Наследование, окраски луковицы лука репчатого

Автор: Логунов А.Н., Тимин Н.И.
Журнал: Овощи России @vegetables
Рубрика: Современные направления селекции овощных культур
Статья в выпуске: 2 (23), 2014 года.
Бесплатный доступ
В статье представлены результаты изучения наследования окраски сухих чешуй луковицы лука репчатого. Показано, что различия по окраске луковиц (желтые, коричневые, белые, красные и другие окраски) обусловлены отличиями в генотипе растений от одного до трех генов.
Лук репчатый, расщепление инбредных (i 1-i 2) и гибридных потомств (f 1, bc 1) по окраске луковиц, гены и генотипы окрасок
Короткий адрес: https://sciup.org/14025077
IDR: 14025077 | УДК: 635.25:631.524.01
Inheritance of bulb color in onion
The results of study of inheritance of bulb color in onion are presented in the article. It was shown that the differences in bulb color (yellow, brown, white, red, etc) are determined by several genes in genome.
Текст научной статьи Наследование, окраски луковицы лука репчатого
Знания о наследовании окраски чрезвычайно важны для селекционных исследований, так как она связана с другими важными признаками. К тому же в современных условиях рынка важна привлекательность и оригинальность окраски новых сортов и гибридов лука. Знание закономерностей наследования признака окраска сухих чешуй луковицы позволит селекционеру вести направленный отбор. Необходимость знания наследования этого признака имеет значение и в семеноводстве лука репчатого (Пивоваров и др., 2001; Rabinowich 2002, Khar et al. 2008). Появ- ление в сортовых посевах среди растений, имеющих красную или белую окраску луковицы, луковиц с желтой окраской или среди растений, имеющих желтую, – луковиц с красной окраской является крайне нежелательным явлением, поскольку при апробации семеноводческие посевы выбраковываются.
Изучение генетики лука ведется с 1925 года, когда были обнаружены мужски стерильные растения. С тех пор прошло более 85 лет, а число идентифицированных главных генов рода Allium L., и, в частности, лука репчатого, все еще очень ограничено, и они относятся к качественным признакам. Одним из наиболее изученных селекционно ценных признаков лука репчатого является окраска сухих че-шуй луковицы, которая варьирует от белой через различные оттенки желтой и красной до темно-красной и коричневой. В коммерческом отношении существует деление по окраске на 4 группы: белая, желтая, красная и коричневая.
Причины недостатка генетических знаний следующие: перекрестноопы-ляемая природа культуры лука, сильная инбредная депрессия, длительность получения поколений (2-3-летний цикл развития), ограниченное число работ по генетике Allium (Титова, Ершов, 1999).
Из литературы известно несколько работ, в которых показаны результаты изучения наследования окраски сухих чешуй луковицы лука репчатого (Clarke et al., 1944; El-Shafie and Davis, 1967; Khar et al., 2008). Наиболее полно представлены результаты наследования различных окрасок лука в работе El-Shafie, Davis (1967), в которой показано, что различия в окрасках луковицы (белая, бежевая, ликерная, желтая, розовая, красная) обусловлены пятью главными генами: I,C,G,L,R.
Доминантный ген I – ингибитор образования пигментов окраски. При его гомозиготном ( I I ) состоянии проявляется белая окраска луковицы, а в гетерозиготном состоянии ( Ii ) наблюдается бежевая окраска, а гомозиготный рецессивный ген ( ii ) не влияет на проявление других генов окраски.
Для образования пигментов любой окраски луковицы в генотипе растения необходим основной фактор окраски – доминантный ген C в гомозиготном CC или гетерозиготном Cc состоянии, а рецессивный ген cc обуславливает белую окраску независимо от других генов окраски.
Доминантный ген G (в гомозиготном состоянии GG и гетерозиготном - Gg состоянии) обеспечивает проявление желтой окраски луковицы. Гомозиготный рецессив данного гена ( gg ) обуславливает ликерную (темно-желтую)
окраску.
При комплементарном взаимодействии двух доминантных генов L и R проявляется красная краска луковицы.
Кроме того, имеются литературные данные, которые показывают, что разная окраска луковиц лука репчатого обусловлена взаимодействием генов по типу рецессивного или доминантного эпистаза.
