Наследование и экспрессия способности к индукции соматического эмбриогенеза in vitro в онтогенезе растений перца сладкого

Автор: Тимина О.О., Цыкалюк Р.А., Орлов П.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биотехнология. Культура клеток и тканей
Статья в выпуске: 3 т.40, 2005 года.
Бесплатный доступ
Оценивали показатели экспрессии и наследования способности к соматическому эмбриогенезу in vitro у 35 различных генотипов перца сладкого на разных этапах онтогенеза. Рассматриваются механизмы регуляции индукции образования соматических зародышей на уровне генов.
Короткий адрес: https://sciup.org/142133082
IDR: 142133082 | УДК: 635.649:581.143.6:575.117.2
Inheritance and expression of capacity for induction of somatic embryogenesis in vitro in ontogenesis of sweet bell red pepper plants
The authors estimate the expression and the character of inheritance of capacity for somatic embryogenesis in vitro in 35 different genotypes of sweet bell red pepper on different ontogenesis stages. The mechanisms of genic regulation for induction of formation of somatic embryos were considered. The necessity of genetic control for formation of somatic embryoids with the purpose of prediction of regeneration capacity of certain genotypes, optimization of nutrient media and development of methods for control of determinant's expression was shown.
Текст научной статьи Наследование и экспрессия способности к индукции соматического эмбриогенеза in vitro в онтогенезе растений перца сладкого
При изучении индуцированного органогенеза и соматического эмбриогенеза особый интерес представляет, на наш взгляд, не эмпирическая разработка протоколов различных методов для отдельных генотипов, а фундаментальные исследования генетики развития изучаемого объекта и экспрессии конкретных генов. Предлагается следующая стратегия исследований соматического эмбриогенеза у растений перца сладкого: оценка генофонда с целью выделения разнокачественных доноров признака; детальная характеристика экспрессии признака в онтогенезе; выявление биотических и абиотических факторов, индуцирующих экспрессию; выявление закономерностей изменчивости и наследования признака у доноров различных типов, а также возможных плейотропных эффектов; разработка методов управления экспрессией признака.
Ранее нами были выделены генотипы перца сладкого, характеризующиеся способностью к индукции формирования соматических зародышей в культуре клеток и тканей in vitro (7). В задачу настоящей работы входила оценка степени проявления способности к индукции соматического эмбриогенеза в онтогенезе и характера ее наследования у гибридов перца сладкого, полученных при участии разнокачественных доноров.
Методика . Объектом исследования служили 35 генотипов перца сладкого Capsicum annuum L. var. annuum , включая 12 гибридов F1 (8). Определяли частоту индукции соматических эмбриоидов (%), общее количество эмбриоидов, число нормальных эмбриоидов и альбиносов-химер. В качестве эксплантов использовали незрелые зиготические зародыши различного возраста, выделенные фрагменты гипокотиля, семядолей и настоящих листьев растений, выращенных из семян в стерильных условиях на среде Мурасиге-Скуга (без гормональных добавок), разбавленной в 2 раза.
С целью оценки характера наследования способности к индукции соматического змбриогенеза в онтогенезе проводили скрещивания индуцибельных генотипов по неполной диаллельной схеме. Для этого в течение весенне-летнего периода растения выращивали в пленочных необогреваемых теплицах по общепринятой технологии. Растения материнской формы опыляли избыточным количеством отцовской пыльцы. У родительских форм и гибридов F1 извлекали зародыши на разных фазах развития — глобулярной, сердечковидной, торпедовидной, стика, полукольца и кольца по Fari (2). Выделенные зародыши, участки гипокотиля, семядолей и настоя- стик (рис. 1 и 2).
Рис. 1. Частота образования соматических эмбриоидов Capsicum annuum L. in vitro на разных фазах развития экспланта (среднее по 35 генотипам): I, II, III, IV, V, VI, VII, VIII и IX — соответственно глобула, сердце, торпеда, стик, полукольцо, кольцо, гипокотиль, семядоли и настоящий лист.
V VI VII VIII IX Фаза развития экспланта
щих листьев помещали на питательную среду, содержащую 2,4-Д (3). Опыт закладывали в 2-кратной повторности (не менее 10 флаконов в каждой). Статистическую обработку данных и оценку степени доминирования ( hp ) проводили по Жученко с со-авт. (9). Генетические параметры определяли на основе полудиаллельных скрещиваний (10).
