargus herbigradus и Panamomops dy-bowskii (O.Pickard-Cambridge, 1873). Кроме того,
R. lividus, наряду c Anguliphantes angulipalpis, Hahnia pusilia C.L.Koch, 1841 и Helophora insignis, входит в доминантный комплекс пауков березовососнового остепненного леса в Пермском Предура-лье [Есюнин, Мазура, 2000], но осенью, тогда как летом для этих лесов характерно высокое обилие Panamomops mengei Simon, 1926, формирующего моно доминантный комплекс. Заслуживает упоминания также присутствие D. bidentata и R. lividus в списке «обычных» пауков хвойных таежных лесов Финляндии. Однако обзор Копонена [Коронен, 1999] основывается на материалах, полученных методом почвенных ловушек, и, поэтому в целом интересные данные не дают представления об обилии этих видов в стратобионтном комплексе.
Для Западно-Сибирской равнины нет данных о структуре группировок стратобионтных пауков хвойных лесов. Имеется только указание Волкова [1987], который на основе ручных сборов отмечает, что в сосняках наиболее обычен вид Porrhomma pygmaeum (Blackwall, 1834). Однако некоторое представление о численности видов пауков в хвойных лесах региона можно получить из работы Та-насевича [Tanasevitch, 2005], т. к. данный обзор основывается, в первую очередь, на почвеннозоологическом материале, правда, собранном в августе-октябре, т. е. в конце лета - осенью. Согласно этой работе, наибольшим количеством особей в хвойных лесах Юганского заповедника и стационара Миссия представлены Micrargus herbigradus, Microneta viaria, P. pygmaeum и рядом других видов.
Чем обусловлено столь существенное отличие состава доминантного комплекса стратобионтных пауков хвойных лесов национального парка «Припышминские боры» на данный момент объяснить, по-видимому, невозможно. Так как, кроме различий в географическом положении, растительности, важным моментом является то, что основная часть литературных данных не привязана ко времени или охватывает другой фенологический период. Однако имеющиеся отрывочные данные о летнем аспекте группировок пауков Предуралья и Урала свидетельствуют о значительной региональной специфике набора доминантных видов.
Заключение
Полученные нами данные о структуре населения пауков лесных биоценозов национального парка «Припышминские боры» свидетельствуют о существенных различиях между группировками пауков сосновых лесопосадок и естественных лесонасаждений. Однако природа этих различий не очевидна, т.к. данные получены в различные годы, несколько разные фенологические периоды и различными сборщиками. Тем не менее, не вызывает сомнения специфичность населения пауков моло- дых (возрастом 10-15 лет) сосновых лесопосадок. Сформировавшиеся здесь комплексы имеют смешанную природу, сочетая как характеристики луговых (низкая доля в общей численности пауков из сем. Linyphiidae и значительная представленность пауков из сем. Gnaphosidae и Lycosidae), так и лесных группировок пауков (характерный для лесов доминантный вид, высокое разнообразие лесных видов).
Как общее, характерное для всего ряда лесов, проявляется тенденция роста плотности населения с увеличением возраста лесонасаждений, которая сопровождается снижением показателей разнообразия. Последнее явление связано не столько с уменьшением количества видов, которое незначительно варьирует между пробами как из лесопосадок, так и лесов, сколько с изменением выравнен-ности населения. Для проб из лесопосадок характерно кодоминирование нескольких обычных видов, тогда как для лесов более типичен монодоми-нантный комплекс.
Плотность населения пауков в изученных лесах в целом сопоставима с оценками плотности, известной для Русской равнины и Урала, тогда как состав доминантного комплекса весьма специфичен. Среди видов, доминирующих летом в группировках хвойных лесов национального парка, Среднего Урала и Пермского Предуралья, нет общих видов. Несколько неожиданно, наиболее массовый в хвойных лесах при-пышминских боров паук - D. Edentata. указывается как доминантный для ельника зеленомошного в Латвии, но в латвийских сосняках он относительно редок. Недостаток данных о структуре населения пауков России не позволяет на данный момент оценить степень своеобразия качественных характеристик изученных в национальном парке «Припышминские боры» группировок.
Авторы глубоко признательны Д. Казанцеву и Л. Шумиловской за представленные для исследования материалы. Исследования частично выполнены при поддержке Государственного задания Минобрнауки России на выполнение научноисследовательской работы № 6.1194.2014/К.