В этом случае при скрещивании растений, имеющих желтую луковицу, с растениями, обладающих белой луковицей, наблюдается различный характер проявления окраски луковиц как у гибридов F1, так и гибридов F2 в зависимости от генов скрещиваемых форм. Поэтому при взаимодействии генов по типу рецессивного эпистаза (ген a>B и b ) у гибридов F1 наблюдается проявление красной окраски ( A-B- ), а в гибридах F2 отмечается расщепление – 9 красных ( A-B- ) : 3 желтых ( Abb ) : 4 белых (3 aaB -, 1 aabb ).
В другом случае, у гибридов F1 проявляется желтая окраска и наблюдается расщепление гибрида F2 по окраске луковиц, как 12 желтых ( 9 R-I- , 3 rrI- ) : 3 красных ( R-ii ) : 1 белая ( rrii ), обусловленное доминантным эпистазом, где ген R – красная окраска, при действии гена супресора I > R и r – желтая и rr – белая окраска (цит. по Лобашеву, 1967, С. 160, 167).
В наших исследованиях по изучению наследования окраски луковиц лука репчатого получены данные, которые показывают отличия от выше ука- занных особенностей наследования окраски лука, известных из литературы.
Изучение генетического контроля окраски луковицы было проведено на расщепляющихся инбредных потом-ствах I1-I2 и гибридах F1 первого поколения. Анализ ряда форм, выявленных в потомствах от самоопыления фертильных растений лука репчатого, показал их генетические различия по признаку окраска сухих чешуй луковицы.
В расщепляющихся инбредных по-томствах, полученных от форм со светло-желтой окраской сухих чешуй луковицы сортов Мячковский 300, Спасский местный и Бессоновский местный, выявлены формы со светло-желтой, темно-желтой окраской и белой окраской чешуй луковиц в различных соотношениях.
Из популяции сорта Спасский местный был проведен отбор формы, которая имела желтую окраску сухих чешуй луковицы (рис. 1). Генетический анализ инбредного потомства данной формы (№ 20) показал наличие измененных форм, имеющих фенотип, отличающийся от родительской формы. Фактическое расщепление по признаку окраска сухих чешуй луковицы составило: 88 желтые: 28 коричневые: 30 светложелтые : 10 темно-желтые луковицы, что соответствует теоретическому отношению 9:3:3:1 (χ2 =0,88 при χ205=7,81) и подтверждает гипотезу о дигенном различии признака (табл. 1).
В инбредных потомствах второго поколения I2, полученных от форм с жел-
-
1. Частоты проявления признака окраска луковицы в инбредных (I1, I2) и гибридных (F1) потомствах от формы лука №20
Номер исходной формы
Окраска исходной формы
Поколение
Фенотип
Генотип родителя
Теоретическое отношение частот
χ 2факт
χ 2теор
Желтая
Светложелт
Коричневая
Темножелт
№ 20
Желтая
I 1
88
30
28
10
BrbrGgCС
9 : 3 : 3: 1
0,88
7,81
№ 20-1-1
Желтая
I2
82
0
0
0
BrBrGGCC
0
0
0
№ 20-1-2
Желтая
I2
63
28
0
0
BrbrGGCC
3 : 1
1,61
3,84
№ 20-1-3
Желтая
I2
46
0
12
0
BrBrGgCC
3 : 1
0,57
3,84
№ 20-1-4
Желтая
I2
46
17
13
3
BrbrGgCC
9 : 3 : 3 : 1
1,36
7,81
№ 20-2
Коричневая
I2
0
0
120
49
BrbrggCC
3 : 1
1.43
3,84
№ 20-3
Светло-желтая