Для вторичного индуцирования эмбриоидов каллус выделяли из семян с нарушенной семенной кожурой, содержащих как эндосперм, так и зародыш (фазы полукольца и кольца). Через 3 мес после культивирования этот каллус субкультивиро-вали на свежей среде с 2,4-Д (1). Культуру помещали на 2 нед в камеру (интенсивность освещения 3000 лк, фотопериод 16 ч), а затем на 1 мес в термостат при температуре 25 оС.
Результаты . При оценке регенерационной способности были выделены генотипы перца сладкого, у которых наблюдался как прямой, так и вторичный эмбрио-идогенез (7). Особенностью прямого образования соматических эмбриоидов как у доноров признака, так и некоторых гибридов F 1 , полученных на их основе, была строгая приуроченность к очень ранним фазам развития экспланта в онтогенезе — торпеда и
Экспланты отдельных генотипов обладали высокой способностью формировать соматические эмбриоиды даже в глобулярной и сердечковидной фазах развития (например межвидового гибрида F1 С. frutescens х С. annuum ). Это, возможно, свидетельствует о неодинаковой адаптивной способности и вследствие этого разной степени выживаемости зародыша на искусственной питательной среде в конкретной фазе развития. Такая дифференциация по выживаемости соматических зародышей может служить основой при разработке экспресс-методов оценки адаптивности к абиотическим стрессорам и связанных с ней плейотропных эффектов генов.
Следовательно, переключение гетерофазной репродукции на гомо-фазную в случае прямого эмбриоидо-генеза (использование в качестве индуктора 2,4-Д) при благоприятных условиях культивирования оказывается необратимым процессом, позволяющим предположить наличие у растений перца сладкого особой генной системы, скорее всего, генов-регуляторов. При этом эффект предполагаемого гена-переключателя развития в значительной степени зависит от физиологического состояния донора-экспланта.
Рис. 2. Структуры, формирующиеся в культуре клеток перца сладкого сорта Толстый барон на разных фазах соматического эмбриогенеза: а, б, в и г — соответственно торпедовидные, тюльпановидные, альбиносы-химеры, скопление клеток зеленой окраски (начало каллусообразования).
Фаза развития растения-донора
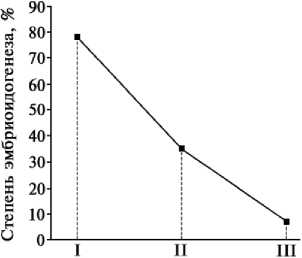
Рис. 3. Зависимость индукции соматического эмбриогенеза у эксплантов из плодов перца сладкого от стадии развития растений-доноров (среднее по пяти генотипам): I, II и III — соответственно начало технической зрелости, полная техническая зрелость и полная биологическая зрелость.
Так, у эксплантов, изъятых из плодов растений на стадии «начало технической зрелости», отмечено интенсивное формирование кластерных биполярных структур, «полная техническая зрелость» — значительное снижение индукции, «полная биологическая зрелость» — формирование в основном неэмбриогенного каллуса (рис. 3). Наиболее вероятной причиной подобной модуляции экспрессии может быть старение организма и возрастное изменение эндогенного баланса гормонов-индук-торов либо образование белков-репрес-соров.
При использовании в качестве эксплантов зародышей в фазах полукольца и кольца у подавляющего большинства генотипов, обладающих высокой способностью к переключению фаз репродукции, наблюдался только каллусоге-нез. Однако у эксплантов потомства F1 в комбинации скрещивания Колобок х Л 48 на этой же фазе развития отмечено прямое образование эмбриоидов, что подтверждает данные Harini и 67
Lakshmisita, наблюдавших аналогичный процесс у сформировавшихся зрелых зародышей перца сорта California Wonder (цит. по 2), и, возможно, свидетельствует о другом механизме каскадной активации соматического эмбриогенеза.
Вторичное образование эмбриоидов выявлено у образцов-доноров Л 49, Гогошары и Колобок. К особенностям вторичного эмбриогенеза относятся как необходимость дедифференциации и субкультивирования, так и стрессовой обработки каллуса. Известно, что при стрессе образуются белки теплового шока (БТШ), которые играют ключевую роль в регуляции клеточного цикла во время гаметогенеза у млекопитающих, а также опосредованно регулируют рост клеток, дифференцировку и экспрессию в онтогенезе некоторых ферментов (11). Возможно, что и в нашем случае БТШ, образование которых могло быть вызвано общим старением культуры, изменением рН, накоплением токсичных соединений, длительным культивированием в темноте, способствовали повышению индуцибельности и экспрес- сии генов-регуляторов.