Список литературы Население стратобионтных пауков (Araneae) лесов национального парка "Припышминские боры" (летний аспект)
- Волков А.Е. К вопросу познания пауков южной подзоны тайги Тюменской области//Экология и география членистоногих Сибири. Новосибирск: Наука, 1987. С. 38-39
- Гаркуша Т.А. Пауки как компоненты почвенных ярусов лесных биоценозов//Проблемы почвенной зоологии. Вильнюс, 1975. С. 113
- Голуб В.Б., Цуриков М.Н., Прокин А.А. Коллекции насекомых: сбор, обработка и хранение материала. М.: Товарищество научных изданий КМК, 2012. 339 с
- Есюнин С.Л. Структура и разнообразие населения пауков зональных и горных тундр Урала//Зоологический журнал. 1999. Т. 78, вып. 6. С. 654671
- Есюнин С.Л., Ефимик В.Е., Мазура Н.С. Структура населения мезофауны подстилки пихто-ельников Висимского заповедника после ветровала//Последствия катастрофического ветровала для лесных экосистем. Екатеринбург: УрО РАН, 2000. С. 69-86
- Есюнин С.Л., Мазура Н.С. Структура и разнообразие мезофауны подстилки лесов с участием березы заказника «Предуралье» (Пермская область)//Вестник Пермского университета. 2000. Вып. 3. Заказник «Предуралье». С. 193204
- Есюнин С.Л., Стёпина А.С. Фауна и биотопическое распределение пауков (Aranei) подзоны южной тайги Западной Сибири//Вестник Пермского университета. Биология. 2014. Вып. 4. С. 24-54
- Есюнин С.Л., Ширпужева А.С. Многолетняя динамика населения пауков парковых колков лесостепного Зауралья//Энтомологические исследования в Северной Азии. Новосибирск, 2006. С. 225-226
- Количественные методы в почвенной зоологии. М.: Наука, 1987. 286 с
- Коробейников Ю.И., Есюнин С.Л. Состав и распределение напочвенных пауков в биоценозах южной тундры полуострова Ямал//Фауна и экология пауков. Пермь, 1995 (1994). С. 17-25
- Кудряшева И.В. Почвенные беспозвоночные (мезофауна) лесов низовьев р. Онеги//Почвенная фауна Северной Европы. М., 1987. С. 39-50
- Мазура Н.С. Структура мезофауны подстилки некоторых лесных сообществ Ильменского заповедника//Животный мир Южного Урала и северного Прикаспия. Оренбург, 1995. С. 116-117
- Национальный парк «Припышминские боры». 2016. URL: http://www.nppbor.ru/nature/flora.html (дата обращения: 5.02.2016)
- Пахоруков Н.М. Характеристика комплексов пауков лесостепных биоценозов Южного Зауралья//Труды Зоол. ин-та АН СССР. 1985. Т. 139. С. 92-98
- Пахоруков Н.М., Исакова С.В., Картышева М.В. Характеристика аранеокомплексов некоторых биоценозов лесостепного Зауралья//Фауна и экология насекомых Урала. Свердловск, 1983. С. 42-43
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 288 с
- Стриганова Б.Р., Порядина Н.M. Животное население почв бореальных лесов Западно-Сибирской равнины. М.: Товарищество научных изданий KMK, 2005. 234 с.
- Тураева А.С., Есюнин С.Л. Осенний аспект населения пауков (Aranei) национального парка «Припышминские боры»//Вестник Удмуртского университета. Серия Биология. Науки о Земле. 2015. Т. 25, вып. 3. С. 61-69
- Ухова Н.Л. Реакция почвенной мезофауны на вывал леса в пихто-ельнике высокотравно-папоротниковом//Проблемы сохранения и восстановления степных экосистем. Оренбург, 1999. С. 133-134
- Ухова Н.Л., Есюнин С.Л., Беляева Н.В. Структура населения и численность почвенной мезофауны в первичнопирогенном сообществе на месте пихто-ельника высокотравно-папоротникового//Биологическое разнообразие заповедных территорий: оценка, охрана, мониторинг. М.; Самара, 2000. С. 169-175
- Штернбергс М.Т. Структура и динамика фауны пауков (Aranei) подстилки ельника зеленомошника в Латвийской ССР//Энтомологическое обозрение. 1984. Т. 63, вып. 1. С. 188-192
- Штернбергс М.Т. Пауки (Aranei) напочвенного покрова и подстилки боров Латвии//Фауна и экология пауков СССР. Л., 1985. С. 141-146
- Ваеv P. V., Реnеv L.D. BIODIV -programs for calculation biological diversity parameters, similarity, niche overlap, and cluster analysis. Vеrsion 4.l. Sofia: PЕNSOFТ, 1993. 43 p
- Esyunin S.L. Geographical variation in spider assemblages (Arachnida: Aranei) of steppe and steppelike habitats of the Urals, Russia//Species and communities in extreme environments. Sofia-Moscow: Pensoft Publ., 2008. P. 143-158
- Koponen S. Common ground-living spiders in old taiga forests of Finland//The Journal of Arach-nology. 1999. Vol. 27. P. 201-204
- Mikhailov K.G. The spiders (Arachnida: Aranei) of Russia and adjacent countries: a non-annotated checklist//Arthropoda Selecta. 2013. Supplement 3. P. 1-262
- Tanasevitch A.V. Linyphiid spiders of the West Siberian Plain (Arachnida: Aranei)//Arthropoda Selecta. 2005. Vol. 14, № 1. P. 33-88