I2
0
32
0
13
brbrGgCC
3 : 1
0,36
3,84
№ 20-4
Темно-желтая
I2
0
0
0
40
brbrGGCC
0
0
0
№20x№20-4
Желтая х темно-желтая
F 1
24
20
18
22
BrbrGgCC x brbrggCC
1: 1 : 1 : 1
0,96
7,81
-
2. Частоты проявления признака окраска луковицы в инбредных (I1, I2) и гибридных (F1) потомствах от формы №103
Номер исходной формы
Окраска исходной формы
Генотип родителя
Поколение
Фенотип
Теоретические отношение частот
χ 2факт
χ 2теор
Желтая
Светложелтая
Коричневая
Темножелтая
Бежевая
Белая
№103-1
Желтая
BrbrGgCc
I 1
95
32
27
7
43
16
27:9:9:3:12:4
2,09
11,07
№103-1
Коричневая
BrbrggCc
I2
0
0
26
6
10
2
9:3:3:1
1,2
7,81
№103-2
Светложелтая
brbrGGCc
I2
0
28
0
0
0
8
3:1
0,1
3,84
№103-3
Белая
brbrG-cc или brbrggcc
I2
0
0
0
0
0
37
0
0
0
№103-1 x
№103-3
Коричневая х Белая
BrbrggCc x brbrggcc
F 1
0
0
35
30
28
36
1:1:1:1
2,06
7,81
-
3 .Частоты проявления признака окраска луковицы в инбредных (I1, I2) и гибридных (F1) потомствах от формы лука №313
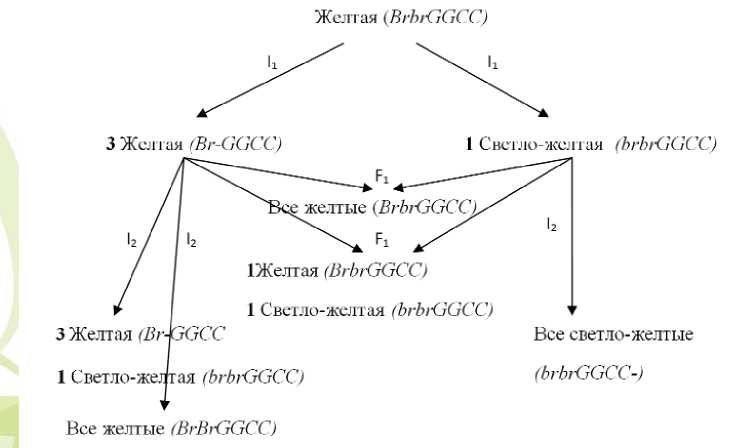
Рис. 4. Схема расщепления инбредных и кроссбредных потомств лука репчатого от формы №313 по признаку окраска сухих чешуй луковицы.
анализа и анализа расщепления в ин-бредных потомствах установлено наличие трех генов ( Br,G, C ), определяющих различия между коричневой, темножелтой, желтой, светло-желтой, бежевой и белой окрасками сухих чешуй луковицы (рис. 1, рис. 2,). При этом установлены генотипы форм растений, имеющие разную окраску луковицы: Br-G-C- – желтая; Br-G-cc , Br-ggcc – бежевая; Br-ggC- – коричневая; brbrG-C- – светло-желтая; brbrggC- – темножелтая; brbrG-cc , brbrggcc – белая окраска сухих чешуй луковицы.
В результате изучения инбредных потомств, полученных из сорта Даниловский 301 луковицы которого имеют красную окраску чешуй, были установлены фенотипические и генотипические характеристики форм по признаку
Номер исходной формы
Окраска исходной формы
Предполагаемый генотип исходной формы
Поколение
Фенотип
Теоретическое отношение частот
χ 2факт
χ 2теор
Желтая
Светложелтая
№ 313
Желтая
BrbrGGCC
I 1
44
11
3 : 1
0,73
3,84
№ 313-1
Желтая
BrBrGGCC
I2
63
0
0
0
0
№ 313-2
Светло-желтая
brbrGGCC
I2
0
24
0
0
0
№ 313-3
Желтая
BrbrGGCC
I2
59
23
3 : 1
0,41
3,84
№ 313-2x
№313-1
Светло-желтая х
Желтая
brbrGGCC x BrBrGGCC
F 1
35
0
0
0
0
№ 313-2x
№313-3
Светло-желтая х
Желтая
brbrGGCC x BrbrGGCC
F 1
26
23
1 : 1
0,18
3,84
-
4 . Частоты проявления признака окраска луковицы в инбредных (I1, I2) и гибридных (F1) потомствах от формы №3
Номер исходной формы
Окраска исходной формы
Поколение
Фенотип
Генотип
Расщеплени е
χ 2факт
χ 2теор
Красная
Розовокоричневая
№3
Красная
I 1
80
24
LlRR
3:1
0,20
3,84
№3-1
Красная
I2
53
-
LLRR
-
0
0
№3-2
Красная
I2
21
5
LlRR
3:1
0,46
3,84
№3-3
Розово-коричневая
I2
-
23
llRR
-
-
-
№3-2 x №3-3
Красная x Розовокоричневая
F 1
36
30
LlRR x llRR
1:1
0,54
3,84

Рис. 5. Расщепление по признаку окраска сухих чешуй луковицы в инбредном потомстве первого поколения I1 №3 (А – розово-коричневая, Б – красная).