Как показано, для успешного осуществления андрогенеза у растений перца необходимо сочетание таких факторов, как отзывчивый генотип, наличие в питательной среде гормона-индуктора, определенная стадия развития микроспор и способность последних к выживанию in vitro, подходящее физиологическое состояние донора, стрессовые условия на начальных этапах культивирования (3-5, 12, 13). Одни и те же факторы, необходимые как для андрогенеза, так и соматического эмбриогенеза, позволяют предположить хотя бы частичное совпадение генетических систем, обусловливающих эти процессы.
Разобщенность и дифференцированность образования соматических эмбриоидов у разных доноров косвенно свидетельствует, на наш взгляд, о том, что эм-бриоидогенез в условиях in vitro можно разделить на ряд этапов: индукция, форми-
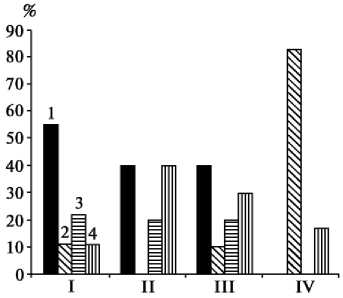
Рис. 4. Степень доминирования ( h p ) способности к переключению фазы репродукции у гибридов F 1 перца сладкого по этапам соматического эмбриогенеза: I, II, III и IV — соответственно индукция, формирование общего числа эмбриоидов, дифференциация альби-носов и нормальных эмбриоидов; 1, 2, 3 и 4 — соответственно отрицательное сверхдоминирование, отрицательное доминирование, сверхдоминирование и промежуточный характер доминирования.
рование альбиносов-химер и нормальных эмбриоидов.
У гибридов F 1 степень доминирования по способности к индукции эм-бриоидогенеза зависела от комбинации скрещивания и фазы развития экспланта (табл., рис. 4).
В целом, хотя для всего процесса и отмечено преобладание отрицательного сверхдоминирования или отрицательного доминирования, при формировании общего числа эмбриоидов выявлены отрицательное сверхдоминирование, при дифференциации альбиносов — отрицательное и положительное доминирование в одинаковой степени, при образовании нормальных эмбриоидов — резко выраженное отрицательное доминирование. Такая дифференцированность свидетельствует о том, что каждый
Оценка характера наследования способности к переключению фазы развития у эксплантов гибридов F1 Capsicum annuum L. in vitro на разных этапах соматического эмбриогенеза
|
Комбинация скрещивания |
Частота индукции соматических эмбриоидов, % |
h p |
Общее число эмбриоидов |
h p |
Число альбиносов-химер |
h p |
Число нормальных эмбриоидов |
h p |
||||||||
|
Р 1 |
Р 2 |
F 1 |
Р 1 |
Р 2 |
F 1 |
Р 1 |
Р 2 |
F 1 |
Р 1 |
Р 2 |
F 1 |
|||||
|
Ф а з а «т о р п е д а» |
||||||||||||||||
|
Л 48 х Прометей |
6,7 |
8,3 |
12,5 |
6,00 |
15 |
25 |
24 |
0,80 |
15 |
25 |
23 |
0,60 |
0 |
0 |
1 |
(+) |
|
Колобок х Прометей |
9,1 |
8,3 |
0 |
–23,00 |
29 |
25 |
0 |
–13,50 |
27 |
25 |
0 |
–26,00 |
2 |
0 |
0 |
–1,00 |
|
Л 49 х Прометей |
8,3 |
8,3 |
22,7 |
(+) |
21 |
25 |
125 |
51,0 |
21 |
25 |
113 |
45,00 |
0 |
0 |
12 |
(+) |
|
Добрыня х Прометей |
18,18 |
8,3 |
12,5 |
–0,15 |
51 |
25 |
49 |
0,85 |
49 |
25 |
49 |
1,00 |
2 |
0 |
0 |
–1,00 |
|
Колобок х Л 48 |
9,1 |
6,7 |
10,0 |
1,75 |
29 |
15 |
33 |
1,57 |
27 |
15 |
33 |
2,00 |
2 |
0 |
0 |
–1,00 |
|
Л 49 х Л 48 |
8,31 |
6,7 |