той окраской № 20-1-(1,2,3,4) наблюдали три типа расщеплений в соотношении: 3 желтых : 1 светло-желтая; 3 желтых : 1 коричневых; 9 желтых : 3 коричневых: 3 светло-желтых : 1 темножелтых.
В потомствах от коричневых форм наблюдали один тип расщепления на два фенотипических класса: 3 коричневых и 1 темно-желтых. От форм, имеющих светло-желтую окраску луковицы, расщепление произошло на 3 части светло-желтых и 1 часть темно-желтую. В потомствах от форм с темно-желтой окраской сухих чешуй луковицы расщепления не наблюдали.
При проведении скрещивания между исходной желтой (№20) формой и инбредной формой с темно-желтой (№20-4) окраской сухих чешуй в гибриде F1 (№20х№20-4) отмечали расщеп-
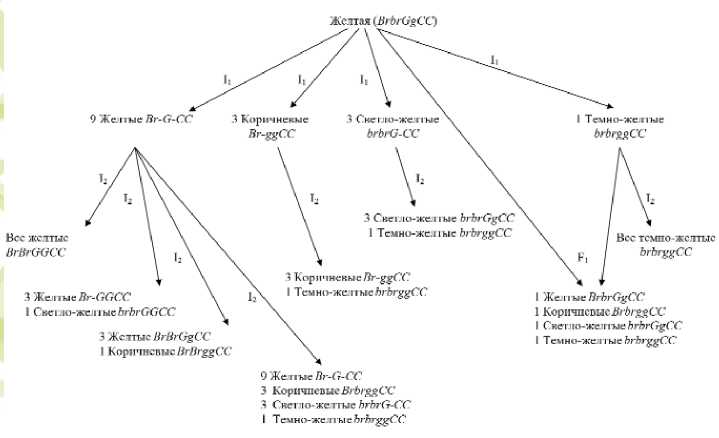
Рис. 1. Схема расщепления инбредных (I1, I2) и гибридных потомств (F1), и генотипы форм.
ление на четыре фенотипических класса: 24 желтые: 20 светло-желтые: 18 коричневые: 22 белые, что соответствует теоретическому отношению частот 1 : 1 : 1 : 1 (χ2 =0,96 при χ205=7,81). Это подтверждает, гипотезу о дигенном различии форм по данному признаку.
Так как предложенной в литературе номенклатурой генов окраски лука невозможно объяснить данные расщепления потомств, для объяснения результатов нами предлагается добавить новый ген ( Br – от англ. brown – коричневый), доминантная аллель которого в сочетании с рецессивным геном gg детерминирует коричневатый оттенок, а рецессивная аллель ( br ) – его отсутствие. Сочетание двух доминантных аллелей генов Br и G определяет проявление желтой окраски.
Таким образом, расщепление, на- блюдаемое в инбредном потомстве формы №20, объясняется дигенным различием. Исходя из этого предположения, инбредное потомство формы №20 имеет следующие генотипы: Br-G-CC – желтая, Br-ggCC – коричневая, brbrG-CC – светло-желтая, brbrggCC – темно-желтая окраска (рис.1).
Исходная форма № 103 из сорта Спасский имела желтую окраску сухих чешуй, и в результате самоопыления в потомстве проявились растения, отличающиеся от родительской формы, при этом отмечено расщепление инбред-ного потомства на 6 фенотипических классов по окраске луковицы.
Расщепление по окраске сухих че-шуй луковицы составило: 95 желтые : 43 бежевые : 27 коричневые : 32 светложелтые : 7 темно-желтые : 16 белые (табл.2, рис. 2, 3). Проведенный анализ показал, что наследование окраски обусловлено тремя генами (χ2факт=2,09 при допустимом значении χ205=11,07), а доля генотипов представлена таким образом: 27 желтые : 12 бежевые : 9 коричневые : 9 светло-желтые : 3 темножелтые : 4 белые луковицы (табл. 2, рис.3).
В инбредном потомстве второго поколения I2, исходная форма № 103-1 которого имела коричневую окраску луковицы, появились формы, отличающиеся от исходного растения: 26 – коричневые, 6 – темно-желтые, 10 – бежевые, 2 – белые луковицы, что соответствует отношению частот как 9:3:3:1 (χ2факт=1,2 при χ205=7,81).
В результате самоопыления светло-желтой формы I2 №103-2 наблюдали расщепление: 28 светло-желтых, 8 белых луковиц. Проведенный анализ χ2 показал, что наследование обус-
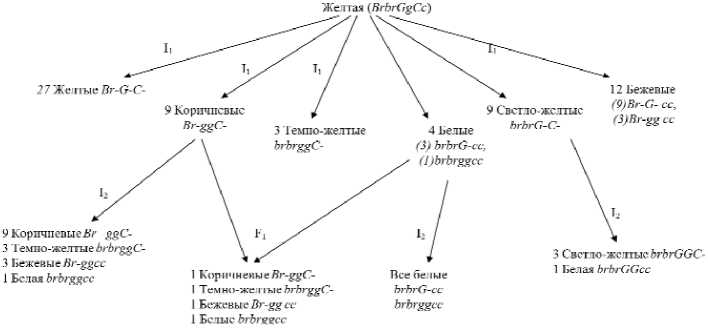
Рис. 2. Схема расщепления инбредных (I1I2) и гибридных потомств (F1), от формы №103.
ловлено моногенным различием, отношением частот как 3:1 (χ2факт=0,11 при χ205=3,84). В инбредном потомстве I2 №103-3, полученном от белой формы, расщепления не наблюдали. В анализируемом скрещивании форм коричневой и белой окрасок сухих че-шуй луковицы в гибриде первого поколения F1 проявились четыре типа разных окрасок: 35 растений имели коричневую окраску: 30 темно-желтую: 28 бежевую: 36 белую, что соответствует отношению частот как 1:1:1:1 (χ2 =2,06 при χ205=7,81) и подтверждает дигенное наследование различий данного признака.
Для установления генотипов форм инбредного потомства №103, мы предполагаем, что если в генотипе присут- лученной от сорта Мячковский 300, имеющей у исходной формы желтую окраску сухих чешуй, наблюдали расщепление на 2 фенотипических класса: 44 желтые : 11 светло-желтые, что соответствует теоретически ожидаемому отношению частот как 3:1. Анализ критерия Пирсона подтвердил гипотезу о моно-генном различии по данному признаку (χ2 =0,73 при χ205=3,84) (табл. 3, рис.4).
Во втором поколении инбридинга I2 в потомствах от форм №313-1 и №313-2 расщепления не наблюдали, а в потомстве №313-3, исходная луковица которого имела желтую окраску, произошло расщепление на два типа окрасок в соотношении 3 желтая и 1 светло-желтая (χ2=0,41 при χ205=3,84).
При скрещивании форм желтой со светло-желтой окраской сухих чешуй, в гибриде первого поколения F1 313-2x №313-1 наблюдали единообразие с доминированием желтой окраски сухих чешуй луковицы. Во втором случаи при скрещивании 313-2x 313-3 в гибриде F1 наблюдали расщепление на два типа окрасок в соотношении 1 желтые и 1 светло-желтые (χ2=0,18 при χ205=3,84).
Результаты анализа расщепления инбредных потомств I1-I2, полученных от самоопыления гетерозиготных форм (подобие гибридов F2), и расщепляющихся гибридов F1, полученных от скрещивания гетерозиготной формы с гомозиготной (подобие гибрида BC), дали возможность установить генетические особенности разных окрасок луковицы лука репчатого. Окраски луковиц обусловлены следующими генами: желтая определяется наличием доминантного гена G в сочетании с доминантными генами Br и С , светло-желтая – доминантный ген G в сочетании с рецессивным геном br при наличии доминантного гена C , темно-желтая окраска – двумя рецессивными генами br и g при наличии доминантного гена C , коричневая – доминантным геном Br и наличием рецессивного гена g и белая окраска проявляется при наличии рецессивного гена br и гена с – ингибитора генов G и g .
На основании гибридологического ствует рецессивная аллель гена ингибитора c (c>G, c>g), которая определяет проявление белой окраски, то при наличии доминантной аллели гена Br в генотипах Br-G-cc или Br-ggcc проявляется бежевая окраска сухих чешуй луковицы. А присутствие рецессивной аллели гена br в генотипах brbrG-cc или brbrggcc окраска сухих чешуй становится белой (табл.2, рис. 2).
Исходя из вышеизложенного, можно предположить, что инбредное потомство №103 имеет следующие генотипы: Br-G-C- , – определяет желтую окраску, Br-ggC- коричневую, brbrggCc- темно-желтую, а генотип brbrG-C- – светло-желтую окраску. Бежевая окраска сухих чешуй луковицы представлена следующим генотипами Br-G-cc, Br-ggcc , а белая окраска определяется brbrG-cc и brbrggcc (рис.2).