0 |
–9,00 |
21 |
15 |
0 |
–6,00 |
21 |
15 |
0 |
–6,00 |
0 |
0 |
0 |
(+) |
|
Добрыня х Л 48 |
18,18 |
6,7 |
4,2 |
–1,40 |
51 |
15 |
6 |
–1,50 |
49 |
15 |
6 |
–1,53 |
2 |
0 |
0 |
–1,00 |
|
Л 49 х Колобок |
8,31 |
9,1 |
0 |
–23,00 |
21 |
29 |
0 |
–6,25 |
21 |
27 |
0 |
–8,00 |
0 |
2 |
0 |
(–) |
|
Добрыня х Колобок |
18,18 |
9,1 |
8,3 |
–1,20 |
51 |
29 |
40 |
0 |
49 |
27 |
40 |
0,18 |
2 |
2 |
0 |
(–) |
|
Добрыня х Л 49 |
18,18 |
8,3 |
9,3 |
–0,80 |
51 |
21 |
26 |
0,67 |
49 |
21 |
24 |
–0,78 |
2 |
0 |
1,8 |
0,80 |
|
Ф а з а |
«с т и к» |
|||||||||||||||
|
Л 48 х Прометей |
30,77 |
0 |
4,3 |
–0,72 |
225 |
0 |
34 |
–0,70 |
225 |
0 |
34 |
–0,70 |
0 |
0 |
0 |
— |
|
Колобок х Прометей |
0 |
0 |
8,3 |
(+) |
0 |
0 |
11 |
(+) |
0 |
0 |
11 |
(+) |
0 |
0 |
0 |
— |
|
Л 49 х Прометей |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
|
Добрыня х Прометей |
0 |
0 |
4,2 |
(+) |
0 |
0 |
8 |
(+) |
0 |
0 |
8 |
(+) |
0 |
0 |
0 |
— |
|
Колобок х Л 48 |
0 |
30,8 |
0 |
–1,00 |
0 |
225 |
0 |
–1,00 |
0 |
225 |
0 |
–1,00 |
0 |
0 |
0 |
— |
|
Л 49 х Л 48 |
0 |
30,8 |
11,1 |
–0,28 |
0 |
225 |
6 |
–0,95 |
0 |
225 |
6 |
–0,95 |
0 |
0 |
0 |
— |
|
Добрыня х Л 48 |
0 |
30,8 |
0 |
–1,00 |
0 |
225 |
0 |
–1,00 |
0 |
225 |
0 |
–1,00 |
0 |
0 |
0 |
— |
|
Л 49 х Колобок |
0 |
0 |
8,3 |
(+) |
0 |
0 |
2 |
(+) |
0 |
0 |
0 |
— |
0 |
0 |
2 |
(+) |
|
Добрыня х Колобок |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
|
Добрыня х Л 49 |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
0 |
0 |
0 |
— |
П р и м е ч а н и е. Прочерк означает отсутствие проявления признака; h p — степень доминирования.
этап эмбриогенеза контролируется независимыми генетическими си-стемами. В комбинациях скрещивания Колобок х Прометей, Л 49 х Л 48, Добрыня х Л 48, Л 49 х Колобок, Добрыня х Колобок h p по абсолютной величине оказалась больше единицы, что свидетельствует как об эпистатичных генных взаимодействиях, так и о функционировании разных генетических систем, контролирующих этапы эмбриогенеза у использованных доноров.
Следует отметить еще один тип генных взаимодействий, характеризующих формирование нормальных эмбриоидов зеленой окраски у эксплантов в гибридных комбинациях Л 48 х Прометей и Л 49 х Прометей (см. табл.). Проведенный функциональный тест на аллелизм выявил появление в F 1 нормальных эмбриоидов, отсутствующих у соответствующих родительских форм, что свидетельствует о комплементарном типе взаимодействия генных систем. Помимо того, что это наиболее ценные идентифицированные генетические системы, обусловливающие в дальнейшем регенерацию растений, они являются разными у доноров Л 48, Л 49 и сорта Прометей.
Выявленные генные взаимодействия не позволили определить генетические параметры на основе полудиаллельной схемы скрещиваний из-за несоответствия экспериментальных данных предлагаемой модели: коэффициенты регрессии недостоверны. Поэтому для более детальной генетической идентификации доноров в дальнейшем необходимо провести генетический анализ с использованием гибридов F 2 , F 3 и беккроссов.
В результате генетического анализа эксплантов в фазе стик у гибридов Л 49 х Прометей, Добрыня х Колобок, Добрыня х Л 49 было выявлено полное затухание процесса образования соматических зародышей; Колобок х Прометей, Л 49 х Колобок — смена типа взаимодействия генов; Л 48 х Прометей, Добрыня х Прометей, Колобок х Л 48, Л 49 х Л 48, Добрыня х Л 48 — смена характера доминирования, что и обусловливало различную экспрессию способности к индукции соматического эмбриогенеза в онтогенезе (см. табл., рис. 1.).
Таким образом, у растений перца сладкого имеется по крайней мере два механизма генной регуляции образования соматических эмбриоидов: первый связан с действием предполагаемого гена-переключателя гетерофазной репродукции на го-мофазную и функционирует на ранних стадиях онтогенеза незрелого зиготического зародыша; второй — со сменой генных взаимодействий и характера доминирования и функционирует на поздних стадиях онтогенеза. И хотя пока неизвестны молекулярные механизмы каскадной активации и репрессии процесса соматического эмбриогенеза in vitro, выявленная специфика экспрессии и влияние генетического фона позволяют уже на этом этапе исследований целенаправленно проводить синтетическую селекцию и отбор на регенерационную способность у образцов перца сладкого.
Л И Т Е Р А Т У Р А
-
1. B i n z e l M.L., S a n k h l a N. Somatic embryogenesis — an approach for regeneration and transformation of pepper. In: Proc. of the 13th National Pepper Conference. Naples, Florida, USA, 1996: 38-39.
-
2. F a r i M. Impact of cell- and tissue culture techniques on the breeding of Capsicum . In: EUCARPIA IX-th Meeting on Genetics and Breeding of Capsicum and Eggplant. Budapest, 1995: 53-63.
-
3. V e n c z e l G., G e m e s n e J u h a s z A. Pepper breeding methods and strategies related with in vitro haploid research. In: EUCARPIA X-th Meeting on Genetics and Breeding of Capsicum and Eggplant. Avignon, 1998: 96-97.
-
4. G e m e s n e J u h a s z A., S a g i Z.S., S a l a m o n P. e.a. Experiens and results of in vitro haploid methods application in pepper breeding programme. In: EUCARPIA X-th Meeting on Genetics and Breeding of Capsicum and Eggplant. Avignon, 1998: 201-203.
-
5. M i t y k o J., C h a m b o n n e r D., A d a m G. e.a. In vitro haploidy of spice and bell peppers: It's control for large-scale application. In: EUCARPIA IX-th Meeting on Genetics and Breeding of Capsicum and Eggplant. Budapest, 1995: 64-67.
-
6. Б а т ы г и н а Т.Б., В а с и л ь е в а В.Е. Размножение растений. СПб, 2002: 10-13, 15.
-
7. Т и м и н а О.О., О р л о в П.А., Ц ы к а л ю к Р.А. Регенерационная способность представителей
рода Capsicum L. В сб.: Биотехнология на рубеже двух тысячелетий. Саранск, 2001: 144-145.
-
8. B a r a l J.B., B o s l a n d P.W. An updated synthesis of the Capsicum genus. Capsicum and eggplant newsletter, 2002, 21: 11-21.
-
9. Ж у ч е н к о А.А., А н д р ю щ е н к о В.К., К о р о л ь М.М. и др. Изменчивость и наследование хозяйственно ценных признаков у томатов. Кишинев, 1973.
-
10. Ф е д и н М.А., С и л и с Д.Я., С м и р я е в А.В. Статистические методы генетического анализа. М., 1980.
-
11. Х л е б о д а р о в а Т.М. Как клетки защищаются от стресса? Генетика, 2002, 38, 4: 437-452.
-
12. D u m a s d e V a u l e x R., C h a m b o n n e t D., P o c h a r d E. Culture in vitro d'anth e res des piment
(Capsicum annuum L.): am e lioration des taux d'obtention de plantes chez diff e rents genotypes par des traite ments a +35 0C. Agronomic, 1981, 1: 859-864.
-
13. A h m a d i a n T e s t i 11 a n o P.S., G o n z a l e z - M e l e n d i P., F a d o n В. e.a. Cell biology of the pollen developmental program and the induction of microspore embryogenesis in Capsicum annuum L. In: EUCARPIA X-th Meeting on Genetics and Breeding of Capsicum and Eggplant. Avignon, 1998: 183-186.
Приднестровский государственный университет