Подобное взаимодействие генов окраски сухих чешуй луковицы также мы отмечали у инбредной семьи № 313, по-

Рис.3. Расщепление по признаку окраска сухих чешуй луковицы в инбредном потомстве первого поколения I 1 №103 (верхний ряд слева направо: коричневая; темно-желтая; желтая; нижний ряд: светло-желтая; бежевая; белая).
окраска сухих чешуй лука репчатого.
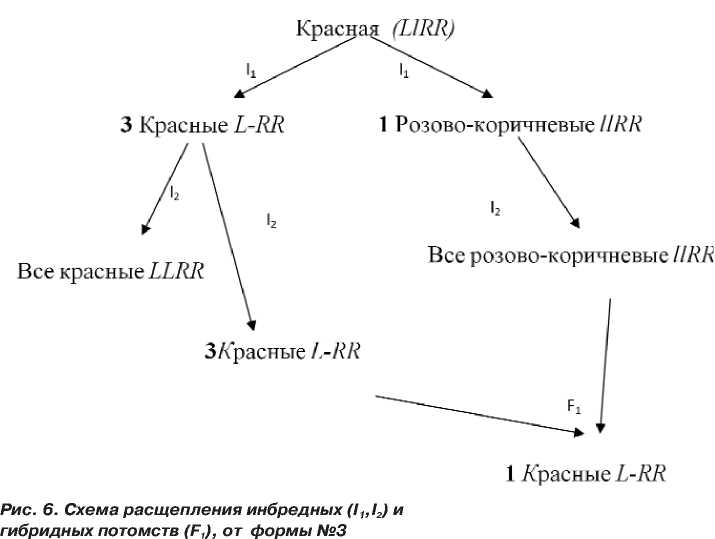
В инбредном потомстве №3 (рис. 5, 6), полученном от самоопыления растения, которое имело красную окраску сухих чешуй, наблюдали расщепление на 2 фенотипических класса, где 80 луковиц имели красную и 24 розово-коричневую окраску сухих чешуй, что соответствует теоретическому расщеплению как 3:1 – χ2факт=0,20 при χ205=3,84 (табл. 4).
В инбредных потомствах второго поколения I2 №3-1 и №3-3 расщепления не наблюдали, а потомство № 3-2 расщепилось на 2 фенотипических класса: 21 красные и 5 розово-коричневые, что соответствует ожидаемому отношению частот как 3:1 (χ2факт=0,46 при χ205=3,84.). В анализирующем скрещивании F1 №3-2х №3-3 наблюдалось расщепление 36 красных и 30 розово- коричневых, что соответствует теоретическому отношению частот как 1:1 (χ2факт=0,54 при χ205=3,84.) и подтверждает тот факт, что данный признак обусловлен моногенным различием.
Из литературных источников известно (El-Shafie and Davis, 1967; Khar 2008), что красная окраска луковицы обусловлена наличием двух комплементарно действующих доминантных генов ( L-R- ), которые подавляют проявление всех остальных генов окраски. Исходя из этого, мы предполагаем, что генотипы инбредного потомства №3 имеют следующий вид L-RR – красные и llRR – коричнево-розовые (рис.5,6).
Таким образом, исследованиями ин-бредных форм (I1-I2), полученных от самоопыления растений сортов лука репчатого, установлены генотипы инбред-ных форм, определяющие окраску сухих чешуй луковицы, которые могут быть использованы при получении и идентификации инбредных линий в се- лекции.
Список литературы Наследование, окраски луковицы лука репчатого
- Лобашев М. Е. Генетика: Учебник. -2-е изд. -Л.: ЛГУ, 1967.
- Пивоваров В.Ф., Ершов И.И., Агафонов А.Ф. Луковые культуры. -М.,2001. -497 С.
- Титова И.В., Ершов И.И. Генетика лука репчатого.//Генетические коллекции овощных растений. -СПб., 1999. -Ч.2 -С. 52-69.
- Clarke A.E., Jones H.A., Little T.M. Inheritance of bulb color in the onion//Genetics. -1944. -V.29. -Р 569-575.
- El-Shafie M.W., Davis G.N. Inheritance of bulb color in the Allium cepa L.//Hil-gardia. -1967. -V. 38. -Р.607.
- Khar A., Jekse J., Havey M.J. Segregation for onion bulb colors reveal that red is controlled by at least three loci.//J. Amer. Soc. Hort. Sct. -2008. V.133(1). -Р.42-47.
- Rabinowitch H.D., Currah L. Allium. Crop Science: Recent advances. -2002. -544 p.
